
Abstract
The binding of murine IgM mAbs to five different clades of HIV-1 was examined using a modified ELISA-based virus capture assay. Two murine multispecific IgM mAbs that exhibit both lipid and gp41 epitope specificities, and one murine IgM mAb that exhibits lipid-binding specificity, were utilized. The binding of the IgG and the IgM isotypes of human mAb 2F5 to clades A through AE were also evaluated. The binding of 2F5 to HIV-1 was dependent upon the antibody isotype. Monoclonal IgM antibodies bound significantly lower amounts of HIV-1 than the corresponding IgG isotype. Although murine IgM mAbs bound HIV-1 to varying degrees in the virus capture assay, they failed to neutralize HIV-1 in a TZM-bl pseudovirus assay. In contrast, 2F5-IgM mAb bound certain HIV-1 isolates, and also neutralized them, although not as efficiently as the 2F5-IgG isotype. Studies on the relationship between virus binding and neutralization in a TZM-bl pseudovirus assay indicated that in most cases, mAbs that exhibited neutralization also bound the virus.
Introduction
Multispecific monoclonal murine IgM antibodies (WR316 and WR320) generated against clade B (HXB2) MPER peptide (amino acids 662–684) that simultaneously bind to HIV-1 gp41 and lipid epitopes have been produced in our laboratory and extensively characterized for their lipid and gp41 epitope specificity (2,9,12,13). The core-binding specificity for WR316 (SLWNWF) overlaps the published core specificity of 4E10, while the binding epitope for WR320 (LELDKWASL) maps to the 2F5 binding epitope (13).
The importance of lipids in the dynamic process of HIV-1 fusion and entry and/or in the budding process is strengthened by the demonstration that the murine anti-phosphatidylinositol-4-phosphate (PIP) monoclonal antibody WR304 neutralized the infection of two primary HIV-1 isolates (clades A and B) in a PBMC assay (3,4). Similarly, WR301, another murine monoclonal antibody raised against PIP, also neutralized a tier-1 virus in the PBMC assay (13). Both WR301 and WR304 exhibit only lipid binding, and neither binds to recombinant gp120 or gp41. However, the binding of these murine mAbs to the virus has not been examined.
In the current study, we investigated the ability of murine mAbs to bind to pseudoviruses from clades A, B, C, D, and CRF01_AE, using a modified virus capture assay. In addition, utilizing human mAb 2F5, we evaluated if the isotype of the mAb affected the binding to the pseudoviruses. The relationship between virus binding and neutralization in a TZM-bl pseudovirus assay was also examined.
Materials And Methods
Pseudoviruses and antibodies
293T cells (5 × 106) were transfected with 8 μg of env expression plasmid, and 24 μg of an env-deficient HIV-1 backbone vector (pSG3ΔEnv) using FuGene transfection reagent (Roche, Indianapolis, IN). Culture supernatants containing the pseudoviruses were harvested on day 3, stored at −80°C overnight, and then transferred to a liquid nitrogen freezer. The p24 concentration in each stock was determined using a commercially available p24 ELISA kit (ABL Inc., Kensington, MD). The HIV-1 isolates used in the present study were CCR5-tropic pseudoviruses belonging to five different clades (Table 1). 93RW is a dual-tropic virus.
TZM-bl pseudovirus assay was performed as described in the materials and methods section.
Murine monoclonal IgG antibody (mAb Hyb 278-01) specific to tetanus toxoid was purchased from Abcam Inc. (Cambridge, MA). Murine IgM mAb clone TEPC 183 specific to phosphorylcholine was purchased from Sigma-Aldrich (St. Louis, MO). Human IgM mAb IS5-20C4 specific to keyhole limpet hemocyanin (KLH) was purchased from Miltenyi Biotec (Auburn, CA). IgG and IgM variants of human mAb 2F5 were purchased from Polymun Scientific GmbH (Vienna, Austria). Murine IgM mAbs specific to PIP and multi-specific mAbs that simultaneously bind to lipids and also to the MPER region of gp41 were produced and characterized as previously described (13,21). Anti-murine and anti-human IgM antibodies were purchased from KPL (Gaithersburg, MD) and Thermo Scientific (Rockford, IL), respectively.
Virus capture assay for IgM monoclonal antibodies
The virus capture assay was modified from Nyambi et al. (15). Protein G-coated 96-well microtiter plates (Thermo-Fisher, Pittsburgh, PA) were washed, and 100 μL of a 10-μg/mL solution of mAb 2F5 (IgG) in sodium carbonate buffer (pH 9.6) was added to duplicate wells. To measure the binding of multi-specific IgM monoclonal antibodies to HIV-1, anti-murine or anti-human IgM antibodies (100 μL/well) were coated at a concentration of 10 μg/mL on protein G plates. The plates were incubated overnight at 4°C. The next day, the contents of the plates were dumped and tapped before the addition of 100 μL of murine IgM mAbs (WR301, WR316, and WR320), control antibodies TEPC 183 and IS5-20C4, or human IgM mAb 2F5, at a concentration of 10 μg/mL. The IgG and the IgM plates were sealed and incubated overnight at 4°C. The next day, the plates were washed once with PBS before the addition of 250 μL of blocking buffer (0.5% casein and 0.5% BSA in PBS) for 1 h at 37°C. The plates were washed once with RPMI-1640 media. Culture supernatant containing HIV-1 (100 ng of p24/mL for all HIV-1 clades tested) was diluted in RPMI-1640 media, and 100 μL was added to each well. The plates were incubated for 1 h at 37°C. The plates were washed five times with RPMI-1640 media before the addition of 200 μL of a 1:5 dilution of lysis buffer in RPMI-1640 media (p24 kit; ABL Inc.). Samples were assayed for p24 according to the manufacturer's instructions.
The binding of the antibodies to the virus was determined by the amount of viral p24 present in each case. Data are expressed as picograms per milliliter ± SD. Each experiment was repeated a minimum of three times and all data points were done in duplicate. In each experiment, virus only and mAb only were also included. The p24 values for the negative control Hyb278-01 (anti-tetanus toxoid antibody) for all viral isolates tested ranged from 0–7.7 pg/mL (background value). Therefore, a p24 value ≥15.4 pg/mL (twice the background value) was considered positive. IgM TEPC 183 mAb and IgM mAb IS5-20C4 were used as non-specific binding controls for lipid and protein binding, respectively. After averaging the p24 values for the control antibodies TEPC 183 and IS5-20C4 for each of the 10 viral isolates tested, it was found that the background binding ranged from 1.8–2.7 pg/mL. A p24 value ≥5.4 pg/mL (twice the background value) was considered positive for the IgM binding studies. In all cases, the p24 values obtained with virus alone were subtracted from p24 values obtained with virus and antibody before being analyzed.
TZM-bl assay for neutralization (IC50)
Virus (25 μL; 200 TCID50) was incubated with dilutions of (25 μL) test IgG (25 μg/mL), or IgM (250 μg/mL) mAbs in duplicate for 1 h at 37°C in a total volume of 50 μL of growth medium in 96-well flat-bottom culture plates (Corning-Costar, St. Louis, MO). A 50-μL aliquot containing 1 × 104 TZM-bl cells and 60 μg/mL DEAE-dextran was added to each well. Each plate included wells with cells and virus (virus control) or cells alone (background control). After 48 h, the cells were lysed and 100 μL of the reconstituted substrate was added to each well. The relative luminescence units (RLU) were measured using a Victor 2 luminometer (Perkin-Elmer Life Sciences, Shelton, CT). Wells producing RLU greater than 2.5 times background (signal of cells without virus) were scored as positive (10). The 50% inhibitory dose (ID50) was defined as the dilution that resulted in a 50% reduction in RLU compared to virus control wells after subtraction of background RLU (11). Titers were calculated as previously described (5).
Statistical analysis
The data were analyzed for significance using paired t-tests. A p value ≤0.05 was considered significant and is depicted by an asterisk in Fig. 1.
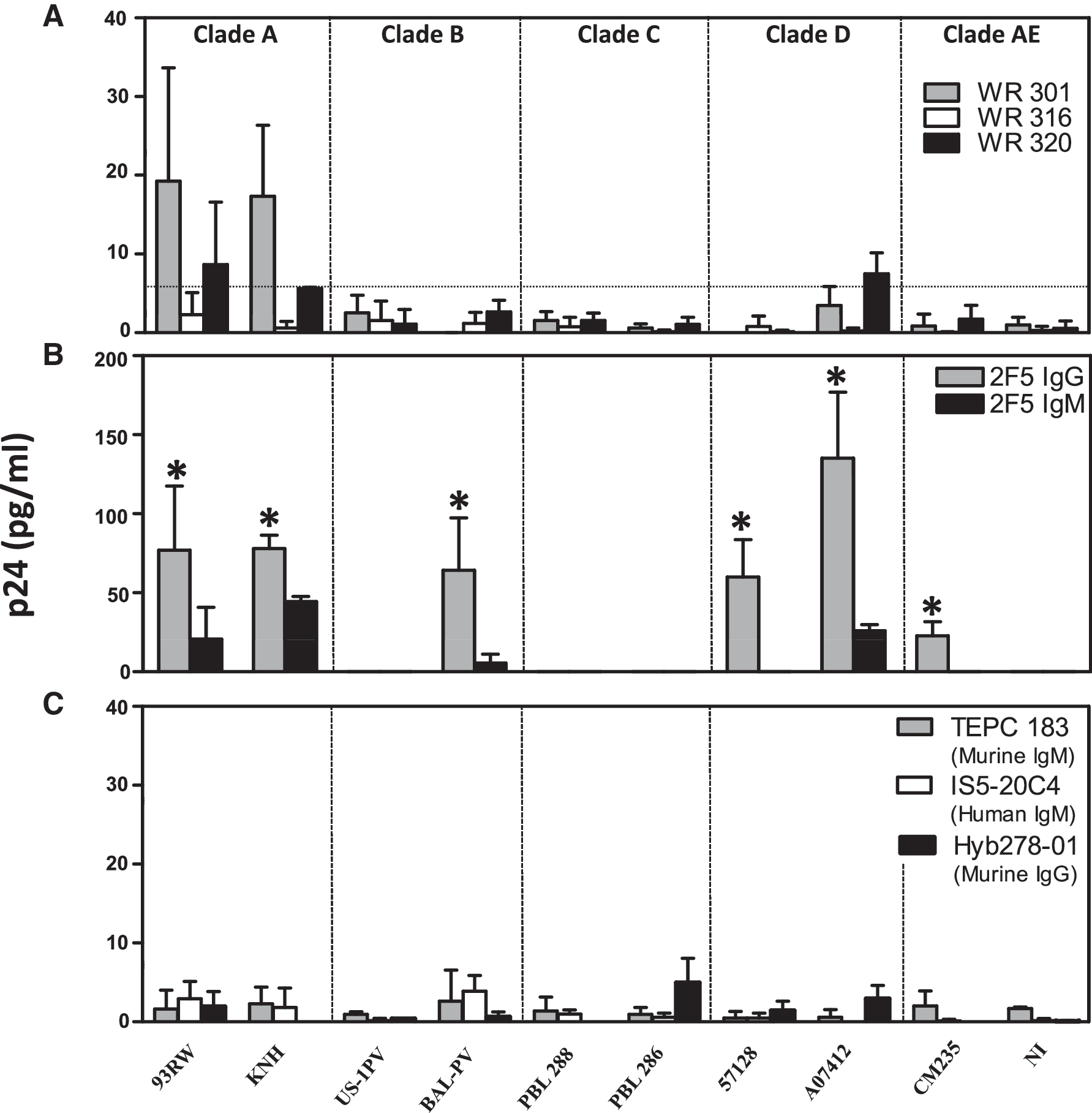
Binding patterns of murine and human monoclonal antibodies to HIV-1 clades. The binding of
Results
Murine IgM mAbs (WR301, WR316, and WR320), and human mAb 2F5 (the IgG and IgM isotypes) were examined for binding to clades A through AE in the virus capture assay. As shown in Fig. 1A, WR301 (grey bars) bound to 93RW and KNH (clade A viruses), while WR320 (black bars) bound low levels of 93RW. The binding was slightly above the cut-off value of 5.4 pg/mL, as depicted by the dotted line for KNH (panel A; 5.6 pg/mL). WR320 also bound low levels of clade D virus (A07412; black bars). In contrast, WR316 (white bars) did not bind to any of the clades tested. All three murine mAbs failed to bind to clades B, C, and AE.
The binding of the IgG and the IgM isotypes of 2F5 to clades A through AE is shown in Fig. 1B. The binding of 2F5 IgG (grey bars) to 93RW and KNH (clade A), BAL-PV (clade B), 57128 and A07412 (clade D), and CM 235 (clade E), was significantly higher in all cases (p ≤ 0.04), compared to 2F5 IgM mAb (black bars). US-1PV did not bind to either of the 2F5 isotypes. In clade C viruses (PBL 286 and PBL 288), since the 2F5 binding site was mutated, the mAb did not bind to either of the 2F5 isotypes.
Fig. 1C shows the binding of anti-TEPC 183 IgM mAb (grey bars) specific to phosphorylcholine (negative control for lipid binding), anti-IS5-20C4 IgM (white bars) specific to KLH (negative control for protein binding), and mAb Hyb278-01 specific to tetanus toxoid (black bars) to all 10 of the pseudoviruses. The p24 values obtained with both the negative controls were averaged (2.7 pg/mL). Therefore, a p24 value greater than or equal to twice the negative controls (5.4 pg/mL) for the test samples was considered positive. mAb Hyb278-01 (Fig. 1C) served as the negative control for 2F5 IgG. The data were analyzed after the p24 value obtained with virus alone (in the absence of antibody) was subtracted from all the test samples. Although only two viruses per clade were analyzed, it appears that both murine and human mAb IgM and IgG capture clade A and clade D viruses at higher levels than clades B, C, and AE.
The ability of murine IgM mAbs WR301, WR316, and WR320 to neutralize the virus in a TZM-bl pseudovirus assay was examined (Table 1). All of the IgM mAbs were tested at a concentration starting at 250 μg/mL, while 2F5 IgG was tested starting at 25 μg/mL for the neutralization experiment. Murine IgM mAbs (WR301, WR316, and WR320), and the human 2F5 IgM mAb, that bound pseudoviruses in the virus capture assay were the ones chosen for testing in the TZM-bl neutralization assay. Therefore neutralization of the following viruses was examined: clade A (93RW and KNH), clade B (BAL-PV), and clade D (A07412). Since the mAbs did not bind to clade C (PBL 286 and PBL 288) or clade CRF01_AE (CM235 and NI) viruses, they were not included in the neutralization assay. Murine IgM mAb (WR301) bound low levels (just under the cut-off range) of clade B (US-1PV) virus. Therefore, we also included this virus in the neutralization assay. WR301, WR316, and WR320 did not neutralize any of the viruses tested (93RW, KNH, US-1PV, BAL-PV, and A07412) in the pseudovirus assay (Table 1), although these mAbs bound to clades A and D viruses to varying degrees in the virus capture assay, as demonstrated above. 2F5 IgM mAb neutralized 93RW, KNH, and A07412 (IC50 1.25–156.6), but not US-1PV or BAL-PV. In contrast, 2F5 IgG mAb neutralized these viruses with an IC50 value of 0.06–6 μg/mL (Table 1).
Discussion
In the present study, a virus capture assay previously developed by Nyambi et al. (15,16,17) was modified to determine the binding of human and murine mAbs that recognize HIV-1 gp41 epitopes. The antibody was bound to protein G-coated ELISA plates through its Fc region, allowing the antibody binding sites to be accessible before capture of the virus (6). Using this assay, we analyzed the binding of murine IgM mAbs and the IgG and IgM variants of 2F5 to R5-tropic pseudoviruses from clades A, B, C, D, and AE. The reason for focusing on R5-tropic pseudoviruses was that it has been reported that newly transmitted viruses specifically use R5 co-receptors (19). To minimize envelope variability, pseudoviruses representing clades (A, B, C, D, CRF01_AE) that encompass the majority of clades currently circulating around the world were chosen.
The three murine mAbs analyzed showed varying degrees of binding to the pseudoviruses tested. WR301 bound to clade A viruses, while WR320 bound low levels of clade A and clade D viruses (A07412). WR316 did not bind to any of the viral isolates tested. Since WR301 exhibits only lipid and not protein specificity, the binding that was observed must have been due to phosphatidylinositol-4-phosphate (PIP) on the surface of the virions. In contrast, mAb 2F5 exhibits specificity not only to the MPER region of gp41, but also to various lipids (8,12,20). Although the 2F5 binding epitopes for the IgM and the IgG variants were identical, there was a dramatic difference in the binding of the IgM isotype. The IgM isotype of 2F5 bound clades A, B, D, and AE at a significantly lower level (two- to 150-fold) than the corresponding 2F5 IgG isotype. These results suggest that steric hindrance could be contributing to the decreased binding observed with the IgM antibody, as the IgM antibody is a pentamer and is considerably larger than the IgG molecule.
It is well known that retroviruses including HIV-1 acquire their lipids during budding from the plasma membrane of a host cell (14). A comprehensive mass spectrometry analysis of the lipid content of HIV-1 demonstrated that it was enriched in glycerophosphatidyl inositol monophosphate and glycerophosphatidyl inositol bisphosphate, cholesterol, ceramide, and ganglioside GM3 (7). Therefore, it is not surprising that WR301 bound to the virions in the virus capture assay, since it was induced against PIP. In the absence of any experimental data, we can only speculate on possible reasons for the differences seen in the binding of WR301 to the various virions examined in this study. To the best of our knowledge, a systematic study of the lipids and lipid compositions of the various HIV-1 clades has not been reported. Since the virions bud from lipid rafts on the cells, they could acquire different amounts of different host lipids on their surface, leading to heterogenous lipid compositions on the surface of virions among the different clades.
It was previously shown that WR301 neutralized a tier-2 virus in the PBMC assay (13). In the present study, although WR301 exhibited binding to the virus, it did not neutralize clades A, B, and D viruses in the pseudovirus assay. The reasons for this are not clear; one could speculate that WR301 must bind to the correct target on the virus to be able to neutralize it, or that there is steric hindrance by the pentameric IgM molecule. The steric hindrance argument is strengthened by the demonstration that 2F5 IgG neutralized the viruses tested with an IC50 value 16- to 29-fold lower than 2F5 IgM (Table 1). In addition, US-1PV and BAL-PV were neutralized by the IgG variant of 2F5, but not by the IgM variant, although 2F5 IgG did not bind to US-1PV in the virus capture assay. We hypothesize that the binding site on US-1PV for 2F5 mAb may be cryptic and not accessible in the virus capture assay. However, this site may be revealed only during the fusion step leading to neutralization of the virus by the mAb. Furthermore, a difference in the neutralizing ability of the two isotypes in the PBMC assay was previously reported (22), and recently confirmed (13). In any case, it appears that the IgG molecule neutralizes the virus much more efficiently than the IgM antibody.
Our data as well as data from others indicate that binding does not equate to neutralization using the assays currently available. The two predominant HIV-1 neutralization assays used are the PBMC and the TZM-bl assays. These two assays only show 60% concordance in qualitative neutralizing activity, thus further complicating the interpretation of correlation between binding and neutralization (18). The PBMC assay incorporates multiple rounds of infection and captures all stages of the virus entry, replication, and budding. However, it introduces additional complexities, including host genetics and donor cell population variability. Unlike the PBMC assay, the TZM-bl assay is a single-round infection in which approximately 85% of the virus enters the cells by endocytosis. The two assays also differ in the amount of CCR5 receptors present on the cell surface (18). TZM-bl cells contain 100-fold more CCR5 receptors on their surface than primary PBMCs (reviewed in 18). This could potentially interfere in the neutralization by IgM antibodies through steric hindrance.
The absence of a correlation between binding and neutralization could be due to a number of factors, including the presence of non-functional envelope proteins on the surface of the virion. Binding of the mAbs can result in the shedding of gp120 and exposure of gp41. Experiments with additional mAbs that either bound the V3 loop of gp120, or the mannose residues on gp120, showed binding to the different HIV-1 clades in the virus capture assay (data not shown). With some of these viruses, a significant increase in virus binding was observed in the presence of sCD4, as determined by Biacore analysis (23), thus indicating that gp120 was present on the surface of the pseudoviruses examined in this study.
We are currently in the process of converting some of the murine multi-specific IgM antibodies to the IgG isotype. This will address the issue of steric hindrance of the IgM pentamer. We predict that the IgG antibodies will be better at neutralization.
Footnotes
Acknowledgments
This work was supported by a cooperative agreement (W81XWH-07-2-0067) between the Henry M. Jackson Foundation for the Advancement of Military Medicine, Inc., and the U.S. Department of Defense (DOD).
Disclaimer
The views and opinions expressed in this article are those of the authors and do not reflect the official policy of the Department of the Army, the DOD, or the U.S. government.
Author Disclosure Statement
No competing financial interests exist.