
Abstract
Chicken anemia virus (CAV) is an economically important virus affecting the chicken meat and egg industry. CAV is characterized by anemia, lymphoid depletion, and immunosuppression. Microarrays were used to investigate the response of MDCC-MSB1 cells (MSB1) to infection with CAV at 24 and 48 h post-infection (hpi). The major genes responding to CAV infection include genes involved in inflammation, apoptosis, and antiviral activity. Several cytokines were differentially regulated at either 24 or 48 hpi, including interleukin 2 (IL-2), interleukin receptors IL-1R, IL-22R, IL-18R, and IL-7R, and interferon-α (IFN-α). While there were many genes differentially regulated in this experiment, only two genes were common to both time points, suggesting a dramatic change in gene expression over the two time points studied. The present study is the first microarray experiment to investigate CAV, and we identified a number of key pathways involved in viral infection. Overall, there were more genes upregulated at 24 hpi than at 48 hpi, including genes involved in cytokine signaling, apoptosis, and antiviral activity. The two time points were vastly different in their gene expression patterns, in that at 24 hpi there were many genes involved in the response to infection, whereas at 48 hpi there were many genes associated with apoptosis and immunosuppression.
Introduction
Chicken anemia virus (CAV), a member of the Circoviridae family, Gyrovis genus (1), is a non-enveloped, icosahedral virus about 25 nm in diameter with a negative sense single-stranded circular DNA genome. The CAV genome consists of 2.3 kb with three open reading frames (ORFs) encoding for VP1, the major structural protein, VP2, a scaffolding protein, and VP3, a non-structural protein called apoptin, due to its ability to induce apoptosis (2).
CAV has been recognized since the late 1970s as a pathogen that mainly causes clinical disease in chickens prior to 3 wk of age. The virus is associated with signs such as anemia, lymphoid depletion, hemorrhages, and immunosuppression in young birds (3). Subclinical infection in older chickens does not generally cause anemia; however, it still results in significant economic losses. One of the more important aspects of subclinical CAV infection is its ability to suppress the immune system, resulting in more serious problems when associated with other viruses (4 –6). CAV infection has also been associated with secondary bacterial infections and vaccine failures (7). CAV infections also cause problems in specific-pathogen-free (SPF) flocks used for vaccine production and research purposes. It is imperative that vaccine manufacturers maintain CAV-free SPF flocks. Unfortunately, seroconversion of CAV in SPF flocks often happens at or after sexual maturity, resulting in costly losses to the vaccine manufacturers and the SPF industry. Reasons for this late seroconversion are unknown.
To gain a better understanding of the host responses to CAV, whole genome microarrays were used to monitor gene expression in CAV-infected chicken cells. Investigation of the differential expression of the host genes during virus infection enable a greater understanding of the molecular basis of CAV pathogenesis, and can be used to gain an insight into mechanisms of host-virus interactions. In the present study, we report on gene expression in MSB1 cells infected with CAV at two time points during infection. The results provide a global view of the changes in response to CAV infection, from which we were able to identify groups of differentially expressed genes that point to possible mechanisms that may account for the immunosuppression associated with this virus.
Materials and Methods
MDCC-MSB1 cells (MSB1), a Marek's disease virus-transformed chicken lymphoblastoid cell line, was used in all experiments in this study (8). MSB1 cells are the only available in-vitro model to study CAV infection. MSB1 cells were grown at 37°C in a 5% CO2 atmosphere in RPMI-1640 medium containing 10% fetal calf serum. The cells were subcultured at 2- to 3-day intervals and seeded at 1 × 106 cells/mL/cm2. An Australian strain of CAV (CAU269/7), supplied by Dr. Glenn Browning at the University of Melbourne, was used for this study. CAV was propagated in MDCC-MSB1 cells as described previously (9). Twelve separate MDCC-MSB1 cell cultures in flasks, each containing one million cells, were used in this experiment. Six were infected with CAV at multiplicity of infection of 2, and the remaining six cultures were used as uninfected controls. CAV infection was checked using PCR to amplify a region of the CAV VP2 gene. All cell cultures were seeded at the same time. Cells were harvested at 24 and 48 hours post-infection (hpi).
All infected cultures were checked prior to RNA extraction to ensure the cells were infected with CAV. Total RNA for all 12 samples was isolated using the Meridian total RNA isolation kit (Cartagen Molecular, Inc., Seattle, WA). Quantity and quality of all RNA samples were checked using a nanodrop spectrometer. Then 5 μg of total RNA was reverse transcribed into cDNA and indirectly labeled with Cy3 using the ULS cDNA Synthesis and Labeling Kit (Kreatech Technologies, Amsterdam, the Netherlands). The labeled probes were concentrated using Microcon Ultracel YM-30 columns (Amicon Bioseparations, Bedford, MA), and the quality and label incorporation of each sample was verified using the NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE). Each Cy3-labeled cell culture sample was individually hybridized to the whole genome chicken array (single-color hybridization). This whole genome chicken microarray was printed with a MicroGrid II spotting robot in house, using a set of 20,460 long oligos (65–75 nt) printed in duplicate on each array. This array was designed at the Roslin Institute (Roslin Midlothian, Scotland, U.K.), based on chicken Ensembl gene transcripts and other genomic information supplied by various research groups around the world (
Whole genome chicken arrays were pre-hybridized at 42°C for 45 min in pre-hybridization buffer (25% [v/v] formamide, 5 × SSC, 0.1% [w/v] SDS, and 10 mg/mL salmon testes DNA). Following addition of the labeled probe and hybridization solution (25% [v/v] formamide, 5 × SSC, 0.1% [w/v] SDS, and 25% [v/v] KREAblock), all arrays were incubated for 16 h at 42°C. Post-hybridization, all arrays were washed once (2 × SSC, 0.1% [w/v] SDS) for 5 min at 42°C, once (0.1 × SSC and 0.1% SDS) for 10 min at 25°C, and three times in 0.1% SSC, each for 1 min at 25°C. Each microarray was subjected to a quality control check to ensure successful hybridization, that included calculating the overall intensity correlation, and flagging and removing spots where the replicates were not consistent or below two standard deviations of the background.
Each array was globally normalized, in which the output from each array is multiplied by a normalization factor such that the total signal intensities of all arrays are equivalent. Post-normalization arrays were separated into their corresponding groups (infected and control) at each time point, and the groups were compared directly to each other. Subsequent statistical tests were carried out using Genespring version 7.2 (Silicon Genetics, Redwood City, CA), to determine all genes differentially regulated by a factor of twofold or greater (p = 0.05) between the uninfected and infected groups at each time point. Quality threshold (Qt) clustering was performed to group genes that responded to infection in a similar way across the two time points, using standard correlation set at a minimum of 0.9 and a minimum group size of 10. Gene ontologies were assigned using GO annotation and mapped to their top-level entry. Pathway analysis was conducted using Genowiz Version 4.0.5.3.
Real-time PCR reactions were performed in duplicate using SYBR-green master mix in a final volume of 25 μL. All primers were used at a concentration of 50 nM per reaction. Then 5 μg single-stranded cDNA was synthesized using the Invitrogen Superscript III First Strand kit (Invitrogen, Carlsbad, CA). Gene-specific primers were designed for six randomly chosen differentially-expressed genes. Reactions were initially denatured at 95°C for 5 min, followed by 40 cycles of 95°C for 15 sec, and 60°C for 1 min. β-Actin was chosen as the internal control and was used to correct the differences in cDNA concentration in different samples. The expression results for β-actin were checked on all arrays to ensure its use as a suitable control. The number of cycles (ct) was used to calculate the relative expression of each gene in both the viral infected and uninfected samples. Amplification of specific target sequences were verified by examining the corresponding melting curves.
Results
Six independent cell cultures (three CAV-infected and three controls) per time point were used in single-color microarray experiments to detect genes that consistently responded to infection with CAV. At 24 hpi 70% of cells displayed cytopathic effects (CPE), and at 48 hpi 95% of cells showed signs of CPE. Control cell cultures at both time points displayed no visible signs of infection.
All microarrays were successfully hybridized and passed our quality control measures. Over 80% of probes on each individual array were expressed above background prior to normalization and analysis. A total of 880 genes at 24 hpi and 732 genes at 48 hpi were differentially expressed (p = 0.05) due to infection. Of the total genes differentially expressed, 463 and 449 genes were found to be differentially regulated at 24 hpi and 48 hpi, respectively, by applying a twofold cut-off and a false-discovery rate (FDR) of 0.05 (10). Subsets of these genes are represented in Table 1. Twenty-six genes were similar between the two time points at an FDR of 0.05; however, only two genes were found to be common between the two time points at a twofold cut-off (Fig. 1).
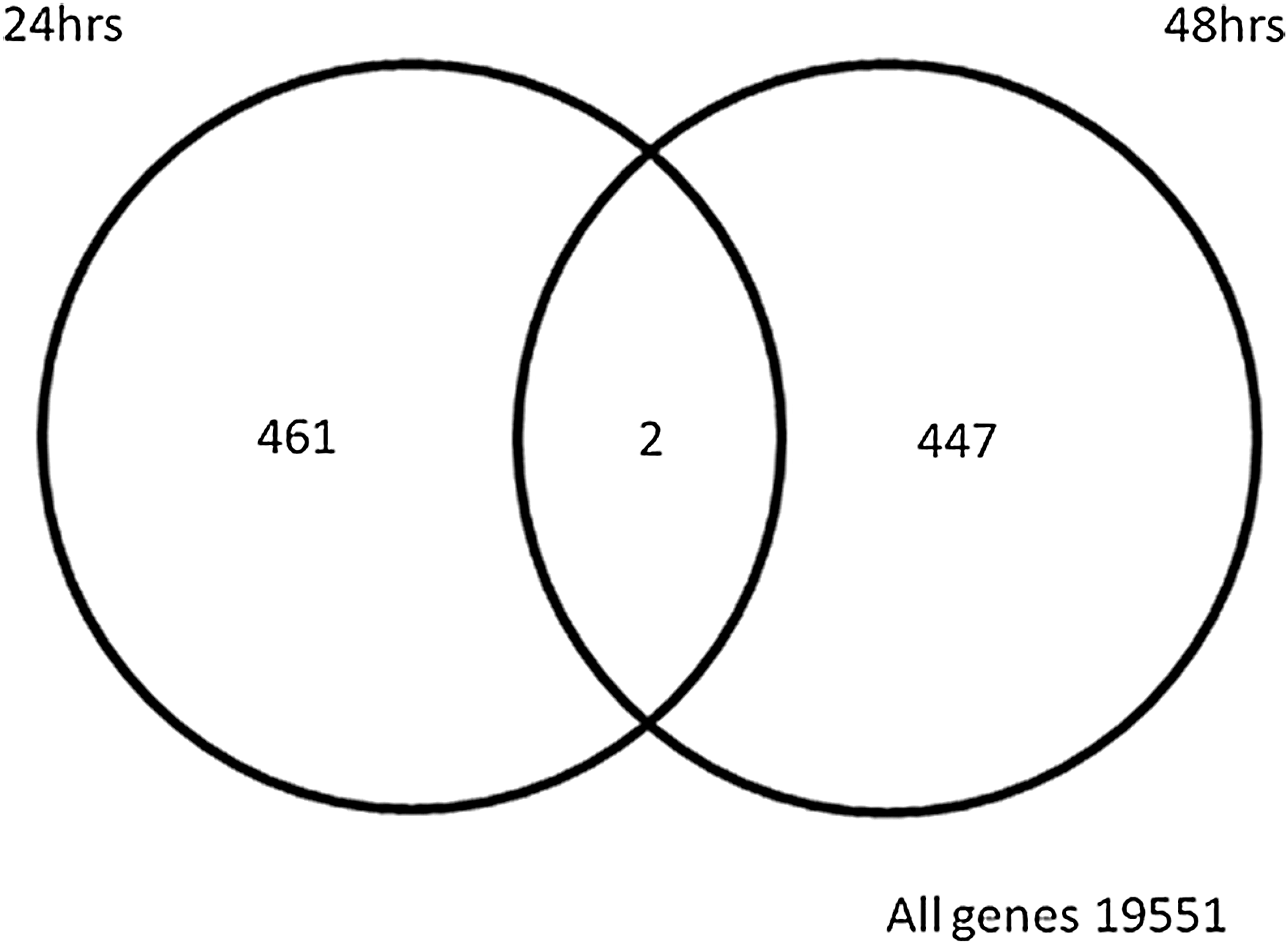
Venn diagram displaying the total number of differentially expressed genes in response to in vitro CAV infection at 24 and 48 hpi. The intersection of the two circles highlights genes that were regulated at both time points studied (this Venn diagram was generated using the software GeneSpring; CAV, chicken anemia virus; hpi, hours post-infection).
CAV, chicken anemia virus.
For each time point genes regulated above twofold were subject to pathway analysis to determine what pathways may be important during CAV infection. A total of 57 pathways were detected at 24 hpi, whereas there were 62 pathways detected at 48 hpi. Of the total pathways detected, pathways with only one gene present in both or one of the time points were excluded from further analysis. This resulted in 20 pathways containing multiple genes at 24 hpi, and at 48 hpi there were 17 pathways with multiple genes. Each time point had a distinct pathway signature, as shown in Fig. 2.
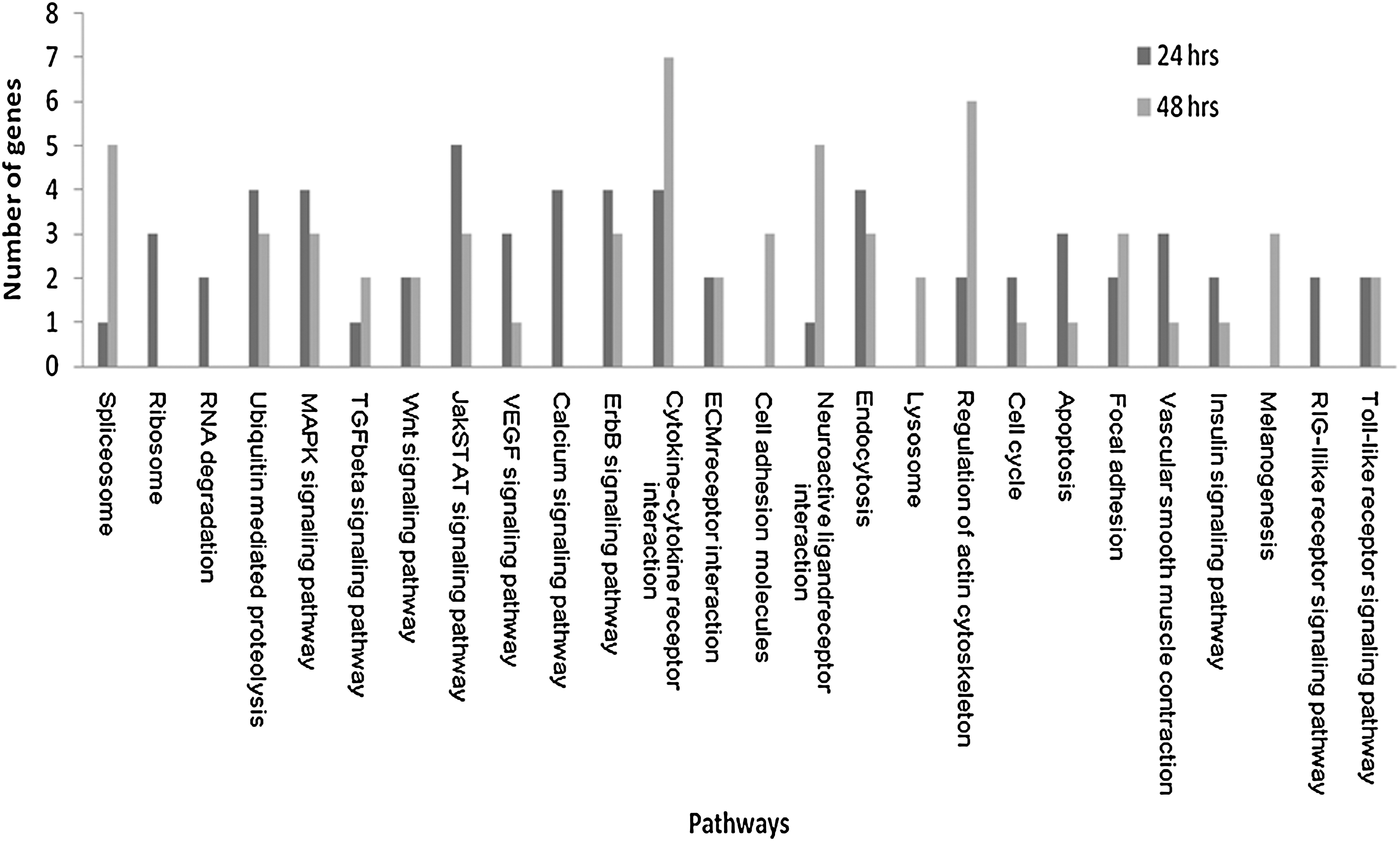
Pathways associated with differentially-regulated genes of CAV infection at 24 and 48 hpi (only pathways with multiple genes at either time point are displayed; CAV, chicken anemia virus; hpi, hours post-infection).
Prior to examining the roles of individual genes and pathways associated with CAV infection, we utilized the gene ontology (GO) terms associated with these gene products, in order to gain some insight into the types of processes that may be modulated by CAV infection (Supplementary Tables 1 and 2; see online supplementary material at
Clustal analysis of the regulated genes detected 30 distinct clusters; however, 40% of the regulated genes were unassigned (minimum group size >10; Supplementary Tables 1 and 2; see online supplementary material at
Several of the genes found to be regulated in this experiment were further analyzed by q-PCR, and while the absolute fold change in mRNA abundance was not identical to the microarray, directional changes were consistent in 5 out of 6 genes tested (Table 2). These results are equivalent to those of a number of other microarray experiments, in which researchers reported 70–90% confirmation of directional fold changes (11 –16).
RT-PCR, real time polymerase chain reaction.
Discussion
Microarray technologies provide a global approach to the characterization of gene expression patterns. This technique offers an expeditious and systematic means to generate the data required to elucidate genes whose products may be responsible for a host's response to viral infection. The use of the whole genome chicken array has provided a large data set from which we have been able to derive potentially valuable observations. Gene expression data obtained from infected and uninfected MSB1 cells were highly reproducible, yielding strong statistical support for differences in the expression levels of many genes. These results are supported independently by RT-PCR data for a range of differentially expressed genes from both time points studied. To date, there have been few studies that have explored gene expression in response to CAV in-vitro infection (17,18). However, these studies only looked at a few genes at a time, and did not explore this viral infection on a global scale. Our ultimate aim was to explore previous suggestions of what genes and pathways are involved in a host's response to CAV infection, and to confirm their expression using a whole genome chicken microarray (2,3). While we have been able to confirm many of the genes previously thought to be involved, we have also elucidated a number of new mechanisms that may play an integral role in this viral infection.
Interestingly only two genes were found to be common across both time points, suggesting that the in-vitro gene expression response to CAV infection changes markedly over the two time points studied. These rapid changes are complemented by the large number of clusters determined by cluster analysis, which revealed 30 distinct clusters based on their similar response to infection, suggesting a varied response to infection. In addition to the 30 clusters, over 40% of the genes were unassigned, signifying a high level of variation in the in-vitro response between the two time points studied (Supplementary Tables 1 and 2; see online supplementary material at
The major differences between virally-infected and uninfected cells were centered on genes involved in inflammation, apoptosis, and antiviral activity. A number of cytokine pathways were detected at both time points post-infection. Interestingly, at 24 hpi a number of genes involved in cytokine pathways are upregulated in response to infection, whereas at 48 hpi they are all downregulated. At 24 hpi IL-1R, IL-22Rα, and IFN-α are upregulated. These two cytokine receptors are implicated in the host inflammatory immune response (19 –21), and IFN-α is known to possess antiviral properties (21,22). IL-1 has been previously linked to CAV infection, and reductions have been seen in IL-1 production in infected spleen macrophages, and this decrease has been linked to interference with immune competence at all levels (23). However, here we found an increase in the production of the IL-1 receptor, suggesting an inflammatory response, a known characteristic of the IL-1 family (21). At 48 hpi IL-18Rα is upregulated, whereas IL-2 and IL-7R are downregulated. IL-18 is a member of the IL-1 family, thus this upregulation is likely to be involved in an inflammatory response. Both IL-2 and IL-7 have roles in the proliferation and development of T cells, and consequently downregulation of these genes would potentially affect the overall growth of MSB1 cells. In addition, IL-2 is necessary for the development of T-cell immunologic memory, one of the unique characteristics of the immune system (24). By downregulating IL-2, it is possible that CAV is circumventing this vital immune function, which may result in immunosuppression of the host cells. Not surprisingly, similar antiviral cytokine responses have been shown in the chicken's response to MDV, including the induction of IL-1β, IFN-α, and IL-18 (25-26).
Chemokines are part of the cytokine family and are defined by their chemotactic activity toward different cells of the immune system (27). Chemokines are also known for their role in angiogenesis, and at 24 hpi the angiogenic factor K60 is upregulated. This upregulation of K60 may be related to the upregulation of the IL-1 receptor, as K60 has previously been shown to be strongly induced by the proinflammatory cytokine IL-1β (20,28,29). K60 is a member of the CXCL class of chemokines, which are known for their role in inflammation. At 48 hpi there are three chemokine receptors that are downregulated, including CXCL receptor 5, CCXC receptor 1, and a CC chemokine receptor cluster. While this is in contradiction to the promotion of inflammation mentioned earlier via the upregulation of IL-18Rα, the reduction of chemokine activity is stronger, and may be related to a reduction in the inflammatory response caused by CAV at the later time point of virus infection.
As mentioned earlier only two genes were found to be regulated at both time points studied. The first gene was MHC class IV antigen. MHC class IV is one of three types of the MHC in chickens, and has been implicated in genetic resistance to MDV, together with MHC Rfp-Y (30,31). Both the MHC class IV antigen and the MHC Rfp-Y class I α chain are upregulated at 24 hpi, and they may play a role in the cell response to infection. The second gene is CASK interacting protein, and this gene is initially downregulated, but is twofold upregulated, at the later time point post-infection. Upregulation of CASK has been previously associated with a reduced rate of cell growth (32), and may be contributory to a decrease in cell growth at 48 hpi.
Apoptosis has been previously implicated in CAV infection, and it is believed that apoptin is one of the elements responsible for inducing the apoptotic pathway (2,23,33 –35). Findings of a previous study investigating the underlying mechanisms of CAV-induced apoptosis suggested that apoptin-induced apoptosis is independent of p53 and upstream caspases, and that author reported that caspase-3 and Bcl-2 are involved, but not essential (2). In our study, p53 was not differentially expressed at either time point; however, Bcl-2-like protein and caspase 9 were upregulated at 24 hpi. Caspase-9 has been previously shown to be involved in the production of caspase-3 (36); thus it is possible that we have captured an early snapshot of the apoptotic pathway. Interestingly, two other genes that have been previously associated with apoptosis were upregulated at 24 hpi, namely TIA-1 (37) and CIAPIN1 (38,39). TIA-1 is an RNA-binding domain-containing protein that is suggested to promote apoptosis (37), whereas CIAPIN1 is thought to be a cytokine-induced anti-apoptosis molecule (38). It would be useful to investigate these two genes in relation to apoptin and CAV-induced apoptosis. The results of this investigation have corroborated the occurrence of apoptosis induced by CAV infection.
Some of the other genes detected in this study that are likely to have been influenced by CAV infection include ChT1 thymocyte antigen precursor, T-cell receptor β, genes involved in transcription, cathepsin L, and heat-shock proteins. CAV is known to cause cell death and thymic depletion. Downregulation of ChT1 and T-cell receptor β may be linked to the depletion of thymocytes in the thymus post-CAV infection (34,35). Genes involved in transcription were upregulated at 24 hpi (BHLH-PAS, NR5A2, RUNX2/CBFA1, HSTF1, and TCF-9), possibly an initial response to the viral infection, whereas transcription genes were downregulated at 48 hpi (PBXIA, B2, and POU2F3), and this may reflect the ability of CAV to induce immunosuppression. Upregulation of cathepsin L in response to viral infection has been reported previously in chickens infected with MDV, and this may be a common response to viral infection (12). At both time points there were heat-shock protein genes (HSP2, HSP25, and HSP47) that displayed reduced expression, and these genes are often stress-induced (40), so it is intriguing to ask why they are downregulated. It is possible that these genes are involved in modulation of the inflammatory response (12), therefore their reduction would allow the virus to induce an inflammatory response.
Six microRNAs were shown to be downregulated at 48 hpi, including gga-mir-137, which until now was only a predicted microRNA in chicken. While microRNAs are thought to play a large role in regulating gene expression (41), it is difficult to say how these six microRNAs are involved in CAV infection, due to the lack of functional information available for these small molecules. However, it is interesting that viral infection downregulated all of the microRNAs detected, potentially reducing their control over gene expression in MSB1 cells. Perhaps the downregulation of these microRNAs is linked to CAV's ability to induce immunosuppression.
Overall pathway analysis of the genes differentially expressed at both time points revealed a number of immune-related pathways. These pathways included a number of important signaling pathways involved in the host response to viral infection. This analysis also detected the apoptosis pathway, which plays a pivotal role in CAV infection. While the pathways detected here complement our investigations into individual regulated genes, it must be noted that with the current state of the chicken genome annotation it is difficult to resolve all pathways involved in CAV in-vitro infection (42).
It must also be noted that as the cell line in this study harbors MDV, it is possible that the gene expression results here may be a reflection of the interaction between MDV and CAV. However, this is the only cell line currently available to study CAV infection in vitro.
Many of the genes represented on this whole genome array are of partially or completely unknown function, including many of those that display differential expression. Several of these genes of unknown function are some of the most highly expressed at both time points post-infection (Supplementary Tables 1 and 2; see online supplementary material at
In conclusion, the findings outlined here elucidate some of the mechanisms of CAV infection in vitro. These mechanisms include inflammation, apoptosis, antiviral activity, immunosuppression, and the possibility that CAV has developed strategies to circumvent the host cell immune response. One of the most interesting observations made here is that at 24 hpi many of the immune genes are upregulated, suggesting the cell is responding well to the virus. However, by 48 hpi many of the immune pathways have been diminished, possibly a reflection of CAV's ability to induce immunosuppression. Future microarray investigations should investigate CAV infection in vivo in order to delineate if the response described here is indicative of how infection unfolds in chickens. Obviously, the time points post-infection when samples are taken will be critical to the interpretation of the results of these experiments, as the gene expression changes that occur during infection are extremely dynamic. In addition, forthcoming studies should focus on defining the functions of the genes identified in this examination of in-vitro viral infection.
Footnotes
Acknowledgments
The authors would like to thank the Australian Poultry Cooperative Research Centres for their support and funding. We would also like to acknowledge Dr. Andrew Bean for his valuable knowledge of immunology, Ms. Barbara Konsak for her assistance with the pathway analysis, and Dr. Tracey Hinton for her support with the in-vitro CAV infection. Thanks also to Dr. Michelle Wilkins and Dr. Pengju Guo for their comments on this manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.