
Abstract
Chickens infected with Marek's disease virus (MDV) become lifelong carriers regardless of their susceptibility to clinical disease. Therefore various viral immune-evasive mechanisms must play a role in MDV-host interactions. MDV has previously been shown to influence the expression of major histocompatibility complex (MHC) class II molecules. However, little is known about the underlying mechanisms of this phenomenon. In the present study, we studied the effect of MDV infection on the expression of several genes associated with IFN-γ-inducible MHC class II expression at 4, 7, 14, and 21 days post-infection (dpi). There was a significant (p ≤ 0.05) downregulation of MHC class II β chain expression throughout the experiment, while other components of the MHC class II heterotrimer (i.e., α chain and the invariant chain) were significantly downregulated only at 4 and 21 dpi. Furthermore, the expression of components of the IFN-γ-receptor complex was significantly downregulated at 4 dpi. In contrast, a number of other IFN-γ-signaling molecules, including signal transducer and activator of transcription 1 (STAT1), interferon-responsive factor 1 (IRF-1), and class II transactivator (CIITA) were significantly upregulated at most time points. The results of this study shed light on the possible mechanisms by which MDV may evade host immunosurveillance.
Introduction
Given its central role in modulating the host immune response, major histocompatibility complex (MHC)-mediated antigen presentation has been a target for immune-evasive mechanisms by many viruses. For instance, a number of viruses including human cytomegalovirus, human immunodeficiency virus, adenoviruses, and Epstein-Barr virus (EBV), interfere with the MHC class I-mediated antigen presentation pathway by inhibiting expression of multiple interferon (IFN)-γ-inducible genes (4). Similarly, virus-mediated interference also occurs at several levels other than gene expression, such as antigen processing, MHC class I molecule assembly, and surface expression of MHC molecules (5,22). With respect to MDV, however, there are only limited data available on its interference with MHC class I-mediated antigen presentation. For instance, MDV appears to downregulate the surface expression of MHC class I molecules in infected cells in vitro (17,20).
Similarly to MHC class I, many types of viruses including members of herpesviruses, poxviruses, and paramyxoviruses can also manipulate MHC class II-mediated antigen presentation (16). While almost all herpesviruses studied to date cause downregulation of MHC class II-mediated antigen presentation, the influence of MDV on the MHC class II seems to vary depending on the experimental model. For instance, Niikura et al. showed that MHC class II surface expression is enhanced in MDV-infected cells derived from chickens bred as a cross between MD-susceptible 15I5 and 71 lines infected with the virulent Md11 strain of MDV (24). Furthermore, Dalgaard et al. (2009) reported that at 4 wk post-infection, the virulent GA strain of MDV upregulates MHC class II expression on different subsets of T cells from chickens carrying the B19 and B21 MHC haplotypes, which are associated with susceptibility to and resistance to MD, respectively (11). However, in the same study it was shown that MDV downregulates MHC class II expression on T cells at 1 wk post-infection (11). In contrast to the observations that MDV enhances MHC II expression, Sarson and colleagues analyzed spleen transcriptome at 4, 7, 14, and 21 dpi, using a chicken immune-specific microarray analysis, and showed a significant downregulation of MHC class II β chain (B-LB) gene expression at multiple sampling points in spleen tissues of B19 and B21 lines of chickens infected with virulent JM16-MDV (28). In another experiment examining gene expression at similar sampling points using outbred specific pathogen-free chickens, a significant downregulation of MHC class II invariant chain (Ii) gene expression was noted in chicken spleens at 7 and 14 d after MDV infection (27). Furthermore, a 2D-gel electrophoresis analysis of very virulent RB1B-MDV-infected chicken spleen proteome at 7, 14, and 21 dpi, revealed a significant downregulation of the MHC class II α chain at 7 dpi (29). Based on the above observations, it may be concluded that both the genetic background of the host and the strain of the virus can affect MDV's influence on MHC expression.
MHC class II expression is mainly regulated at the level of transcription by a key coactivator known as MHC class II transactivator (CIITA) (7,30). CIITA is constitutively expressed in professional antigen-presenting cells (APCs), and is inducible in many other cell types following stimulation by IFN-γ (30). Consequently, the MHC class II expression pathway induced by IFN-γ has been a target for a number of viruses (9).
Although the IFN-γ-inducible MHC class II expression pathway has not been well studied in chickens, it has been well characterized in mammalian species (1,8,30). Briefly, IFN-γ interaction with its receptor 1 (IFN-γR1) homodimer on the cell surface causes recruitment of another receptor subunit, IFN-γR2. The cytoplasmic tails of these two subunits are associated with Janus kinases (JAK)-1 and JAK-2, respectively. Approximation of JAK-1 and JAK-2 causes phosphorylation of JAK-2, JAK-1, and IFN-γR1. This eventually results in the formation of phosphorylated homodimers of signal transducer and activator of transcription 1 (STAT1), which is subsequently channeled to the nucleus to activate transcription of various IFN-γ-inducible genes, including CIITA. The promoter for CIITA consists of three regulatory elements: gamma-activation sequence box, E box, and interferon-responsive factor (IRF) box. Interaction of these three elements with STAT1 homodimer, upstream stimulatory factor (USF)-1, and IRF-1, respectively, promotes expression of CIITA, which interacts with several other regulatory molecules occupying class II promoter to form a multi-subunit transcriptosome complex. This complex would ultimately lead to transcription of class II genes.
Motivated by our previous observations of downregulation of different components of MHC class II molecules in MDV-infected chickens, we endeavored to study the effect of MDV on the expression of genes encoding MHC class II molecules, as well as different components of the IFN-γ-inducible JAK/STAT-mediated MHC class II expression.
Materials and Methods
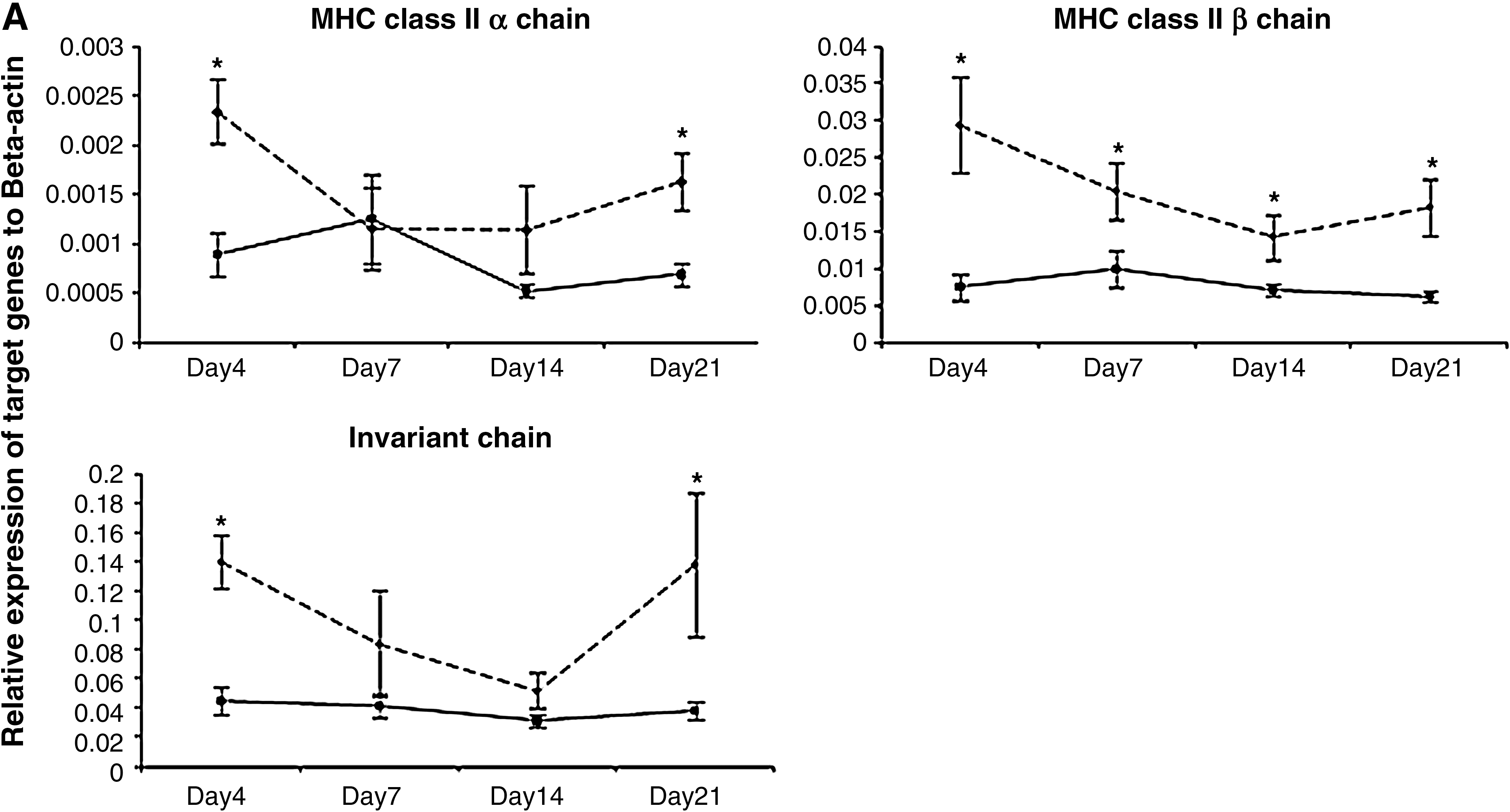
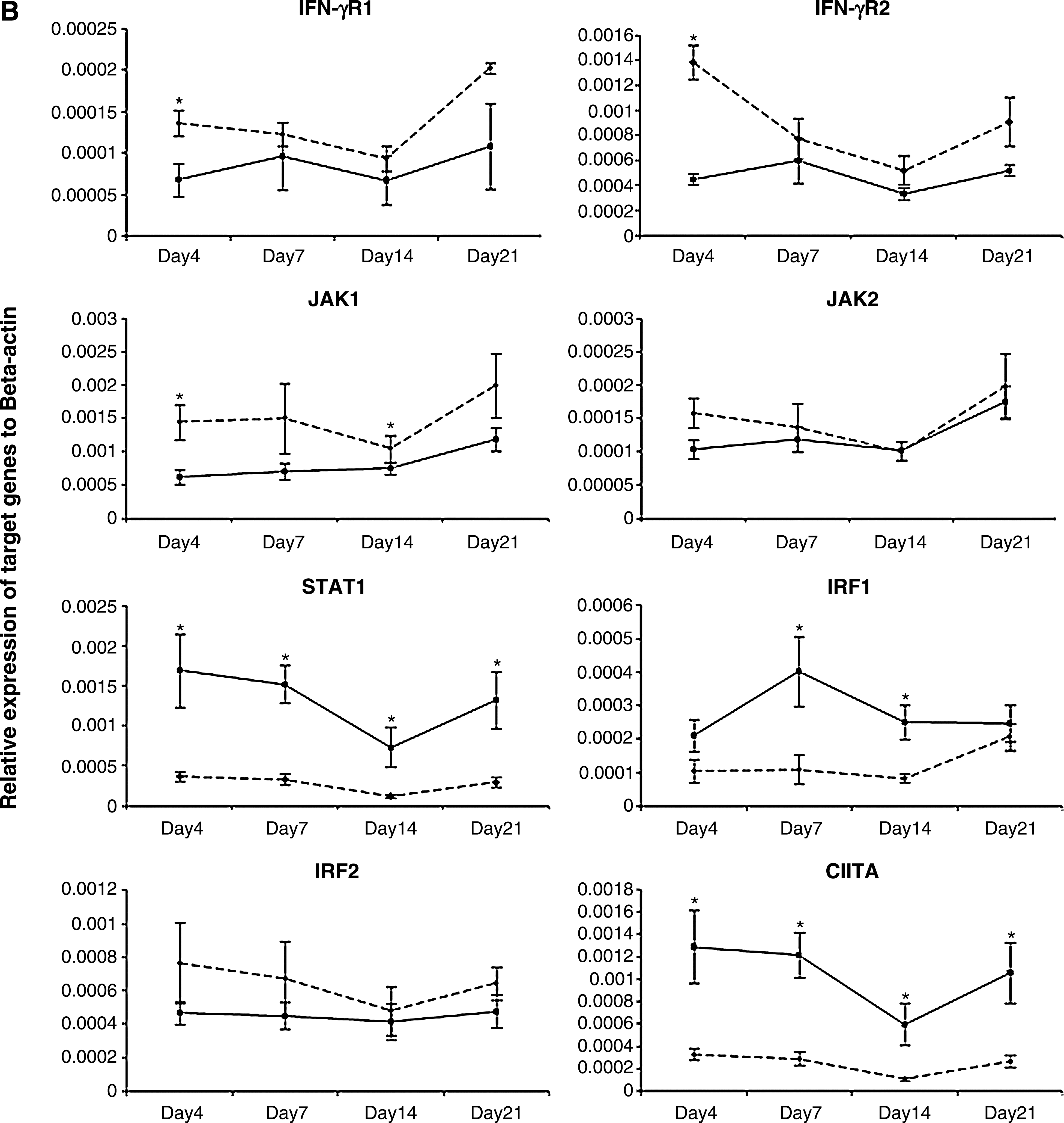
In the current study, 25 5-day-old outbred SPF Leghorn chickens (obtained from the Canadian Food Inspection Agency, Ottawa, Ontario, Canada) were infected intra-abdominally with 250 plaque-forming units of the very virulent RB1B strain of MDV per bird (passage 9; kindly provided by Dr. K.A. Schat, Cornell University, Ithaca, NY). Another group of chickens (n = 16) was kept under similar conditions as uninfected age-matched controls. Spleens from at least six and four randomly selected birds from the MDV-infected and control groups, respectively, were harvested at 4, 7, 14, and 21 days post-infection (dpi), representing different phases of MD pathogenesis (i.e., the early cytolytic phase, the transition from the cytolytic to the latent phase, and from the latent phase to the transformation phase). Subsequently total RNA was extracted from each spleen tissue sample using Trizol (Invitrogen Canada Inc., Burlington, Ontario, Canada), followed by cDNA synthesis using 1 μg of RNA from each sample using oligo(dT)12–18 primers (SuperScript™ First-Strand Synthesis System; Invitrogen Life Technologies, Carlsbad, CA) according to the manufacturer's instructions. Target gene fragments were PCR amplified from cDNA using gene-specific primers (Table 1), cloned into the pDrive cloning vector (Qiagen® PCR Cloning Kit; Qiagen Inc., Valencia, CA), and used as standard controls to generate standard curves by qRT-PCR analysis as described previously (2). Relative expression of each target gene in cDNA samples from all sampling time points was analyzed in a single real-time RT-PCR assay in 384-well plates using a LightCycler 480 instrument (Roche Diagnostics GmbH, Mannheim, Germany). A previously published set of primers was used to amplify β-actin as the reference gene (2). The expression of each target gene was calculated relative to the expression of the β-actin gene in the same sample using Pfaffl's formula (26). The average relative expression of each gene (Fig. 1) at a given time point was compared between infected and control groups using the Student's t-test, and differences were considered significant at p ≤ 0.05.
(
Results and Discussion
Based on our observations, MHC class II β chain expression was significantly lower (p ≤ 0.05) in infected chickens throughout the experimental period. With respect to the MHC class II α chain, the expression levels were significantly lower in infected chickens only at 4 and 21 dpi. MDV infection also caused downregulation of Ii expression. Accordingly, we observed that changes in the expression of Ii were significant (p ≤ 0.05) at 4 and 21 dpi. These findings in the context of MD are in agreement with our previous observations (27 –29). For instance, MDV infection downregulates the expression of the MHC class II α chain protein in spleen at 7 dpi in vivo (29). Furthermore, using a microarray approach, it was shown that MDV infection causes significant downregulation of MHC class II β chain gene expression in both MD-resistant B21 and MD-susceptible B19 chickens at almost all sampling time points (27). Similarly, in another microarray study it was demonstrated that MDV infection causes a significant downregulation of Ii gene expression in chicken spleens, at least at 7 and 14 d after MDV infection (28). A recent study has also shown downregulation of MHC class II on T-cell subsets and B cells at 1 wk and 4 wk post-infection, respectively (11). Collectively, these findings may indicate interference of MDV with MHC class II-mediated antigen presentation.
Although there are limited amount data available about MDV's interference with MHC class II-mediated antigen presentation, the use of this as a means of concealing infected cells from host immunosurveillance by many other viruses is well documented (16). There are two main strategies of viral interference with MHC class II-mediated antigen presentation: by negatively affecting MHC class II gene expression, or by interfering with surface expression of MHC class II molecules. While there are a number of examples of viruses that interfere with the surface expression of MHC II, the most common strategy appears to be interference with the IFN-γ-inducible MHC II expression pathway (16). Infections by most viruses induce expression of IFN-γ in the host (14). Similarly, MDV induces the expression of IFN-γ in vitro and in vivo (3,18,25,33), which may lead to the induction of a number of IFN-γ-inducible genes, including various components of MHC class II. Therefore we explored whether the downregulation of MHC-II components is caused by interference of MDV with the IFN-γ-inducible MHC class II expression pathway.
Accordingly, relative expression of eight genes that encode various molecules involved in the IFN-γ-inducible JAK/STAT-mediated CIITA expression pathway was quantified. Among them, the expression of both chains of IFN-γ receptors (IFN-γR1 and IFN-γR2), as well as three other genes encoding JAK-1, JAK-2, and IRF-2 were relatively lower in expression in spleens of infected chickens compared to spleens of uninfected birds, at almost all time points (Fig. 1). However, only changes in IFN-γR1 and IFN-γR2 expression at 4 dpi, and JAK-1 expression at 4 and 14 dpi, were statistically significant (p ≤ 0.05). Although the expression levels were relatively lower in MDV-infected birds, changes in the expression of JAK-2 and IRF-2 were not significant at any time point (p ≥ 0.05). In contrast, infected spleen tissues showed higher levels of STAT1, IRF-1, and CIITA expression at all time points. While changes in STAT1 and CIITA were statistically significant (p ≤ 0.05) at all time points, expression of IRF-1 was only significantly different at 7 and 14 dpi.
Although this is the first report of interference of IFN-γ receptor complex gene expression in the chicken in the context of MDV infection, similar observations have been reported in the context of a number of other viruses, including herpes simplex virus (HSV), cytomegalovirus (CMV), EBV, varicella-zoster virus (VZV), and adenoviruses. For instance, Joseph and Look (2001) showed that adenovirus type 5 infection in respiratory epithelial cells inhibits the IFN-γ-inducible gene expression pathway by directly targeting the IFN-γ-receptor complex, particularly by inhibiting its assembly and phosphorylation of its components, as well as decreasing the expression of the IFN-γR2 gene (19). Furthermore, EBV immediate early protein, BamHI Z fragment leftward open reading frame 1 (BZLF1), interferes with IFN-γ signaling by preventing activation and/or nuclear translocation of several components of the pathway, and causing downregulation of IFN-γR1 expression at the mRNA level (23). Importantly, the critical role of IFN-γ receptors in immunity against various intracellular pathogens has been highlighted by the exacerbated susceptibility of patients with genetic defects in IFN-γR1 and IFN-γR2 to pathogens such as CMV, HSV, and VZV (12).
In the present study, analysis of expression of genes encoding downstream components of the IFN-γ-signaling pathway revealed that several genes were upregulated despite the downregulation of the IFN-γ-receptor complex in MDV-infected chicken spleen tissues. Notably, among them STAT1 expression was higher in infected tissues at all sampling time points. Furthermore, IRF-1 expression was also significantly enhanced in spleens of infected chickens at 7 and 14 dpi. However, there was no significant change in IRF-2 expression. In contrast to our present observations, these genes have been negatively affected by a number of viruses that target IFN-γ signal transduction. For instance, in a microarray analysis of gene expression of human papillomavirus type 31-infected human keratinocytes, STAT1 was significantly downregulated in infected cells (10). However, none of the other components of the pathway were significantly affected. Furthermore, Ebola virus infection has been shown to reduce the IFN-γ-inducible genes, including IRF-1, likely by inhibiting JAK-1 and STAT1 (9). On the other hand, as STAT1 and IRF-1 are key components of the type I interferon (IFN-α/β) signaling pathway, elevated expression of these genes may indicate active signal transduction mediated by type I interferons (14). Furthermore, cytokines other than interferons may also play a role in this context. For example, TNF-α, IL-1, and IL-6 may also upregulate IRF-1 expression (13,15). However, the role of these elements in the elevated expression of STAT1 and IRF-1 seen in the present experiment needs to be further studied.
Interestingly, in contrast to our expectation based on the lower expression of both MHC class II α and β chains, as well as Ii, and also supported by the lower expression of components of the IFN-γ-receptor complex in infected chickens, CIITA transcripts were significantly increased at all time points in the present study. This is of particular interest, as the expression of CIITA usually parallels that of MHC class II molecules, at least in mammals (30). On the other hand, CIITA upregulation in our study coincided with the increased expression of STAT1 and IRF-1 genes. STAT1 and IRF-1, together with two other transcription factors, IRF-2 and USF-1, interact with the P4 promoter of CIITA to mediate its IFN-γ-inducible expression (30,32). The expression of CIITA is primarily regulated at the transcriptional level (30). Therefore, although there is no experimental evidence, it is plausible that the reduced expression of MHC class II genes could initiate a positive feedback mechanism, leading to elevated expression of CIITA. On the other hand, emerging evidence suggests that the function of CIITA is tightly regulated at the post-translational level. For instance, prostaglandin-induced protein kinase A inhibits IFN-γ-induced MHC class II mRNA expression through post-translational modification of CIITA in both mouse and human cells (21). Furthermore, formation of the active form of CIITA is regulated by phosphorylation of serine residues in its amino terminus by extracellular signal-regulated kinase (ERK) 1 and 2 (31). Therefore it is also possible that MDV interferes with CIITA at the post-transcriptional level. This hypothesis, however, was not tested in this study.
Here we have shown that MDV significantly downregulates expression of several components of the MHC class II antigen-presenting molecules in infected chickens. Furthermore, expression of a number of genes encoding components of the IFN-γ-inducible JAK/STAT-mediated induction of CIITA is also significantly affected by MDV infection. While this is the first report of the analysis of expression of IFN-γ signal transduction in the context of MD in chickens, the findings detailed here warrant further exploration into the possible mechanisms by which MDV evades host immunosurveillance.
Footnotes
Acknowledgments
This research was funded by the Natural Sciences and Engineering Research Council of Canada and the Poultry Industry Council. Niroshan Thanthrige-Don is a recipient of a Commonwealth Graduate Scholarship granted by Foreign Affairs and International Trade Canada.
Author Disclosure Statement
No competing financial interests exist for any of the authors of this manuscript.