
Abstract
Highly pathogenic avian influenza H5N1 viruses are capable of causing poultry epidemics and human mortality. Vaccines that induce protective neutralizing antibodies can prevent outbreaks and decrease the potential for influenza A pandemics. Identifying unique H5N1 virus-specific HLA class II-restricted epitopes is essential for monitoring cellular strain-specific immunity. Our results indicate that 80% of the 30 study participants who were inoculated with an H5N1 vaccine produced neutralizing antibodies. We used intracellular cytokine staining (ICS) to screen and identify six DR1501-restricted H5N1 virus epitopes: H5HA148–162, H5HA155–169, H5HA253–267, H5HA260–274, H5HA267–281 and H5HA309–323. Tetramer staining results confirmed that two immunodominant epitopes were DR1501-restricted: H5HA155–169 and H5HA267–281. Both are located at the HA surface and are highly conserved in currently circulating H5N1 clades. These results suggest that a combination of ICS and tetramer staining can be used as a T-cell epitope-mapping platform, and the identified epitopes may serve as markers for monitoring vaccine efficacy.
Introduction
Hemagglutinin (HA) proteins form spikes and mediate influenza virus binding to receptors, resulting in membrane fusion and viral entry. Specifically, the HA1 domain contains major antigenic regions for neutralizing antibodies. Protection against influenza infection is conferred via neutralizing antibodies and stimulated T lymphocytes for the surface proteins HA and neuraminidase (NA) (21,31). The HA2 domain, which consists of internal envelope parts, has greater similarity among multiple influenza A virus subtypes (13,31). Immune T cells can be elicited by natural infections by influenza viruses, especially via HA1 antigen recognition (14). One research team has demonstrated that T cells stimulated with epitopes in the HA2 domain of the H5N1 virus also recognize H1N1 and H3N2 viruses (26). This suggests that B-cell and T-cell epitopes that are specific to influenza virus subtypes exist in the HA1 domain.
Innate and adaptive immune responses are elicited by influenza virus infections or vaccinations. To monitor B- and T-cell stimulation and activation, it is essential to understand the protective capabilities of human immune responses to H5N1 viruses. T-cell immunity is correlated with antibody production and influenza virus clearance (1,28); individuals with better T-cell responses suffer from milder forms of the disease, are more likely to quickly recover, and are less likely to die or spread the virus (10). The MHC II complex, associated with the presentation of specific peptides to CD4+ T lymphocytes, plays a key role in connecting innate and adaptive immunity (3). Naturally-processed peptides that bind to MHC class II molecules have between 13 and 25 residues, with an average length of 15 residues (8). CD4+ T cells also act as effector cells for antiviral responses by killing infected cells and secreting cytokines such as IFN-γ and TNF-α (29). Evaluating antigen-specific CD4+ T-cell responses at the individual cell level supports a wide range of potential applications, ranging from studies of pathogenesis to evaluations of the efficacy of vaccines or immunotherapeutic agents (30).
Many of the most commonly used methods for analyzing T-cell function [including measurement of cytokine secretion, [3H]-thymidine proliferation, 51Cr-release, and limiting dilution assays (12)] are both time-consuming and non-quantitative. ELISPOT (27) and intracellular cytokine assays (12) have recently been developed for the purpose of directly evaluating Ag-specific CD4+ T-cell function. Within the past 10 years, HLA class II tetramer technology has been developed for detecting Ag-specific CD4+ T cells, based on specific interactions between MHC peptides and the T-cell receptors of CD4+ T cells (9). Due to low CD4+ T-cell frequency, this approach requires additional Ag-specific expansion prior to staining with class II tetramers in vitro (6). MHC class II tetramers, which are widely used in basic and clinical immunological research for identifying antigen-specific CD4+T cells, offer valuable information for vaccine development and efficacy evaluation (9).
Each year the World Health Organization makes influenza virus recommendations for vaccine preparation. Due to antigenic drift, it is difficult to develop influenza A virus vaccines that provide long-lasting immunity (4). In 2007, the U.S. Food and Drug Administration approved that country's first H5N1 influenza vaccine. Created by Sanofi Pasteur, the vaccine will not be sold commercially, but will be held in a Strategic National Stockpile of emergency medical countermeasures (5). GlaxoSmithKline (GSK) has received permission from European Union authorities to market an H5N1 adjuvanted pre-pandemic vaccine named Prepandrix™ (5). To date, information on the human CD4+ T-cell epitopes of avian H5N1 hemagglutinin proteins is limited due to antigen or vaccine availability, as well as the small number of human cases caused by the avian H5N1 influenza virus.
Our goal in this study was to identify important epitopes in the HA1 region of the H5 HA protein recognized by CD4+ T lymphocytes. Since HA2 is more conserved, there is greater potential for identifying specific H5 HA1 epitopes for monitoring vaccine efficacy. We recruited 30 participants who were given injections of the GSK H5N1 vaccine. After titrating anti-H5N1 neutralizing antibodies in the vaccinees, we selected the DR1501 allele for identifying restricted epitopes on H5 HA proteins. Our reasoning behind this decision was the large proportion of Minnan Taiwanese who carry this allele. We performed T-cell epitope mapping with peptides covering the entire H5 HA1 domain, and then assayed their individual capabilities to stimulate specific T cells in peripheral blood mononuclear cells (PBMCs) from vaccinated DR1501-restricted participants. Since tetramer-guided epitope mapping requires large quantities of tetramer reagents, responses were determined by intracellular cytokine staining (ICS), followed by flow cytometry. Our results indicate that H5HA148–162, H5HA155–169, H5HA253–267, H5HA260–274, H5HA267–281, and H5HA309–323 contain CD4+ T-cell epitopes. DR1501 MHC class II tetramer staining was conducted to confirm the presence of these epitopes. Our results indicate that HA155–169 and HA267–281 are DR1501-restricted CD4+ T-cell epitopes of H5 HA proteins. We believe this information is useful for researchers working on T-cell immune response evaluation platforms and H5N1 vaccine development.
Materials and Methods
Participants and HLA typing
All 30 participants were identified as Taiwanese of Minnan ancestry. After signing informed consent documents approved by the Institutional Review Board of the National Yang-Ming University, they were inoculated with the GSK H5N1 vaccine. HLA-DR genotypes were determined by low-resolution class II typing using polymerase chain reaction sequence-specific oligonucleotide probes (PCR-SSOP), followed by high-resolution class II typing using DNA amplification and sequencing with allele-specific primers. Participants with DR1501 alleles were selected because of their higher antibody titers, and the general prevalence of the DR1501 allele among all Minnan Taiwanese.
ELISA assays
To evaluate H5N1 vaccine efficacy, ELISA plates coated with H5N1-RG viruses were used to validate the humoral response as previously described (31). Briefly, 96-well microplates (SpectraPlate™-96 HB; PerkinElmer, Waltham, MA) were coated with H5N1-RG strain viruses (HA titers 1:512) at a 1:400 dilution in PBS buffer and held at room temperature for 1 h. After washing five times with 0.05% PBST (PBS buffer containing 0.05% Tween 20), the plates were blocked with 5% milk in 0.05% PBST and held overnight at 4°C. Next, paired sera from each vaccinee were diluted 100-fold with 5% milk in 0.05% PBST, added to each well, and incubated at 37°C for 1 h. After washing five times with 0.05% PBST, 100 μL of 1:4000 diluted anti-human IgG Ab (Amersham Biosciences, Piscataway, NJ) conjugated with horseradish peroxidase (HRP) was added to each well and incubated at 37°C for 1 h. After five extensive washings with PBST, HRP-conjugated anti-mouse IgG (1:4000 dilution; Amersham Biosciences) was added prior to another hour of incubation at 37°C, followed by incubation with 200 μL substrate (0.015% o-phenylenediamine dihydrochloride; Sigma-Aldrich, St. Louis, MO) for 30 min at 37°C. The reactions were stopped by the addition of 3 N HCl; absorbances were measured with a Labsystems Multiskan Ascent Autoreader spectrophotometer (model 354; Thermo Labsystems, Helsinki, Finland) at 492 nm. The cut-off value was set at mean plus 3 standard deviations of pre-immune or non-vaccinated sera. Absorbance above the cut-off value was viewed as positive.
PBMC isolation
PBMCs from DR1501-carrying participants were purified and isolated as previously described (25). Briefly, blood was collected in vacuum tubes (10 mL blood per tube) containing K3 EDTA (ethylenediaminetetraacetic acid) as an anticoagulant. The tubes were centrifuged at 1800 rpm for 15 min at room temperature to separate serum, buffy coat, and RBCs. To inactivate complement, serum from each tube was collected and incubated at 56°C in a water bath for 30 min prior to storage at −20°C. The remaining blood parts were mixed with equal volumes of sterile PBS and added to a Ficoll-Paque™ PLUS centrifuge (GE Healthcare, Piscataway, NJ) at a 2:1 ratio of blood/PBS mixture to Ficoll. Centrifugation (3000 rpm for 30 min) was performed at room temperature; the buffy coat was drawn out and washed twice with ice-cold PBS at 1800 rpm at 4°C for 7 min. PBMCs were cultured in RPMI 1640 medium (Gibco, Carlsbad, CA) containing 10% FBS for use in our experiments.
H5N1 influenza peptides
Individual peptides were 15 aa long, with 8 aa overlaps between adjacent peptides. All peptides used in this study were divided into nine pools of five consecutive peptides and one pool of four peptides. Peptide sequences are shown in Table 1.
CD8+ T-cell depletion
Previous studies indicate that activated CD8+ T cells inhibit the ability of CD4+ T cells to proliferate in vitro (34), thus making it necessary to deplete CD8+ T cells prior to antigen-specific CD4+ T-cell stimulation. We coated 10-cm dishes with 22.5 μL goat anti-mouse IgG antibodies (Jackson ImmunoResearch, West Grove, PA) in 10 mL 1× HBSS at a concentration of 5 μg/mL and held them overnight at 4°C. The plates were washed twice with 10 mL 1× HBSS containing 2% FBS (Gibco), followed by the addition of 20 mL 1× HBSS containing 2% FBS for blocking at 4°C for 1 h. After one wash with PBS, 1–1.5 × 107 purified PBMCs in 10 mL OKT8 supernatant from a hybridoma cell line secreting anti-CD8 antibodies were incubated at 4°C for 1 h and washed twice with 1× HBSS containing 2% FBS, and spun at 1800 rpm for 7 min at 4°C. The PBMCs were resuspended with 1× HBSS containing 2% FBS at a concentration of 107 cells/mL, and added to 10 cm dishes coated with goat anti-mouse IgG antibodies. These plates were incubated at 4°C for 3 h and gently shaken once per hour. Cell-containing suspensions were collected in tubes and centrifuged at 1800 rpm for 7 min at 4°C. The cells were resuspended in RPMI 1640 medium containing 10% FBS and 1% P/S.
Antigen-specific CD4+ T-cell stimulation and ICS
CD8+ T-cell-depleted PBMCs (5 × 106/well) were expanded by stimulating pooled or individual peptides (10 μg/mL) co-cultured in complete RPMI 1640 medium at 37°C. Medium was refreshed with complete RPMI 1640 containing IL-2 (2 ng/mL) once every 2 d, starting on day 4. All cells were collected on day 13 for intracellular cytokine staining.
ICS was used to evaluate specific CD4+ T-lymphocyte activation, using previously described procedures (12). Briefly, peptides (10 μg/mL) were incubated with expanded CD8+-depleted PBMCs at 37°C for 1 h prior to the addition of brefeldin A (10 μg/mL), and then incubated for another 5 h at 37°C. Cells were harvested and stained with FITC-conjugated anti-CD4 MAb or FITC-conjugated mouse IgG1 and κ isotype control for 30 min at 4°C, then fixed with 4% paraformaldehyde at room temperature for 10 min. After one wash with ice-cold PBS, the cells were incubated with SAP buffer containing PE-conjugated anti-IFN-γ MAb or PE mouse IgG1 and κ isotype control for 45 min at room temperature. After extensive washing with ice-cold PBS, these cells were used for flow cytometry analysis.
Tetramer production and staining
DR1501 tetramers were kindly provided by Dr. Eddie James of the Benaroya Research Institute at Virginia Mason Hospital, Seattle, Washington. The details of DR15 tetramer preparation were described previously (7,19,22). Briefly, the extracellular coding region for the DR1501 chain appended to the acidic leucine zipper motif by using the PCR-mediated splicing overlap technique was cloned into a chimeric cassette. A biotinylation-specific sequence was cloned into the 3′-end of the DR1501/leucine zipper cassette. This chimeric cDNA was subcloned into the Cu-inducible pRmHa-3 Drosophila expression vector. Furthermore, the MHC class II complexes were produced by cotransfection with DR α and DR β expression vectors into Schneider S-2 cells and induced by CuSO4. Subsequently, soluble DR1501 tetramers were harvested, concentrated, and by biotinylated with the Bir A enzyme (Avidity Biotechnologies, Denver, CO). After 13 d of incubation at 37°C, 200 μL of resuspended cells (1 × 105 cells per 50 μL) were stained with 4 μL of PE-conjugated peptide-loaded tetramers for 60 min at 37°C, then stained with 3 μL FITC-conjugated CD4 MAbs (BD Pharmingen, Franklin Lakes, NJ) on ice, and analyzed by flow cytometry (105 cell sorting).
Results
HLA types among the 30 participants inoculated with the H5N1 vaccine were identified by PCR-SSOP (Table 2). To determine their specific post-vaccination humoral immunity against H5N1 viruses, we used an H5N1-RG virus-coated ELISA to quantify anti-H5N1 antibodies. Our results indicate that the large majority (24/30; 80%) of the vaccinees were H5N1-seropositive (Table 2). Considering participant availability and HLA prevalence among the vaccinees, we selected the HLA DR1501 allele to map significant T-cell epitopes specifically presented by the class II HLA DR1501 complex.
+, positive results; −, negative results.
ICS assays were used to screen for potential HLA DR1501-restricted T-cell epitopes in the HA proteins of H5N1 viruses. Peptides covering the entire H5 (HA1) hemagglutinin protein in the A/Vietnam/1203/04 virus were grouped into the 10 pools shown in Table 1. After co-culturing with these pooled peptides, CD8-depleted PBMCs were intracellularly stained for IFN-γ. According to results from primary IFN-γ screening and the CD4+ double-staining of T lymphocytes by peptide stimulation in a representative DR1501 participant, the number of T cells stimulated by pools 5, 8, and 9 were at least double those of the DMSO co-cultured control cells (Fig. 1A and B). We also identified which peptides in each pool were DR1501-restricted T-cell epitopes of H5 hemagglutinin proteins. Our results indicate that peptides 22 and 23 in pool 5; 37 and 39 in pool 8; and 41, 43, and 45 in pool 9 activated CD4+ T lymphocytes in DR1501 participant 1 (Fig. 1C). In addition, peptides 22 and 23 in pool 5; 37, 38, and 39 in pool 8; and 45 in pool 9 activated CD4+ T lymphocytes in DR1501 participant 2 (Fig. 1D). As stated earlier, the peptides were synthesized consecutively with 8-aa overlaps, therefore peptide 38 may contain an epitope. Accordingly, we suggest that peptides 22, 23, 37, 38, 39, and 45 are DR1501-restricted CD4+ T-cell epitopes (Fig. 4A).
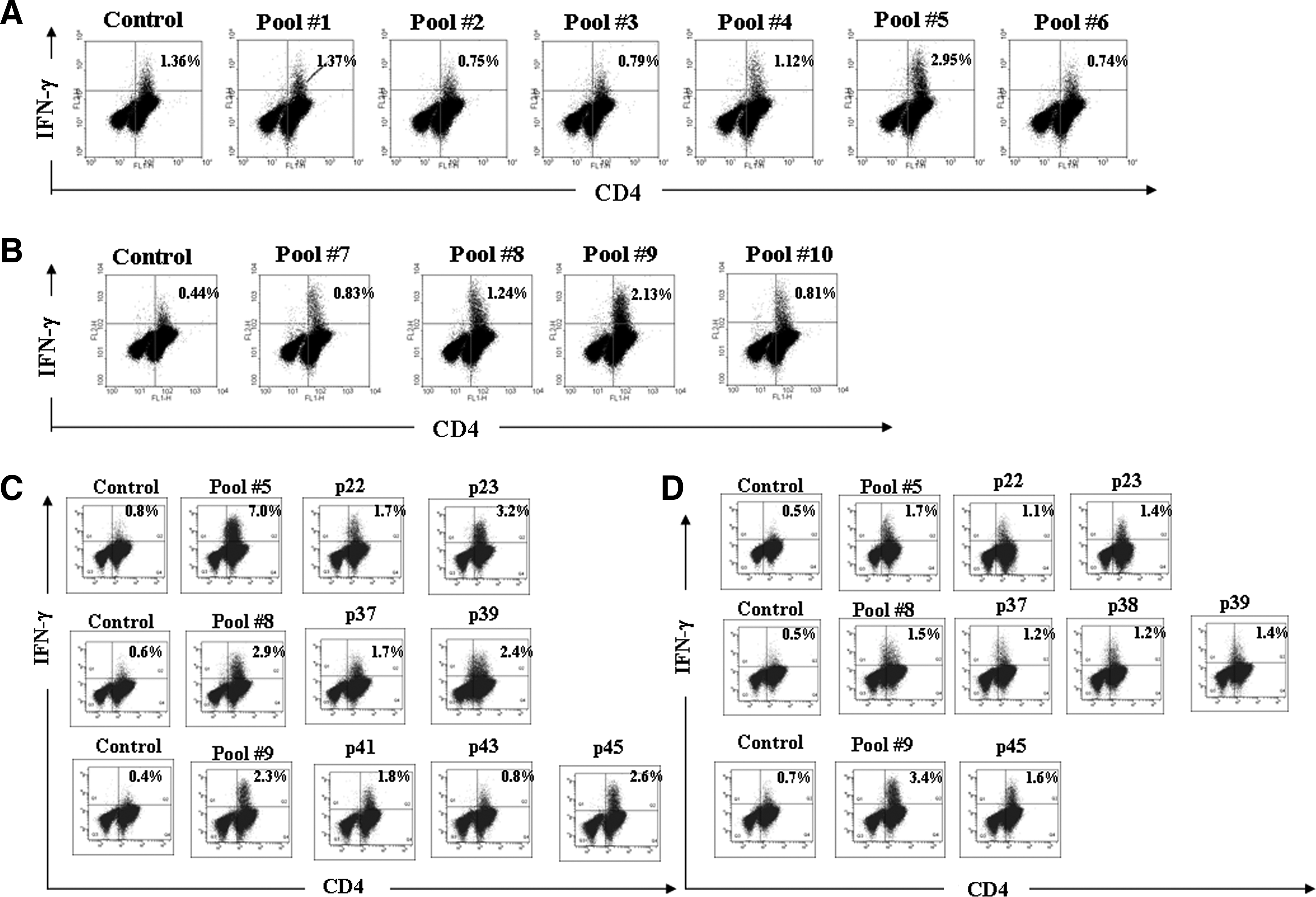
Identifying epitope pools and individual epitopes in the HA1 protein of the A/Vietnam/1203/04 (H5N1) virus by intracellular cytokine staining. Peripheral blood mononuclear cells (PBMCs) collected from participant 1 were stimulated with 10 peptide pools
We also performed tetramer staining to confirm that antigenic peptides containing the H5N1 HA protein were restricted by HLA DR1501. Cells isolated from the two DR1501 participants were stimulated with peptides 22, 23, 37, 38, 39, and 45, and stained with HLA DR1501 tetramers loaded with their corresponding peptides. As shown in Fig. 2, significant T-cell increases were observed in cultures containing peptides 23 and 39 in both DR1501 participants 1 (Fig. 2A) and 2 (Fig. 2B).
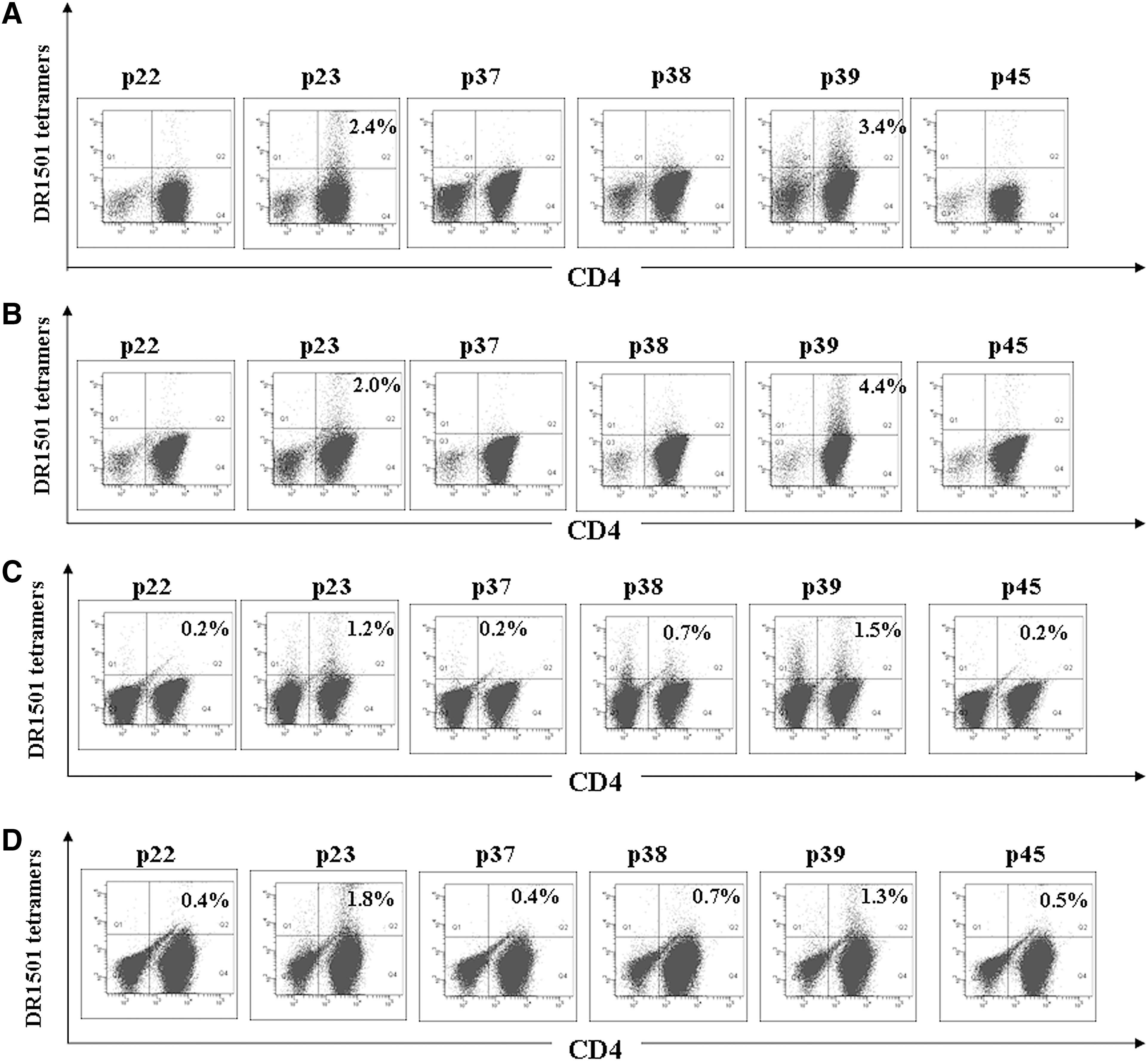
Results from tetramer staining of peptide-stimulated CD4+ T cells. Primary CD4+ T cells were stimulated in vitro with 10 μg/mL peptide in the presence of antigen-presenting cells. After expansion, cells from participants 1
To ensure peptide 23 and 39 derivation from native protein processing, primary CD4+ T cells collected from DR1501-carrying participants were stimulated with glycosylated HA (H5) proteins produced by a baculovirus cell-based system primed by autologous monocytes. Following in vitro expansion, the cells were analyzed by peptide-loaded tetramer staining. As shown in Fig. 2, we found significant increases in DR1501-restricted T-cell numbers in participants 1 (Fig. 2C) and 2 (Fig. 2D) for peptides 23 and 39, confirming that naturally-processed epitopes recognized by specific T cells contain these peptides.
We aligned and compared the amino acid sequences of H5HA155–169 (peptide 23) and H5HA267–281 (peptide 39) with the sequences of HA proteins from H1N1, H3N2, and influenza B viruses. As shown in Fig. 3, both epitopes were highly conserved in the clades of currently circulating H5N1 viruses. These epitopes are only found in H5HA, not in seasonal H1N1, H3N2, or influenza B viruses.

HA155–169
Discussion
The list of newly emerging or re-emerging diseases in the past two decades includes SARS, the Sapovirus, the Nipah virus, and influenza viruses such as H5N1 and H9N2 (16). Vaccines are considered useful prophylactics, with epitopes conserved in antigens considered essential for efficient post-vaccination antibody production. Most researchers have focused on mapping neutralizing epitopes that might be used for peptide vaccine or therapeutic agent development (24,31). In influenza viruses, neutralizing epitopes are involved in antigenic drift, which helps viruses escape attacks by neutralizing antibodies. Identifying T-cell epitopes against infectious microbes is a key step in clarifying how the human immune system reacts to pathogens. However, epitope selection is dependent on a combination of antigen processing, peptide binding to MHC complexes, and interactions between MHC/peptide complexes and corresponding T-cell receptors. MHC class I complexes present antigens to CD8+ T lymphocytes and activate them to kill infected cells (18). MHC class II complexes also play a key role in activating CD4+ T lymphocytes, inducing cytokine secretion, and mediating CD8+ T-lymphocyte or B-cell activation (3).
In this study, we recruited 30 participants who were inoculated with the GSK H5N1 vaccine. After determining the titers of the anti-H5N1 antibodies, we selected the DR1501 allele for identification of restricted epitopes on H5HA proteins. Our reasons for this decision were (1) a large proportion of Minnan Taiwanese carry this allele, (2) participants with DR1501 alleles had higher antibody titers, and (3) the wide availability of blood for analysis from donors with the HLA-DR1501 allele.
Our goal was to map the T-cell epitopes of H5N1 hemagglutinin proteins, using participants who were given two or three H5N1 vaccine booster injections. Results from hemagglutination inhibition (HI) assays indicated that post-vaccination, most of the participants produced neutralizing antibodies comparable to the quantities in their pre-immune sera (data not shown). Results from H5N1-RG-coated ELISA assays to determine antibody titers in H5N1 vaccinees indicated 80% effectiveness in H5N1 antibody production. In comparison, a study involving an inactivated influenza vaccine for military personnel resulted in 70–90% efficacy (11). Measurements of antibody titers secreted by activated B cells can be used to indirectly evaluate CD4+ T-lymphocyte activation status. Accordingly, we suggest that the GSK H5N1 vaccine exhibits greater efficacy in terms of activating both T and B lymphocytes.
In one study of dual-specific T-cell epitopes in H5N1 virus hemagglutinin, Haghighi et al. (15) used peptides that were 15 residues long, with 10 overlapping residues. In a separate study the most abundant individual molecular mass values were between 1700 and 1800, corresponding to an average peptide length of 15 residues bound to HLA-DR1 (8). In the present study the peptides used for CD4+ T-cell epitope mapping were 15 amino acids long, with 8 overlapping residues.
Roti et al. (26) studied the reactions of CD4+ T cells collected from healthy individuals to H5N1 influenza viruses, but focused on DR0101, DR0404, DR0701, and DR1101 HLA alleles, which are more prevalent in Caucasian populations. Since our focus was on Minnan populations, we focused on the DR1501 allele (20). Our ICS assay data indicate the potential presence of HLA DR1501-restricted T-cell epitopes of H5 HA1 proteins in six peptides: 22 (H5HA148–162), 23 (H5HA155–169), 37 (H5HA253–267), 38 (H5HA260–274), 39 (H5HA267–281), and 45 (H5HA309–323). We used peptide-loaded tetramer staining to confirm that H5HA155–169 and H5HA267–281 are indeed DR1501-restricted. One research team recently used an algorithm to predict the presence of T-cell epitopes according to specific haplotypes, and reported that peptides from the H5N1 virus proteome (predicted to bind with different HLAs) were dissimilar to other human proteome peptides (23). We tried using the same algorithm to predict HLA class II DR1501-restricted epitopes of H5 HA1 proteins, but results from the SYFPEITHI, RANKPEP, ProPred, and IEDB predictive programs were inconsistent. According to our ICS results, only a small number of T-cell epitopes can be accurately identified by a predictive program, suggesting that even though computer prediction tools are fast and convenient, conventional assays are still required to confirm their results.
We mapped the H5HA148–162 and H5HA155–169 CD4+ T-cell epitopes directly onto the structure of H5 hemagglutinin. Both were located toward the outer regions in comparison to the potential epitopes H5HA253–267, H5HA260–274, H5HA267–281, and H5HA309–323 (Fig. 4). However, T-cell-recognized epitopes bind with MHC class I or II complexes in the rough endoplasmic reticulum or endocytic compartments following antigen digestion. Peptides that are hydrophilic, hydrophobic, or both, may bind with MHC and stimulate T cell activation, suggesting that T cell epitopes may reside in either outer or inner regions.
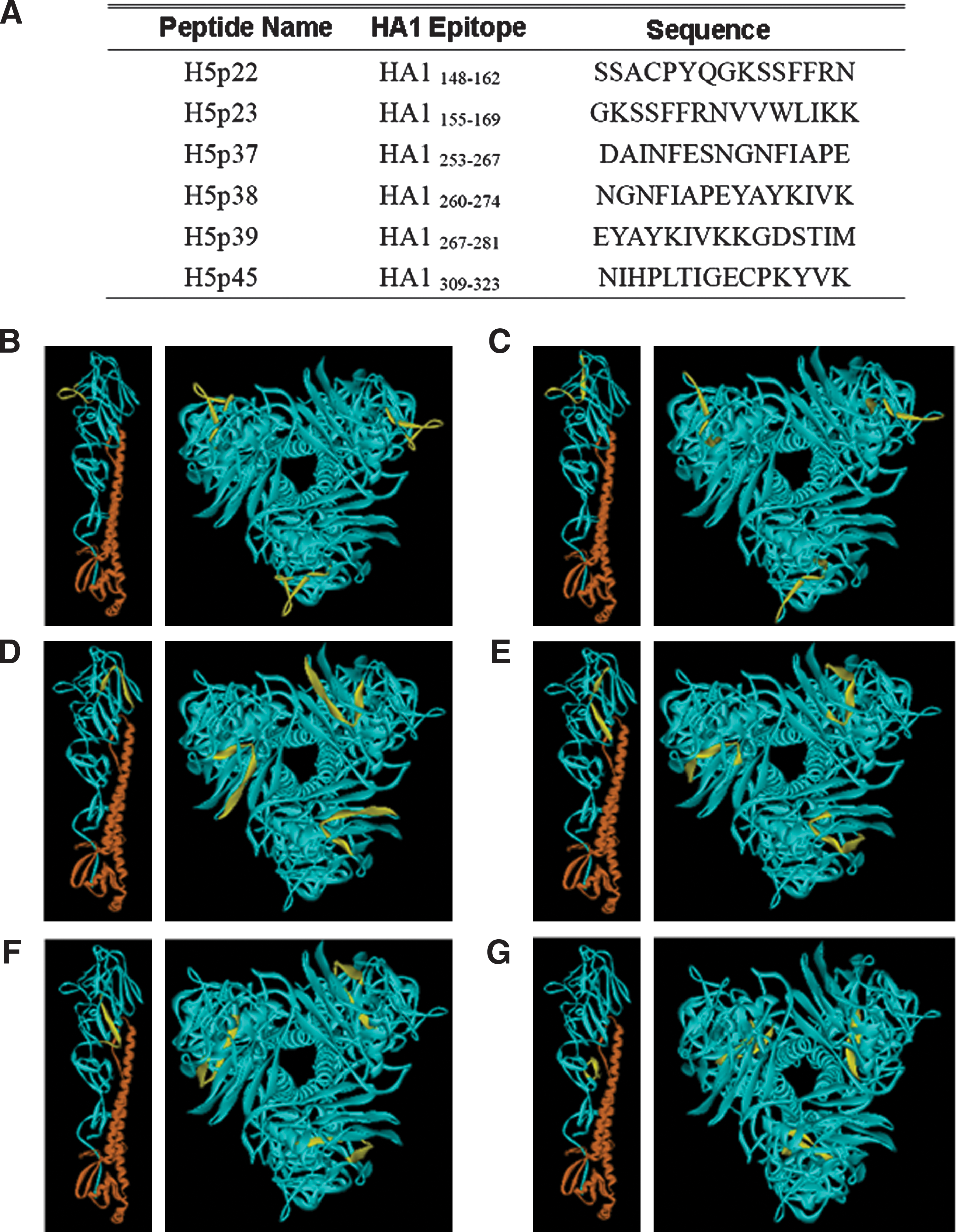
Sequences and direct mapping of potential DR1501-restricted CD4+ T-cell epitopes on the HA mono- and tri-structures of A/Vietnam/1203/04.
Regarding the T-cell epitopes of HA proteins in influenza viruses, HA1 proteins with non-synonymous change properties in specific antigenic regions are capable of mutating under host immune pressure, resulting in antigenic variation that can trigger human epidemics (31,32). As previously described, these T-cell epitopes are essential for initiating adaptive immune responses for clearing or neutralizing influenza viruses, although some researchers have indicated that the epitopes of nucleoprotein or matrix proteins are also important for T- or B-cell activation (2,33). However, these proteins are highly conserved in all influenza A viruses, and therefore cannot be used to identify specific influenza subtypes. One research team recently reported that NP and M protein epitopes in the A/Vietnam/1203/04 H5N1 virus are capable of activating CD4+ T cells collected from healthy individuals who have never been exposed to the H5N1 virus; they identified one DR1501-restricted H5 epitope mapped to the peptide GFLDVWTYNAELLVLMENER (aa 433–452) in the HA2 domain (26). In contrast, H5HA155–169 and H5HA267–281 are both in the HA1 domain and H5N1-specific, suggesting that H5HA155–169-reactive and H5HA267–281-reactive CD4+ T cells in the two DR1501 participants resulted from H5N1 vaccination, without any cross-reactivity with other influenza viruses.
In summary, the combination of intracellular cytokine staining and tetramer confirmation appears to be a valid alternative for epitope mapping. We used this approach to identify H5HA155–169 (peptide 23) and H5HA267–281 (peptide 39) as DR1501-restricted CD4+ T-cell epitopes in H5N1 vaccinees.
Footnotes
Acknowledgments
The authors wish to thank Prof. Eddie James of the Benaroya Research Institute at Virginia Mason Hospital in Seattle for providing DR1501 tetramers and for discussions regarding protocols, and Jon Lindemann for editing assistance. This work was supported by grants from the Republic of China National Science Council (NSC-98-2321-B-010-004 to Y.-M.A. Chen, and NSC-97-2320-B-010-011 to J.C. Huang).
Author Disclosure Statement
No competing financial interests exist.