
Abstract
Foot-and-mouth disease virus (FMDV) causes an acute, highly contagious disease of livestock. Though FMDV is very sensitive to interferon-α (IFN-α), IFN-β, and IFN-γ, the virus has evolved mechanisms to evade such innate responses. For instance, during acute infection, FMDV suppresses IFN-α production by skin and myeloid dendritic cells (DCs). We have previously reported that FMDV infection induces a transient lymphopenia and interruption of T-lymphocyte responses to mitogenic stimuli. To further understand the immunopathogenesis of FMD, we have now analyzed the serum IFN-α response in relation to lymphopenia, and the number and function of plasmacytoid DCs (pDCs) following infection of pigs with multiple serotypes of FMDV. Serum IFN-α peaked 2–3 d post-infection (PI), regardless of FMDV serotype. Lymphopenia coincided with peak viremia and the serum IFN-α response. Circulating pDC numbers and in-vitro pDC IFN-α secretion transiently declined by 48 h following infection. Infection of lymphocytes or pDCs was never detected regardless of the FMDV serotype inoculated or the age of the animal infected. These data indicate that, like other DC subsets, there is suppression of interferon production by pDCs, which abrogates this important innate response. Rapid induction of serum IFN-α, albeit short-lived, may contribute to the rapid resolution of FMDV viremia prior to induction of specific immunity.
Introduction
FMDV is very sensitive to interferons (IFNs), including IFN-α, IFN-β, and IFN-γ (9,26), and numerous studies suggest that these IFNs may have a role in the resolution of FMDV infection. Type I IFNs were first identified as humoral antiviral agents (21). They constitute a first line of defense against viral infections, inducing the expression of IFN-stimulated genes, which inhibit viral replication in susceptible cells (14,42). Chinsangaram et al. have shown that IFN-α-induced RNA-dependent protein kinase (PKR) and 2′-5′-A synthetase/RNase L (OAS) prevent the replication of FMDV in otherwise permissive cells (7). In addition, inoculation of animals with adeno-vectored porcine IFN-α gene leads to protection against FMDV in swine (8), and significantly reduces the severity of the infection in cattle (40). Furthermore, porcine skin DCs are resistant to FMDV because they constitutively express IFN-α and secrete both IFN-α and IFN-β upon exposure to the virus (3).
Other viruses have evolved mechanisms for blocking innate antiviral responses. For example, poxviruses have evolved a number of mechanisms for blocking IFN production and the cellular response to IFN. During infection, toll-like receptor (TLR) stimulators like double-stranded RNA (dsRNA) are sequestered, blocking activation of IFN response molecules like dsRNA-dependent protein kinase. These viruses also encode mimicking cytokines, chemokines, and their receptors, blocking the function of these molecules. There is also interference with secondary signaling molecules, such as blocking degradation of IκB (18,35,36). In addition, bovine herpes virus (27) and pseudorabies virus (1) have been shown to down-modulate expression of MHC class I, making virus-infected cells refractory to NK or CTL killing.
In vitro, infection with FMDV is characterized by the early production of the viral leader protease, which cleaves elongation factor 4G, shutting off cap-dependent translation of cellular mRNA (12). A consequence of this is a blockage of host-cell protein synthesis, which results in inhibition of IFN-α and IFN-β production in infected cells (7,9), permitting viral replication, assembly, and new virus production (15). Furthermore, this results in the lack of the ability of infected cells to translate new MHC molecules containing viral peptides via the endogenous pathway, compromising viral peptide presentation to MHC class I-restricted T cells (34).
During the acute phase of infection in swine, another potential mechanism of immune evasion may be playing a significant role in pathogenesis. We have previously demonstrated that FMDV infection induces a severe lymphopenia and reduced T-cell proliferative response to mitogen. The lymphopenia induced by FMDV is transient, corresponds to the peak of viremia, and there is no infection of lymphocytes in vivo. The recovery from the lymphopenia is rapid (2). Furthermore, T-cell proliferative responses to the mitogen, Con A, are lost. Contrarily, analysis of an in-vitro propagated C strain of FMDV showed infection of lymphocytes in vivo (13). Also, during acute FMDV infection of swine, production of IFN-α from monocyte-derived DCs (MoDCs) and skin-derived DCs (skin DCs) is inhibited. This effect is concurrent with rising viral titer in the blood; however, these cells are not productively infected. Interestingly, there are no changes in the capability of these DCs to take-up particles and process antigens, indicating that the antigen-presenting cell function is normal (28).
In the present studies, we have analyzed FMDV induction of immunosuppression during acute infection of swine to understand the effect of FMDV on porcine plasmocytoid DC numbers and function. Furthermore, we addressed contradictory reports in the literature by comparing lymphopenia and infectivity of lymphocytes by various serotypes of FMDV. Regardless of viral strain, infected pigs showed typical signs of FMDV, including fever, which peaked on days 2–3 post-infection (PI), and was concurrent with viremia and lymphopenia. Lymphocytes isolated during peak viremia were not infected by virus isolation or real-time PCR, and serum IFN-α was detected in all infected pigs 2–3 d PI, again regardless of FMDV serotype. In all animals, circulating pDCs declined sharply in number at 48 h following infection, but returned to pre-infection levels by days 5–6 PI, and pDC function was depressed. These data indicate interruption of IFN-α production by pDCs during the acute phase of infection. However, the early systemic IFN-α response may already initiate an antiviral response in tissues, leading to the rapid resolution of FMDV viremia prior to the induction of specific immunity.
Materials and Methods
Viruses
FMDV A24 Cruzeiro (A24) and O1 Campos (O1C) are natural isolates from Brazil that have been passed in pigs and vesicular fluid diluted in supplemented MEM for inoculation (3). The Plum Island Animal Disease Center (PIADC) Institutional Animal Care and Use Committee approved all procedures performed on these animals. FMDV C-s8C1 is a plaque-purified derivative of the natural isolate of serotype C1 Santa Pau Spain 70. This has been repeatedly passed on BHK-21 and thus is adapted for heparan sulfate binding (20). Pigs at PIADC were infected with the plaque-purified C-s8C1 (FMDV C-s8C1 BHK), and virus collected from vesicular fluid was diluted in supplemented MEM (C-s8C1 PIADC PP1). C-s8C1 PIADC PP1 was again passed in pigs, and virus collected from vesicular fluid was diluted in supplemented MEM as C-s8C1 PIADC PP2. FMDV C3-Resende is a natural isolate (33) that was pig passed prior to use in this experiment. Viral titers were determined on BHK-21 cells by a standard protocol (2).
Animals, virus challenge, and blood sampling
Groups of Yorkshire pigs 2–3 mo old and 6–8 mo old were purchased from Animal Biotech, Inc. (Danboro, PA), and allowed 1 wk to acclimatize before the start of the experiments. Each pig was inoculated with virus intradermally in the heel bulb of both hind feet with 100 μL of 1 × 105 plaque-forming units of either FMDV serotype A24 Cruzeiro, O1 Campos, C3 Resende, plaque-purified C-s8C1, or pig-passed C-s8C1 (C-s8C1 was isolated from vesicles of infected pigs inoculated with plaque-purified virus [C-s8C1 PP1], or C-s8C1 isolated from vesicles of infected pigs inoculated with C-s8C1 PP2 [C-s8C1 PP2]). The animals were examined for clinical signs of infection.
Heparinized and nonheparinized blood collection tubes (Becton Dickinson, Franklin Lakes, NJ) were used for daily blood collection from the anterior vena cava. Serum was obtained from the blood drawn into nonheparinized tubes. Meanwhile, heparinized blood was diluted in an equal volume of PBS and layered over a separation gradient (Lymphoprep™; Life Technologies, Grand Island, NY) for PBMC isolation following the manufacturer's protocol. Whole blood was analyzed on an AcT hematology analyzer (Beckman Coulter, Miami, FL) to determine the percentage of lymphocytes.
Interferon-α ELISA
The IFN-α concentration in serum was determined by an antigen-capture ELISA as previously described (3). Briefly, anti-porcine IFN-α monoclonal antibodies K9 and biotinylated F17 (PBL Biomedical Laboratories, Piscataway, NJ), respectively, were used for capture and detection. The IFN-α concentration in picograms per milliliter was determined by extrapolation from a standard curve.
Virus isolation and detection
Viremia was quantified by a standard assay for isolating virus (31). Briefly, serial 10-fold dilutions of serum samples were added to confluent monolayers of BHK-21 cells in quadruplicate wells in the 96-well format and incubated at 37°C for 24–48 h. The plates were observed for CPE and stained with crystal violet when CPE was detected in the positive control wells. TCID50 was calculated from the dilutions. Samples negative for virus isolation were confirmed using real-time PCR detection of viral RNA as previously described (6).
Flow cytometry
CD4a FITC (74-12-4; BD Biosciences, San Jose, CA) and CD172 PE (Southern Biotech, Birmingham, AL) were used in flow cytometry to identify porcine plasmacytoid DCs. Briefly, 1 × 106 PBMCs were washed in chilled FACS buffer prior to being stained for 30 min on ice with optimal dilutions of the above antibodies or their respective isotype controls. Data were acquired by flow cytometry (FACSCalibur; BD Biosciences) and analyzed with Cell Quest Pro (BD Biosciences) for the percentage of CD4+CD172low pDCs, which was obtained by subtracting the background values.
Phenotype and ELISpot detection of interferon-α-producing cells in PBMCs
Owing to the limited number of pDCs in blood, our experiment was designed to quantify the responses of these cells without the need for separating them from the PBMCs. To attain this goal, we used the unique capability of porcine pDCs to respond to type A CpG (CpG 2216). Guzylack-Piriou et al. have previously shown that CD4CD172 double-positive pDCs in the PBMCs of swine are uniquely responsible for IFN-α responses to CpG 2216 (16). However, we started by confirming that the cells secreting IFNα in response to CpG 2216 expressed both CD4 and CD172. Negative selection with Dynal beads (Dynal Corp., Invitrogen, Carlsbad, CA) was used according to the manufacturer's protocol to deplete PBMCs of various cell types, and the remaining cells were tested for IFN-α secretion in response to CpG 2216. Briefly, PBMCs were incubated with mouse antibodies to specific porcine leukocyte markers, followed by the addition of anti-mouse IgG-coated beads. The bead-positive cells were then separated with a magnetic field. T and B lymphocytes were eliminated with antibodies to CD3 and CD21, while using antibodies to CD14 and CD4 eliminated monocytes and pDCs plus CD4+ T cells, respectively. With antibodies to CD172, the PBMCs were depleted of monocytes and DCs. For positive selection to enrich pDCs, the PBMCs were first cultured for 1.5 h to eliminate monocytes by adherence to plastic, then depletion of T and B cells from the non-adherent fraction was performed, followed by incubation with a cocktail of antibodies to porcine CD4 and CD172. The CD4- and CD172-positive cells were then freed of the beads by enzymatic treatment according to the manufacturer's protocol. The efficiency of depletion of cells relative to whole PBMCs was assessed by flow cytometry. The required cell fractions were washed with and suspended in lymphocyte culture medium (LCM; RPMI-1640 with 10% FBS and antibiotics; Life Technologies). For analysis of IFN-α production, whole PBMCs or sorted cells were cultured in 24-well plates at 1.0 × 106 total cells in 1.0 mL LCM, and immediately stimulated with 10 μg/mL CpG 2216. After 24 h of incubation at 37°C, 5% CO2, and 85% relative humidity, the supernatants were collected and assayed for IFN-α by ELISA as described.
Having established that the cells responding to CpG 2216 were pDCs, these cells were quantified by ELISpot determination of the CpG-induced IFN-α spots. Thus by ELISpot we monitored the effect of FMDV infection in pigs on the number of circulating pDCs responding to CpG 2216. ELISpot was performed according to a previously published protocol (29), with some modifications. Briefly, 96-well MultiScreen™ filtration plates (Millipore Corporation, Bedford, MA) were coated overnight at 4°C with 100 μL of 1.0 μg/mL K9 (PBL) in carbonate/bicarbonate buffer. The plates were washed four times with sterile PBS (pH 7.4), blocked with LCM for 2 h at 37°C, and washed again with sterile PBS. PBMCs stimulated with 10 μg/mL of CpG 2216 in sterile Falcon tubes were dispensed in quadruplicate at 105 cells/well in 100 μL, and the plates were incubated overnight at 37°C. The remaining content of the tubes (0.6 mL) was also incubated overnight and the supernatants harvested for IFN-α ELISA as described above. The plates were washed three times with PBS, and once with distilled water to lyse any remaining cells, and rinsed with PBS. The plates were incubated at 37°C for 1 h with 100 μL/well of F17 (PBL) biotinylated at 1 μg/mL in PBS containing 0.5% Tween 20 (PBST). After four washes with PBS, 0.8 μg/mL streptavidin-HRP in PBST (100 μL/well) was added and the plates were incubated at room temperature for 30 min. The plates were washed four times with PBS, and the spots were developed with 100 μL/well of 3,3′-diaminobenzidine (DAB; Sigma, St Louis, MO) containing 0.3% hydrogen peroxide. After 3 min of incubation with DAB, the plates were washed with distilled water, dried overnight, and the spots were counted on an ImmunoSpot reader (CTL Analyzers LLC, Cleveland, OH). The number of spots corresponds to the number of IFN-α-secreting cells (pDCs) responding to CpG 2216.
Statistical analysis
Differences in serum IFN-α response, pDC numbers, and pDC response to CpG 2216 between various days were analyzed by one-way ANOVA and the Newman-Keuls multiple comparison test using GraphPad Prism Software version 5 (GraphPad Software Inc., San Diego, CA).
Results
Infection of swine with FMDV
All challenged animals showed clinical signs of FMD, with development of fever and vesicles on all four feet, snout, and tongue (data not shown). Peak viremia was detected at 2–3 d PI, and it had cleared by 4 d following inoculation (Fig. 1A–G). These results were consistent with our previous report (2), but in conflict with another report showing viremia detected from day 1 to day 10 PI in swine following inoculation of plaque-purified FMDV, strain C-s8C1 (13). Here, using this same strain, we observed viremia on day 1 PI, which rapidly cleared by day 4. All animals had clinically recovered by day 14 PI.
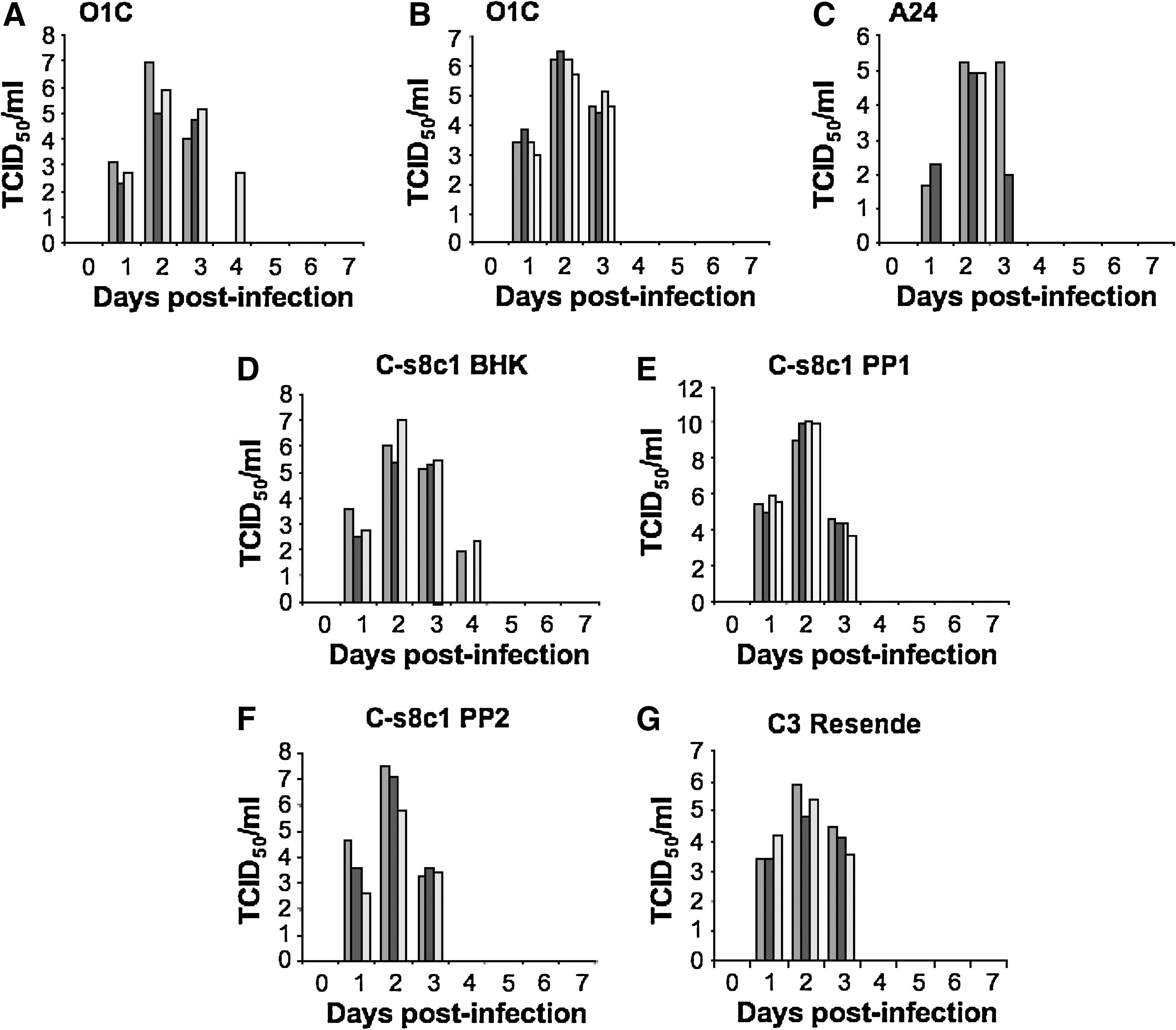
Viremia in FMDV-infected pigs. Pigs were inoculated with virus intradermally in the heel bulb and serum was collected daily for virus isolation. (
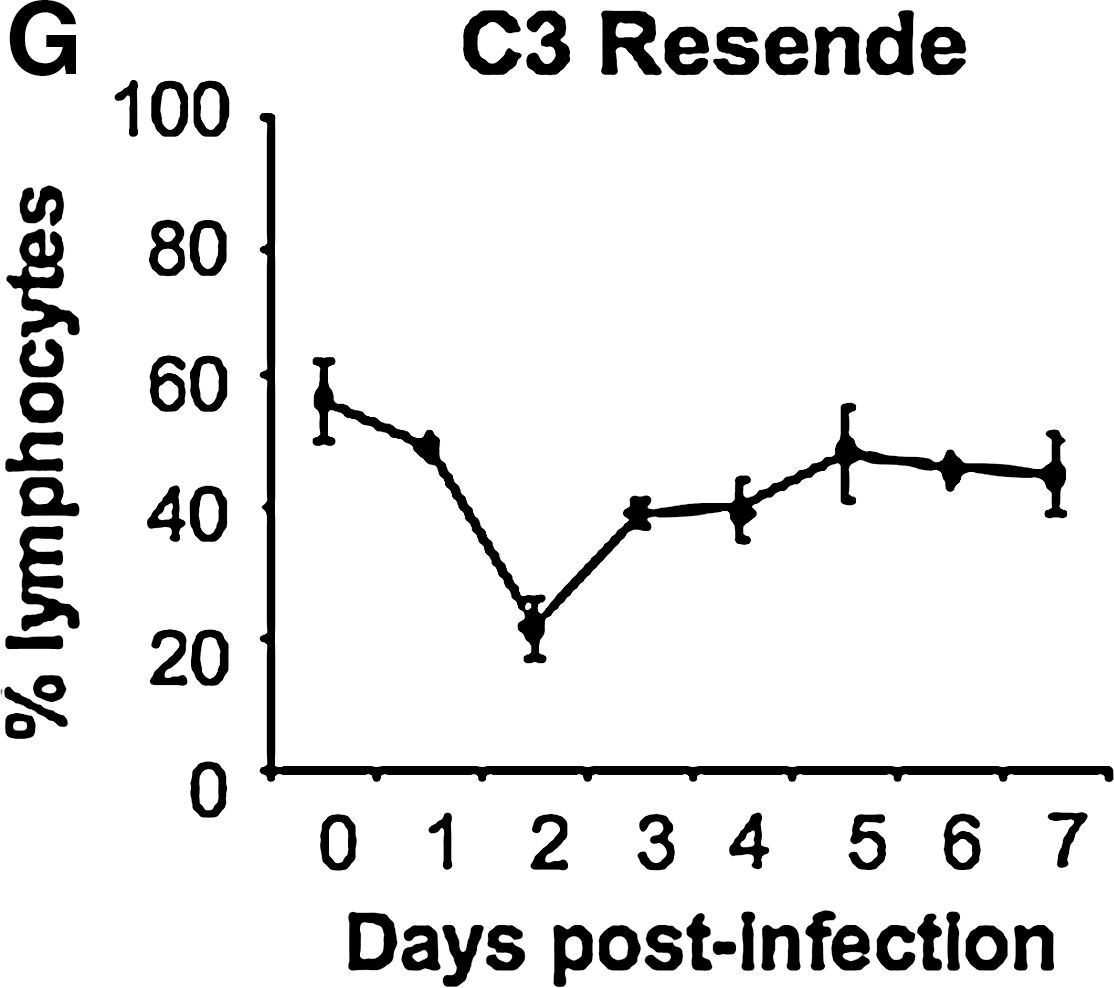
We have previously reported that during infection of young pigs with FMDV strains O1 Taiwan 97 and O1 Campos, a lymphopenia occurs with onset concurrent with the development of viremia (2). We now report identical results with the infection of older (Fig. 2A) and younger (Fig. 2B) animals with O1 Campos. In addition, a lymphopenia was observed when pigs were infected with A24 Cruzeiro and C3 Resende (Fig. 2C and H), indicating that the lymphopenia induced by FMDV is neither serotype-specific nor age-dependent. The results for the plaque-purified virus, C-s8C1, were less clear. First, variability between animals was pronounced, and in addition, the percentage of lymphocytes in some animals declined over the 2-wk study period (Fig. 2D). Testing FMDV strain C-s8C1 harvested from vesicles of these animals (animal-derived virus preparations) in subsequent animal inoculations showed a more familiar pattern, with lymphopenia observed at day 2, and resolving by day 4 (Fig. 2E).
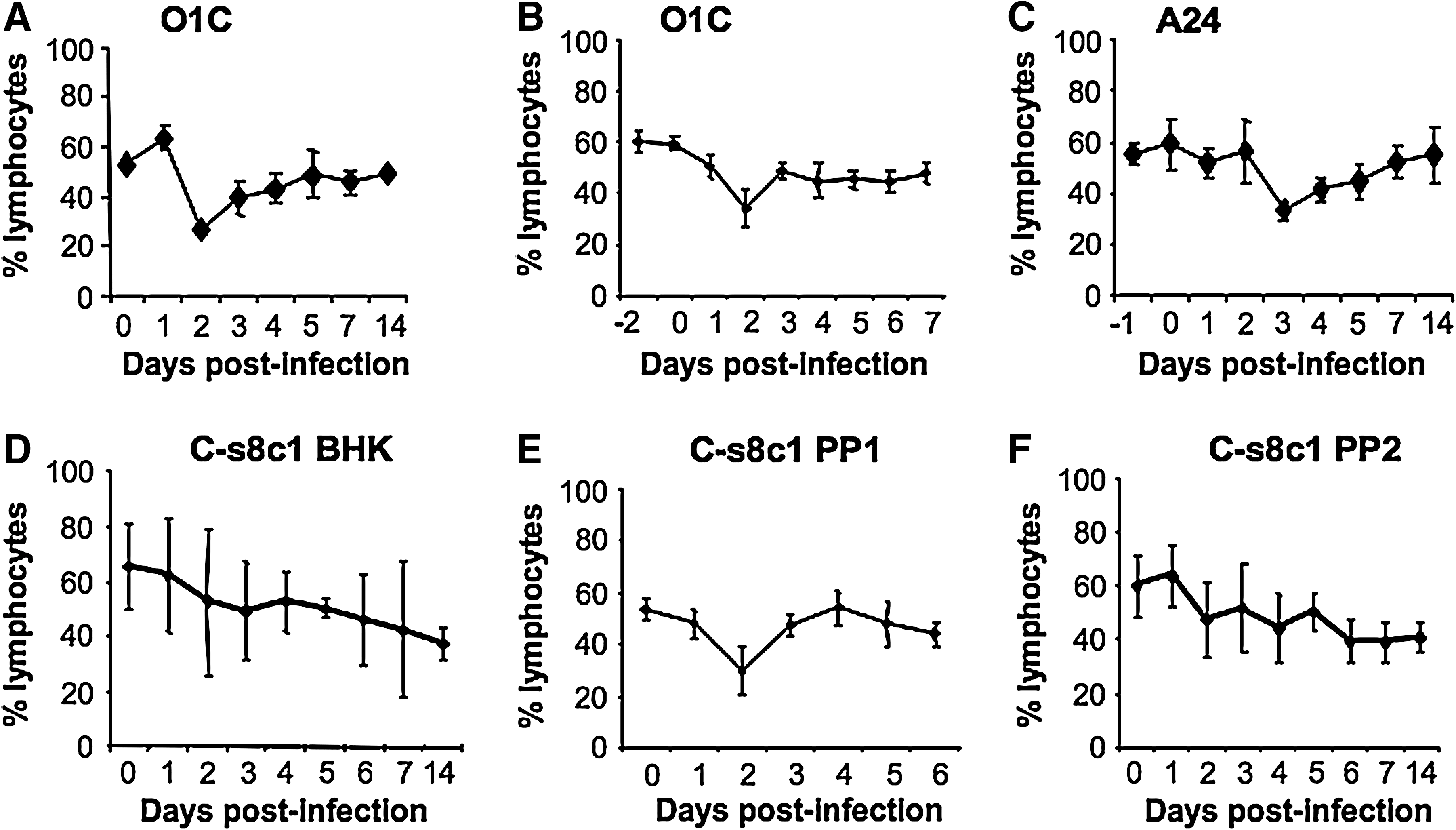
Lymphopenia in FMDV-infected pigs. Whole blood collected from FMDV-infected pigs was analyzed for percentage of lymphocytes. (
Finally, we confirmed our previous report that porcine PBMCs are not infected by O1 Campos, and further show that PBMCs of swine are neither infected by A24 Cruzeiro nor by C-s8C1 and C3 Resende in both in-vivo and in-vitro exposure. In all cases, virus isolation was negative (data not shown). We tested lysates of PBMCs by real-time PCR and virus isolation on BHK-21 cells and all were negative (data not shown). We also co-cultured the PBMCs with BHK-21 cells and also detected no virus regardless of the strain of FMDV used to infect the animals. All of this occurred while the plasma contained readily detectable virus (Fig. 1). The data presented here for the strain C-s8C1 contradicts a previous report by Diaz-San Segundo et al. (13), in which they used the in-vitro propagated, plaque-purified C-s8C1 virus, and showed infection of lymphocytes over an extended period.
Serum IFN-α in pigs infected with FMDV
In vitro, IFN-α has been shown to have strong antiviral effects on FMDV (7,9). In addition, serum IFN-α has been linked to lymphopenia in swine (37) and other animal species (24). Given the rapid clearance of viremia and the transient lymphopenia in porcine FMDV infection, we sought to determine if there was induction of IFN-α secretion early during infection. All pigs had detectable levels of serum IFN-α on day 1 PI, and levels peaked on days 2–3 (Fig. 3). Although peak levels of IFN-α were highly variable, ranging from 200–1500 pg/mL, these reached statistical significance on day 3 compared to day 0 (p < 0.05) in all the study groups. The early response pattern was similar in all sero-types of FMDV tested. However, a biphasic response of serum IFN-α was detected between days 8 and 12 following infection in a few animals, though none of these were statistically significant compared to pre-infection levels of IFN-α in that animal.
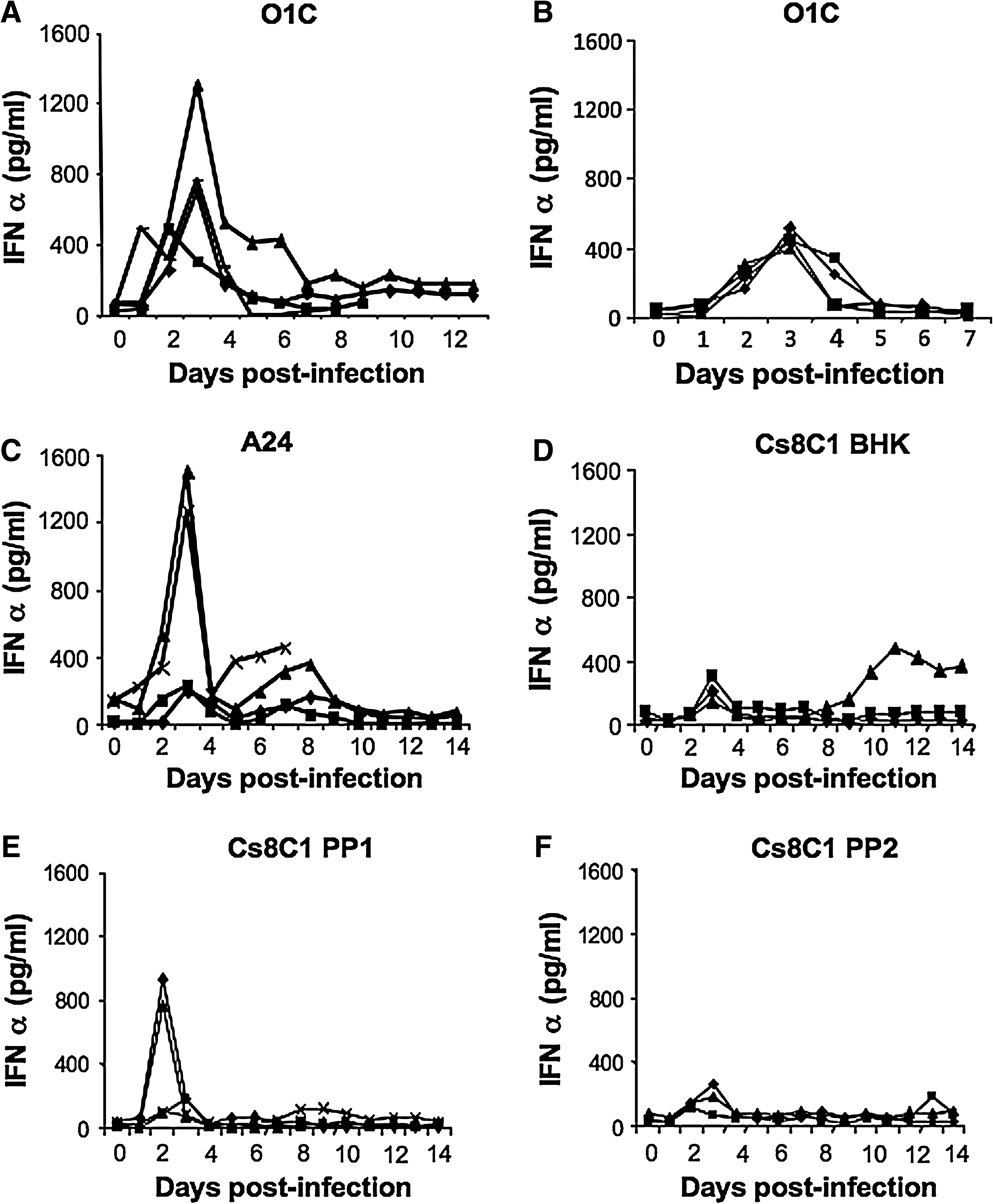
Porcine serum IFN-α response to FMDV infection. Serum collected from FMDV-infected pigs was assayed for IFN-α by a sandwich ELISA. (

Phenotype of cells in porcine PBMCs responding to CpG 2216
Plasmacytoid DCs usually constitute less than 1% of the PBMCs, and secrete IFN-α in response to viral infection. Porcine pDCs express high levels of CD4 and low levels of CD172, and produce large amounts of IFN-α in response to type A CpG (16). By flow cytometry, we identified this population in porcine PBMCs (Fig. 4A). Cells prepared by negative selection confirmed that pDCs are indeed the cells in porcine PBMCs responding to CpG 2216 by secreting IFN-α (Fig. 4B). This was confirmed by analyzing cells positively selected with anti-CD4 and anti-CD172 (Fig. 4B).
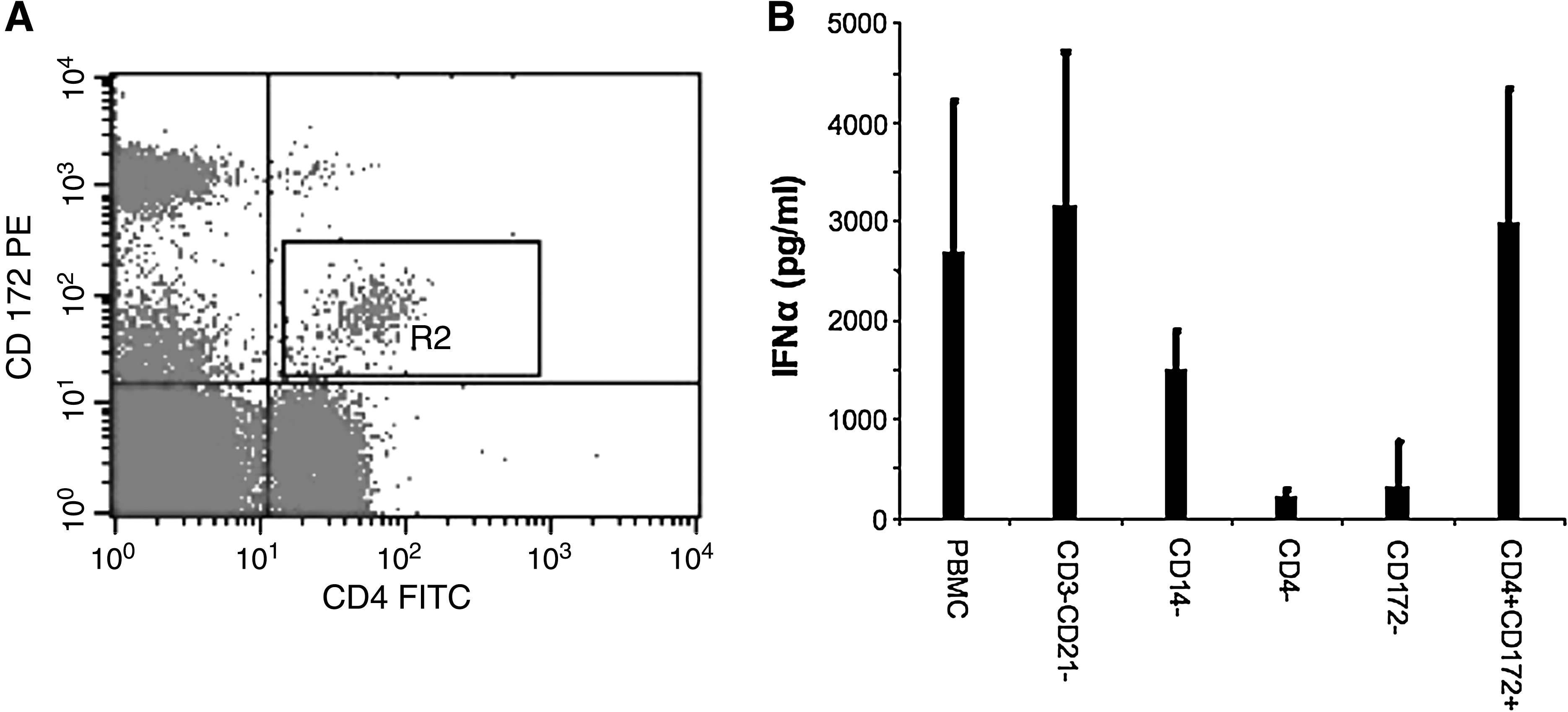
Phenotype of porcine pDCs and PBMCs responding to CpG 2216. (
Effect of FMDV infection on number and function of circulating pDCs during acute infection
Since FMDV infection of pigs modulates the immune response via lymphopenia and results in diminished IFN-α secretion by skin DCs and MoDCs (28), we determined the effect of viral infection on pDC numbers, a potential in-vivo source of IFN-α. We monitored the proportion of pDCs in PBMCs by flow cytometry following FMDV infection. In the naïve pigs, these cells ranged from 0.5–1.5% of the PBMC population. During infection the numbers progressively declined by days 3–5 PI, as shown in the representative dot plots of a representative animal (Fig. 5). By day 7 these cells had returned to pre-infection levels. This trend was observed in all animals analyzed for pDC numbers in the 7 d following infection.
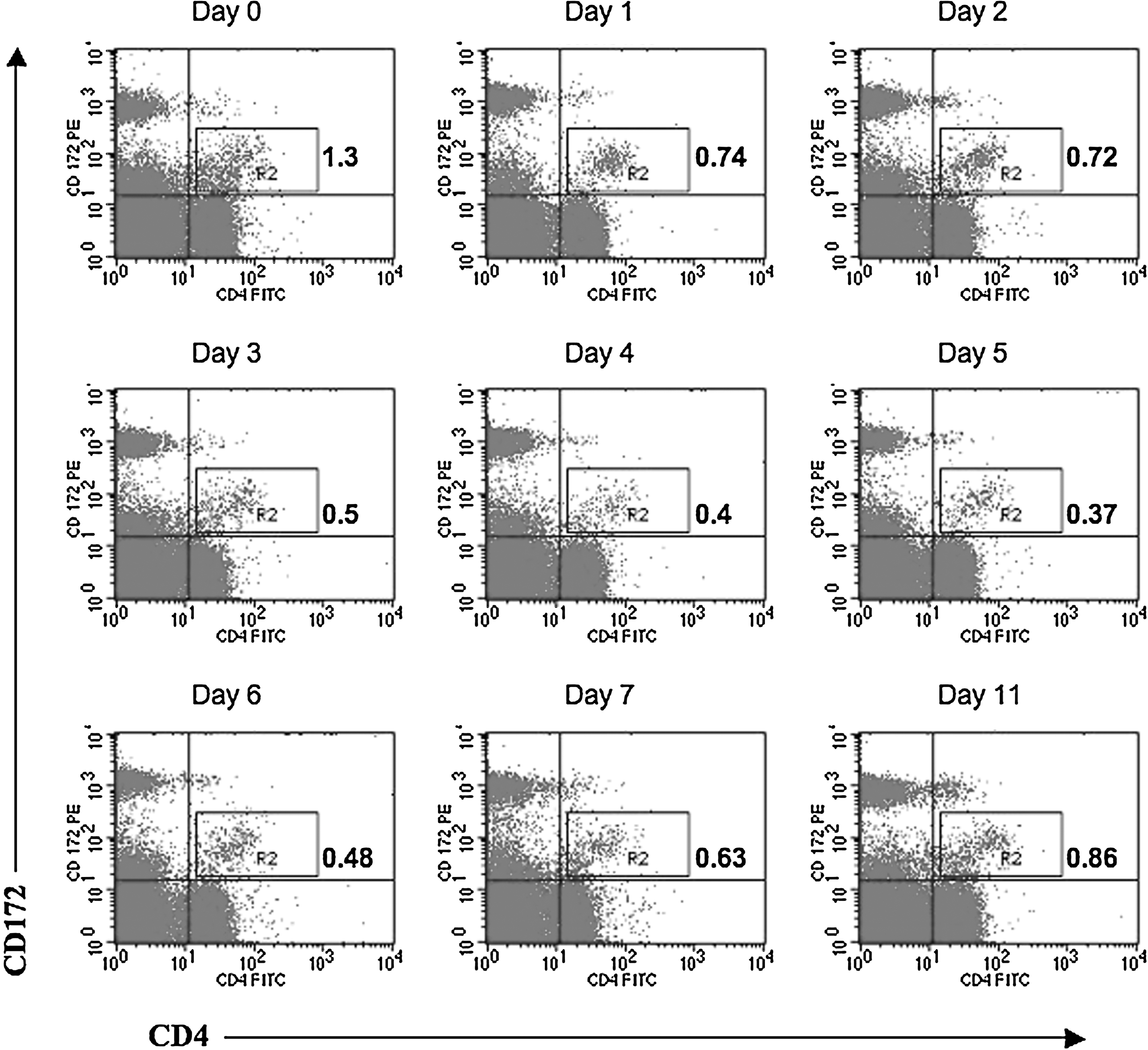
Decreased numbers of pDCs in FMDV-infected pigs. PBMCs isolated from pigs following FMDV infection were stained for CD4 and CD172 and analyzed by flow cytometry. The effect of infection on porcine pDC numbers identified in gate R2 as CD4+CD172low were monitored. Plots are representative of eight different experiments. Color image is available online at
Furthermore, previously swine pDCs were shown to produce large amounts of IFN-α in response to the TLR 9 agonist type A CpG 2216 (16). This was confirmed here by us (Fig. 4B). Thus, using ELISpot detection of cells secreting IFN-α following CpG 2216 stimulation, we found that the number of pDCs in the blood began to diminish on days 2 through 4, with the lowest percentage detectable on day 4 or 5 (Fig. 6A–H). This effect was seen regardless of virus serotype, and it began concurrently with the onset of viremia. We show individual animals analyzed in these experiments, given the variability in response in these outbred pigs. Nevertheless, this decline in pDCs responding to CpG 2216 was statistically significant (p < 0.01) on day 4 relative to day 0 for the virus strains O1C, A24, C-s8C1 PP1, and C-s8C1 PP2.

Decreased numbers of pDCs secreting IFN-α in response to CpG 2216 in FMDV-infected pigs. PBMCs isolated from pigs following FMDV infection were stimulated with CpG 2216, and the number of IFN-α spot-forming units was determined by ELISpot. (
We then measured IFN-α protein in supernatants of cultures of PBMCs stimulated with CpG 2216. These supernatants contained declining amounts of IFN-α, beginning by 1–3 days PI (Fig. 7 A–F), and nearly all animals showed a lack of IFN-α by days 3–5. The exceptions were the animals infected with strain C-s8C1, in which this effect occurred on day 2 in all animals tested. Recovery of IFN-α production varied, with some animals recovering rapidly and others showing a low response for 1–3 d. This observation could be due to the low numbers of pDCs in the PBMCs during acute FMDV infection. However, as shown in Fig. 8, the immunospot intensity was also low on days 2–4 PI, indicating that the amount of IFN-α secretion per responding cell was reduced during FMD. Thus, FMDV infection of pigs resulted in a reduction of both number and IFN-α responses by pDCs.
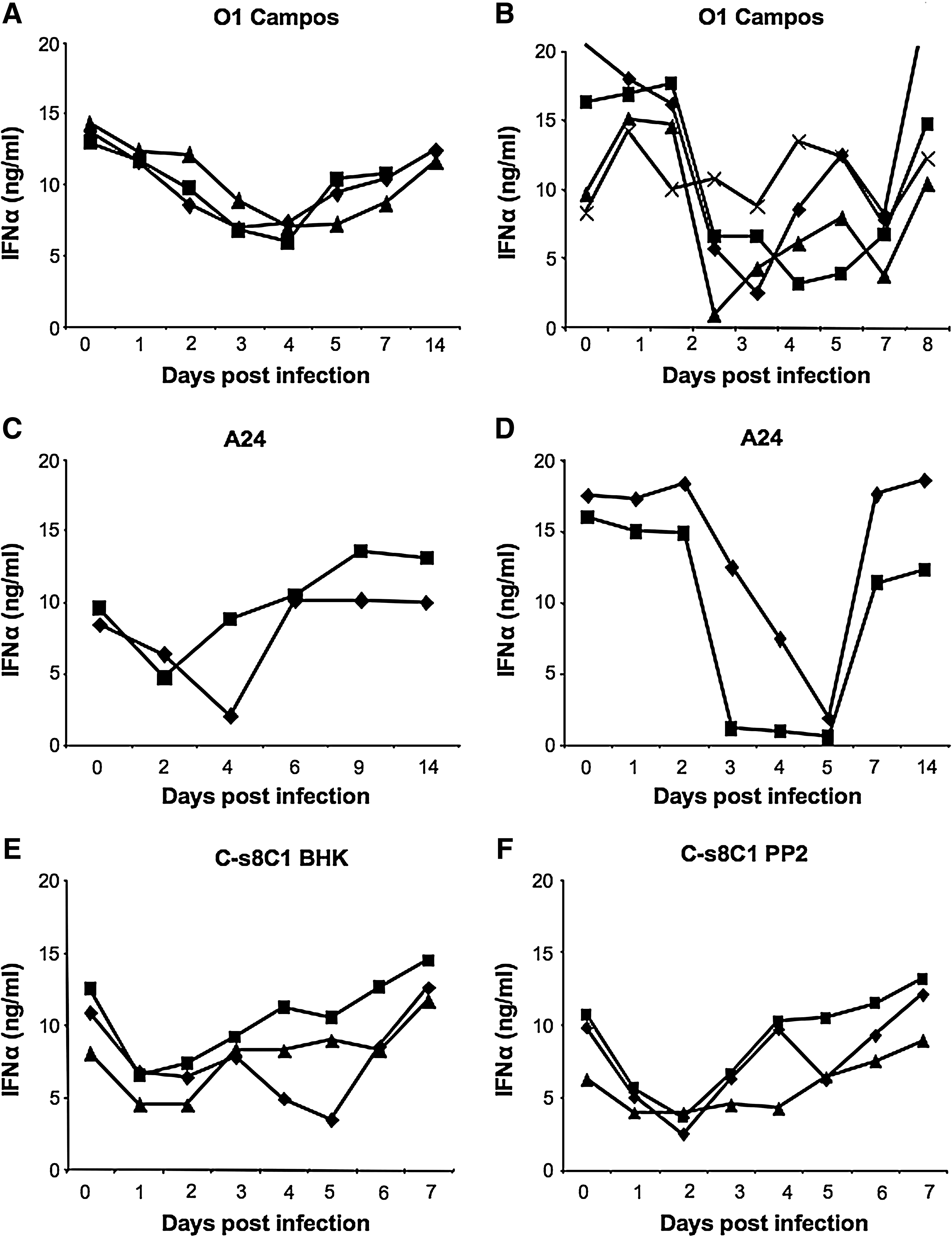
PBMCs from FMDV-infected pigs secrete less IFN-α in response to CpG 2216. PBMCs isolated from pigs following FMDV infection were stimulated with CpG 2216 and the supernatants were assayed for IFN-α by ELISA. (

Declining IFN-α spot intensity of pDCs from FMDV-infected pigs. PBMCs isolated from pigs following FMDV infection were stimulated with CpG 2216, and the number of IFN-α spot-forming units was determined by ELISpot. Photos of triplicate wells for each time point were taken on ImmunoSpot.
Discussion
Fever, loss of appetite, lameness, vesicles on the feet and snout, and early viremia on days 1–4 PI are common in FMDV infection of pigs, regardless of virus strain. In addition, lymphopenia has been identified as a hallmark of porcine (2,11,13), and to a lesser extent bovine (25), FMDV infection. However, the mechanism of the lymphopenia remains largely unknown. Apoptosis has been ruled out as being responsible for lymphocyte destruction (2,13), and in this and our previous reports (2,28) we could not isolate virus from lymphocytes, even at peak viremia. However, Diaz-San Segundo et al. reported isolation of virus from lymphocytes recovered from FMDV strain C-s8C1-infected pigs beginning on day 2 and extending until day 10 PI (13). Animals infected with this in-vitro-derived virus isolate were also reported to have viremia lasting more than 10 d, an uncommon finding among reports of porcine FMDV infection.
Most reports of swine infection show that upon needle inoculation of FMDV, viremia is cleared on day 4–5 PI (2,11,19,26,28,30,40). This could be attributable to the response of the different breeds of pigs used in the experiments by the different investigators, such as the Landrace-large white cross, as opposed to the Yorkshire-Landrace cross commonly used in our experiments. In addition, in-vitro propagation of FMDV alters its receptor recognition, and may widen its types of cellular targets (20). Of note is that Diaz-San Segundo et al. monitored viremia in euthanized animals only, as opposed to longitudinally monitoring the same animals over the 10-d period. All of these differences could account for their unique results.
The presence of IFN-α in serum has been linked to lymphopenia. In pigs infected with classical swine fever virus (CSFV), lymphopenia and a decline in pDC numbers correlates with the levels of IFN-α in the serum (37). Similarly, mice injected with poly I:C exhibit a lymphopenia linked to their serum IFN-α levels (22,24). In this article, the lowest lymphocyte and pDC numbers in the blood occurred when the serum IFN-α response was at its peak. Thus, the serum IFN-α response could partly account for the observed lymphopenia and reduction in number of pDCs, as is the case in CSFV infection (37). The IFN-α-induced lymphopenia is completely reversible and may be due to enhanced lymphocyte adhesion to endothelium of blood vessels during acute infection (24).
Viremia is rapidly cleared in FMDV-infected pigs, and there is no evidence of persistent infections (2). In this article, peak serum IFN-α occurred on days 2–3 PI, and preceded the rapid clearance of viremia by day 4. However, IgM, which is the first antibody isotype secreted in a primary response, was only detected in serum on day 7 (data not shown), after the viremia had resolved. In similar studies, neutralizing antibodies detected by plaque reduction assays were first detected on day 7 after inoculation (15). Therefore, other mechanisms in addition to neutralizing antibodies likely contribute to the early elimination of FMDV from the blood. Considering the high sensitivity of FMDV to IFN-α, the presence in serum of this cytokine reported here likely contributes to the early clearance of FMDV, by inducing an antiviral state in vivo. Such high serum levels of IFN-α have been shown to be protective against FMDV challenge (8). Therefore, the early IFN-α response renders virus-susceptible cells refractory to infection, thereby preventing further replication and spread of the virus within the host, and resulting in a rapid reduction of viremia.
Zhang et al. observed high levels of type I and II IFN mRNA in nasal-associated lymphoid tissues of cattle during the acute phase of FMD, and postulated that the inhibitory effects of these cytokines account in part for the mechanism by which FMDV is controlled in vivo (41). Similarly, an attenuated FMDV strain lacking the leader protease (LLA12) does not spread beyond the site of infection, due to a rapid local IFN-α response (5). It has been demonstrated that IFN-α activates expression of IFN-stimulated genes such as PKR and OAS, which results in the blocking of FMDV replication (7). Furthermore, it has been shown that this cytokine activates NK cells, which not only kill virus-infected cells, but also secrete cytokines such as IFN-γ that activate T and B cells (4,38). Furthermore, FMDV is sensitive to the antiviral activity of IFN-γ (26).
We have previously shown that upon exposure to FMDV in vitro, skin DCs secrete IFN-α and IFN-β (3). Similarly, porcine MoDCs propagated from PBMCs secrete IFN-α in response to FMDV (28). These data indicate that these DC populations may constitute the cellular sources of the serum IFN-α response to FMDV. Plasmacytoid DCs (pDCs) require FMDV complexed with immune serum for uptake and response to the virus in vitro (17). This would suggest that pDCs are an unlikely source of IFN-α in the early phase of infection in naïve animals (days 1–4). However, these cells might behave differently in vivo. Additional sources of IFN-α include macrophages and fibroblasts. A role for fibroblasts in the innate response to FMDV is suggested by the fact that various porcine kidney fibroblast cell lines (SK6, PK15, and IBRS2) secrete IFN-α when challenged with the virus (7,9).
However, the rapid progression of FMDV infection in pigs within the first 2 d of infection suggests the possibility of a window of opportunity for the virus to replicate. Either the virus downregulates the secretion of cytokines early after infection, delaying the attainment of sufficient levels of IFN-α in the serum, or high titers of virus are required to induce IFN-α secretion by the responding cells. The possibility of the former is suggested by the fact that skin DCs and MoDCs isolated from pigs 48 h post-FMDV infection show severely reduced secretion of IFN-α upon in-vitro stimulation (28).
This report provides more evidence for early modulation of the innate responses via the reduced number pDCs and the low IFN-α secretion of pDCs isolated from acutely infected pigs. Similar observations have been made upon analysis of infection with other porcine and human viruses. In addition to CSFV and FMDV infection, in-vitro infection of porcine pDCs with porcine circovirus type 2 results in inhibition of their response to stimulation with TLR 7 and 9 agonists. This is also observed in infection with pseudorabies virus and transmissible gastroenteritis virus. All of these effects are independent of viral replication (39). In humans, a blunted response of blood pDCs and reduced cell numbers are evident in dengue virus infection, although these cells are not productively infected with dengue virus and still remain sensitive to stimulation with CpG 2216 (32). Furthermore, severe infection with herpes simplex virus type 2 leads to a rapid loss of pDC and NK cells (10). Similarly, primary infections with human immunodeficiency virus are characterized by impaired type I IFN production and low counts of pDCs (23). To date we have not demonstrated what causes the low numbers of pDCs during FMDV infection, but one attractive hypothesis is the possibility that these cells migrate from the bloodstream into sites of inflammation.
Conclusion
In summary, the data presented here are consistent with the contribution of IFN-α to the early resolution of viremia in FMDV-infected pigs, before the detection of antibodies to the virus. However, this response is modulated by the virus, creating a narrow but sufficient window of opportunity for replication and shedding. This article lends support to ongoing research to develop systems that induce interferons to augment the host response as an intervention strategy during outbreaks of FMD.
Footnotes
Acknowledgments
This work was supported by Current Research Information System (CRIS) #1940-32000-052-00D (W.T.G.) from the Agricultural Research Service, U.S. Department of Agriculture (USDA), and an interagency agreement (no. 60-1940-8-037) between the Department of Homeland Security, Science and Technology Directorate, and the USDA (W.T.G.). C. Nfon was the recipient of a PIADC Research Participation Program fellowship, administered by the Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and the USDA. We would like to thank Drs. Noami Sevilla and Fayna Diaz-San Segundo (Instituto Nacional de Investigacion y Technologia Agraria (INIA), Madrid, Spain) for providing the FMDV strain C-s8C1 for this analysis. We would also like to thank Mr. Geoffrey Ferman and Ms. Mital Pandya for their technical assistance, and the animal care staff at the Plum Island Animal Disease Center for their professional support and assistance.