
Abstract
Persistent viruses, such as cytomegalovirus or human immunodeficiency virus, cause major perturbations of CD8+ T-lymphocyte subpopulations. To test whether chronic infection with hepatitis B virus (HBV) could also be responsible for such modifications, we analyzed the expression of CD27, CD28, CCR7, and perforin in blood CD8+ T lymphocytes. In comparison to healthy subjects and patients recovering from acute hepatitis B, chronic hepatitis B patients showed higher percentages of naïve CD8+ T lymphocytes (CD45RA+CD27+CD28+), and lower percentages of intermediately-differentiated CD27+CD28−CD8+ T cells. The late differentiated CD45RA+CD27−CD28− subset was also present in a large percentage in some patients, but no statistically significant difference with healthy controls was observed. Removal from the circulation of intermediately-differentiated CD8+ T lymphocytes may occur during chronic HBV infection, favoring the recruitment of naïve cells. This may result in impairment of the generation of functionally-competent memory cells, and an inability to achieve control of HBV replication.
Introduction
In the model of CD8+ T-cell differentiation proposed by Appay and Rowland-Jones (2), an example of a linear model in which the generation of memory cells derives from and follows the engagement of naïve cells into an effector phase, the progression along the functional phases of lymphocyte maturation is characterized by the sequential loss of the co-stimulatory molecules CD28 and somewhat later, CD27. Among the many markers whose expression varies during lymphocyte differentiation, CC chemokine receptor 7 (CCR7) and perforin represent key features: CCR7 essentially defines cells that are able to migrate to lymph nodes (naïve and central memory cells), and perforin, one of the mediators of lymphocyte cytotoxic activity contained in the cytoplasmic granules of effector cells, is characteristically expressed after the early stage of CD8+ T-cell differentiation. The RA isoform of CD45, whose expression is classically associated with the naïve cell phenotype (3), can be re-expressed at all stages of CD8+ T-cell differentiation, reflecting more a state of quiescence than a lack of previous encounter with the antigen (8,11). Phenotypic studies have shown that, during persisting infections, virus-specific CD8+ T cells accumulate at various stages of differentiation correlated to the infecting virus (1). The persistence of a virus, either latent or actively replicating, can therefore be reflected in a significant skewing of the total lymphocyte phenotype towards a particular maturation stage, at which the T cells specific for a given virus appear to accumulate. Even though such an enrichment may be masked by the wide variability of the general T-cell repertoire, infections with human cytomegalovirus (CMV) and human immunodeficiency virus (HIV) are associated with high percentages of the late differentiated CD8+ T cells corresponding to the phenotype of the respective specific cytotoxic T lymphocytes (CTL) (5,10,14,15). CMV seropositivity is associated with the accumulation of CD8+CD28− T cells, a constant feature of the proposed immune risk profile (IRP) of the elderly, which is significantly associated with high short-term mortality rates in this age group (20).
The hypothesis that stems from the above observations is that virus persistence hampers the physiological reversal of acutely expanded T-lymphocyte subsets to small-size memory cell populations, causing the abnormal overrepresentation of given cell phenotypes. We wanted to test this hypothesis by analyzing peripheral blood lymphocytes from chronic hepatitis B patients, another example of a persistent virus infection, with respect to their maturation phenotype in comparison to lymphocytes from healthy controls and subjects recovering from acute hepatitis B.
Materials and Methods
Subjects
Heparinized venous blood samples were obtained from 24 chronic hepatitis B patients, aged 18–56 y, before starting treatment with polyethylene glycol (PEG)-IFN-α 2b (PEG-Intron®; Schering Corporation, Kenilworth, NJ), with or without associated lamivudine (Zeffix®; GlaxoSmithKline, Uxbridge, U.K.), and approximately 6 mo after having completed a 1-y course of therapy. This patient group is extensively described elsewhere (6). Essential clinical and virological data of the chronic hepatitis B patients are reported in Table 1. Control groups included 28 adult healthy blood donors, and 16 adults referred to the Rotterdam Municipal Health Service with a recent history of clinically evident acute hepatitis, and a positive anti-HBc IgM test (AxSym®; Abbott, Chicago, IL). Blood from these two groups was obtained devoid of any personal identification data, including date of birth. These subjects fulfilled the condition of age between 18 and 56 y, but no statistical analysis in comparison with the chronic hepatitis B group could be performed in this regard. All subjects gave written informed consent, and the protocols were approved by the local ethics committee.
Peripheral blood mononuclear cells (PBMC) were isolated from heparinized blood samples by density gradient centrifugation, aliquotted, and stored at −140°C until use.
Lymphocyte phenotyping
PBMC were thawed and stained with fluorescein isothiocyanate (FITC)-conjugated anti-CD45RA (clone HI100; BD Biosciences, San Diego, CA), phycoerythrin (PE)-conjugated anti-CD3 (clone UCHT1; Dako, Glostrup, Denmark), peridinin-chlorophyll-protein complex (PerCP)-conjugated anti-CD8 (clone SK1; BD Biosciences), and allophycocyanin (APC)-conjugated anti-CD4 (clone RPA-T4; BD Biosciences), to determine the percentage of the major T-lymphocyte subpopulations. CD8+ T lymphocytes were purified by positive selection on anti-CD8-coated magnetic beads (MACS® CD8 MultiSort kit; Miltenyi Biotec, Bergisch Gladbach, Germany), according to the manufacturer's instructions. Purified CD8+ T lymphocytes were stained with anti-CD45RA-FITC, anti-CCR7-PE (clone 3D12; BD Biosciences), and biotinylated anti-CD28 (clone CD28.2; BD Biosciences), followed by PerCP-conjugated streptavidin (SAv; BD Biosciences), and anti-CD27-APC (clone O323; eBioscience, San Diego CA). In a separate staining, purified CD8+ T lymphocytes were fixed and permeabilized with Cytofix/Cytoperm® (BD Biosciences) after staining with anti-CD45RA, anti-CD28, and anti-CD27, and further stained with PE-conjugated anti-perforin (clone δG9; BD Biosciences). For each subject, control stainings were performed with fluorochrome-matched isotype controls. The stained cells were fixed with 1% paraformaldehyde, in PBS and analyzed in a FACSCalibur® flow cytometer (BD Biosciences) using CellQuest® Pro software.
Statistical analysis
Percentages of lymphocytes expressing the respective combinations of markers in the study groups were compared by the Student's t test.
Results
The sequence of marker expression during CD8+ T-lymphocyte maturation
A schematic overview of the sequential expression of markers of CD8+ T-lymphocyte differentiation according to the model of Appay and Rowland-Jones (2) is presented in Fig. 1. This model will be referred to throughout this article. We were able to reproduce the proposed sequence of events by associating the reciprocal loss of CCR7 expression and acquisition of intracellular perforin along the pattern of co-stimulatory molecule modulation. A representative distribution of CD8+ T-lymphocyte subsets from a healthy control subject is depicted in Fig. 2.
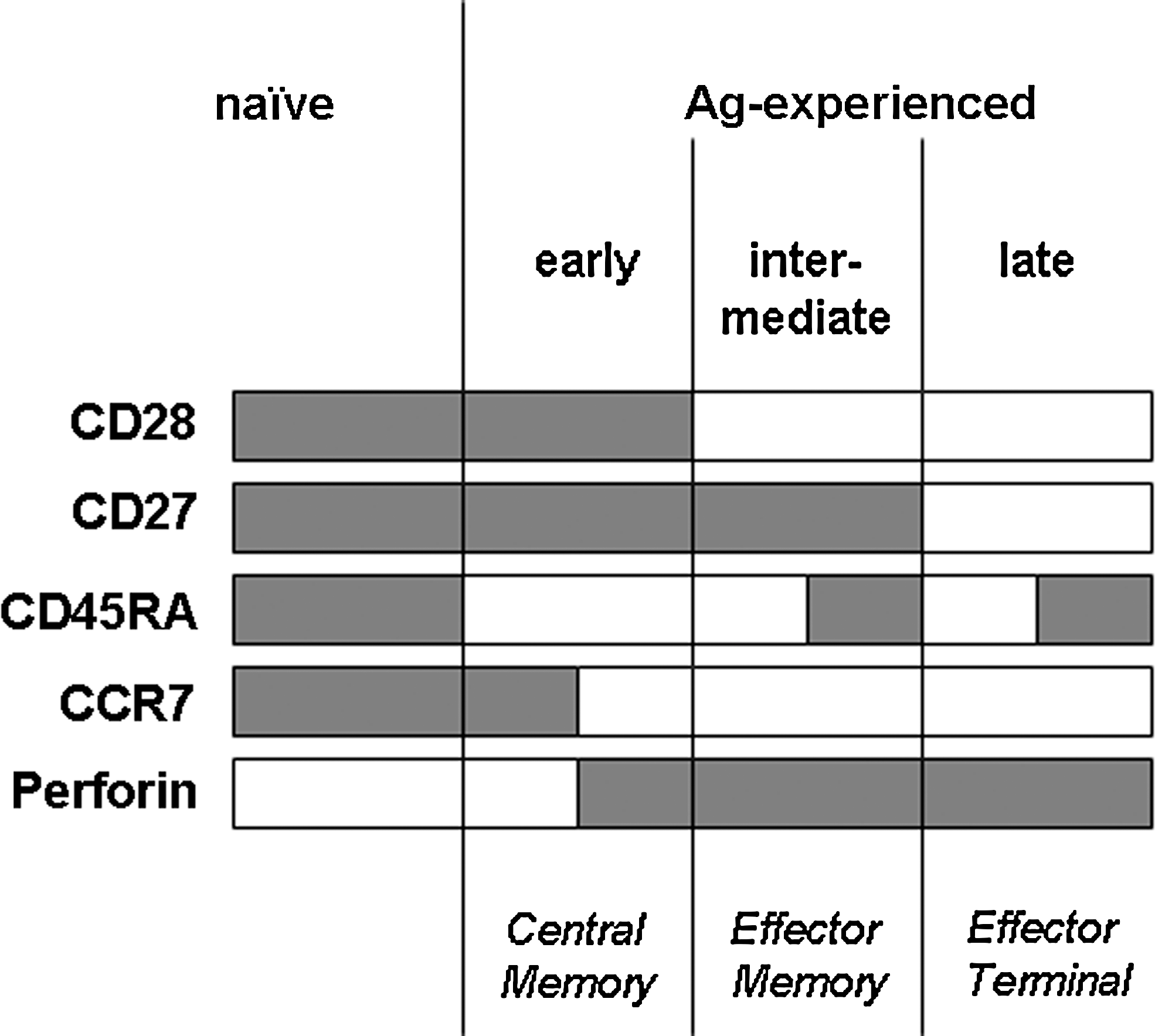
Schematic representation of marker combinations along the differentiation of CD8+ T lymphocytes according to the model proposed by Appay and Rowland-Jones (2). At the bottom of the diagram is shown a tentative correspondence to the functional definitions proposed by Lanzavecchia and Sallusto (16). Gray bars depict expression of the respective markers.
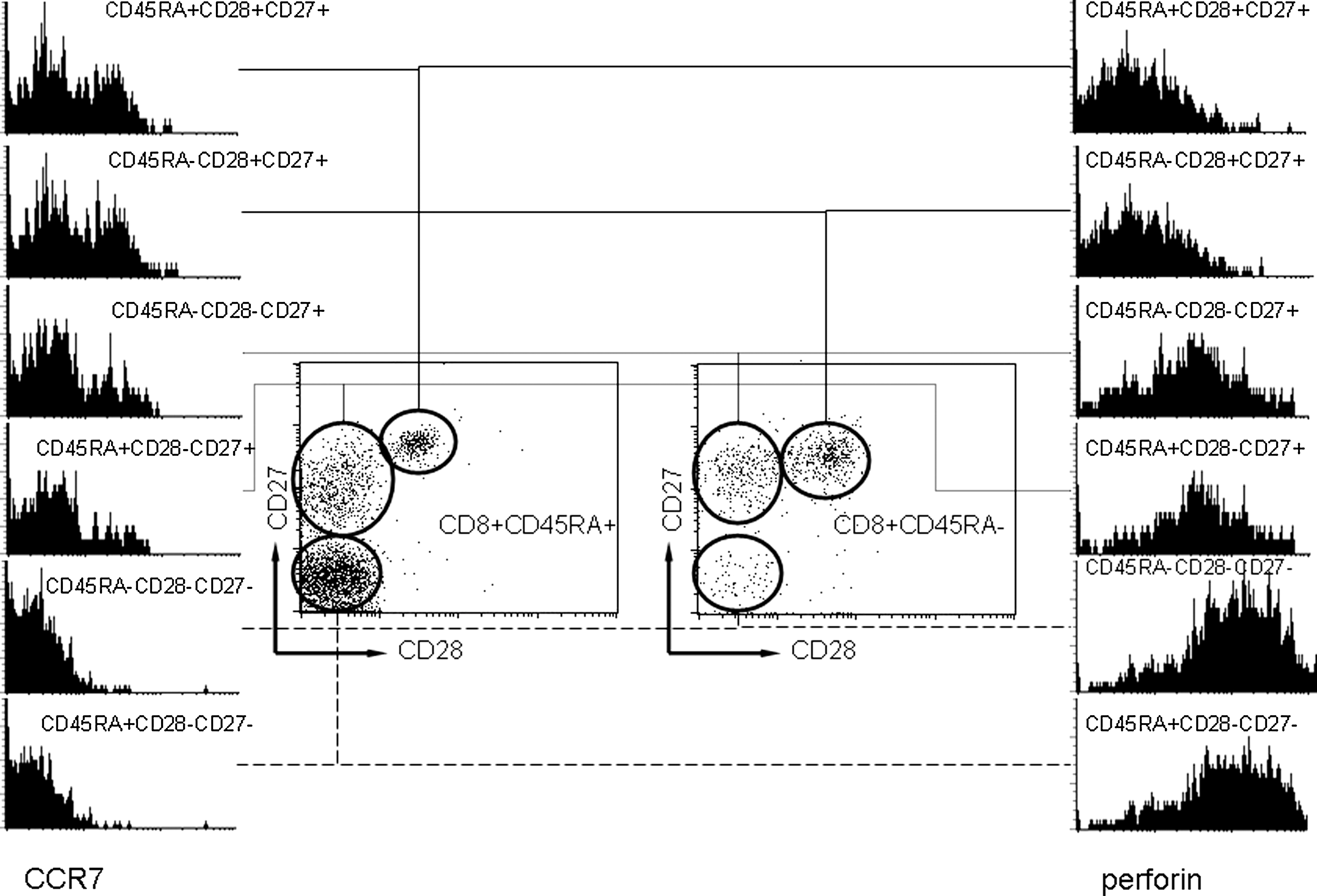
Reciprocal expression of CCR7 (left) and perforin (right) during the differentiation of CD8+ T lymphocytes. The histograms represent from top to bottom the chronological sequence of differentiation events as marked by the sequential loss of co-stimulatory molecules CD28 and CD27 (depicted in the cytograms of CD45RA+ and CD45RA−CD8+ T cells). The (re)expression of CD45RA does not significantly influence the expression of CCR7 and perforin.
Expression of CD45RA and co-stimulatory molecules
The groups of subjects included in the study did not differ significantly in the percentage of CD4+ and CD8+ T cells in peripheral blood (Table 2), even though a trend toward a relatively larger CD8+ population was observed in all HBV-infected groups. Supposedly quiescent cells, as suggested by the expression of CD45RA, represented the majority of circulating CD8+ T lymphocytes in both chronic hepatitis B patients and healthy controls, but were significantly lower in patients recovering from acute hepatitis B (Table 2).
Percentages in boldface indicate a highly significant statistical difference (p < 0.01) with healthy controls.
HBV, hepatitis B virus.
As also shown in Table 2, the expression of the co-stimulatory molecules CD27 and CD28 was then analyzed to allocate the CD8+ T lymphocytes into sequential stages of differentiation. Approximately 50% of circulating CD8+ T lymphocytes in healthy controls and in subjects recovering from acute hepatitis B showed the intermediate phenotype of antigen-experienced cells (CD27+CD28−). In contrast, this cell population was significantly less represented in chronic hepatitis B patients, regardless of the time of sampling.
Percentages of naïve and intermediately-differentiated CD8+ T cells in chronically HBV-infected patients
To assess whether the CD45RA+ cells were to be interpreted as predominantly naïve cells, the CD8+ T-cell population was further subdivided, and the percentages of the various subsets are shown in Table 3. CD45RA+ cells expressing both co-stimulatory molecules (truly naïve cells) were present in significantly higher percentages within the circulating CD8+ T cells from patients with chronic hepatitis B, compared to both healthy controls and acute hepatitis B patients (Fig. 3).
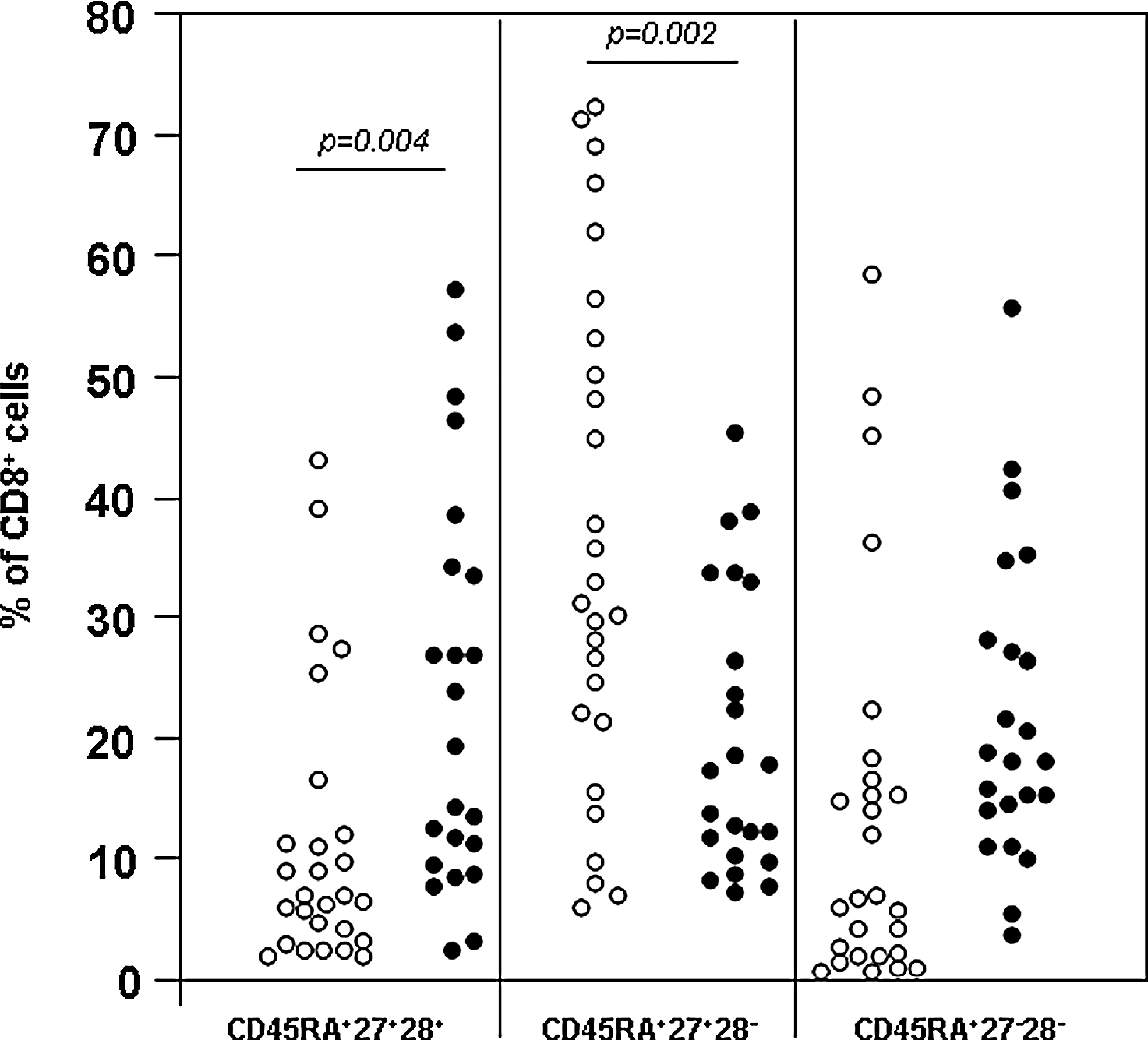
Individual percentages of CD8+ T lymphocytes expressing CD27 and CD28 in association with CD45RA in chronic hepatitis B patients (closed circles) and healthy controls (open circles).
Percentages in boldface indicate a highly significant statistical difference (p < 0.01) with healthy controls.
HBV, hepatitis B virus.
The combined analysis of expression of co-stimulatory molecules and CD45RA has also shown that the reduced percentage of intermediately-differentiated CD8+ T cells seen in chronic hepatitis B patients compared to healthy controls is essentially due to a less well represented CD45RA+CD27+CD28− population (Fig. 3). Convalescent acute hepatitis B patients had percentages of CD27+CD28−CD8+ T cells comparable to healthy controls (54.7 ± 15.7% of total CD8+ T cells), but the fraction of them not expressing CD45RA (29.9 ± 20.4% of CD8+ T cells; Table 3) was significantly higher than in both healthy controls and chronic hepatitis B patients (Fig. 4).

Individual percentages of CD8+ T lymphocytes expressing CD27 and CD28 in the absence of CD45RA in acute hepatitis B patients (closed circles) and healthy controls (open circles).
The late (CD27−CD28−) antigen-experienced CD8+ T cells from patients with HBV infection did not significantly differ from those of healthy controls. However, a trend was observed toward higher percentages of CD45RA+CD27−CD28−CD8+ T cells in chronic hepatitis B patients. The difference of the percentages of this subset between chronic and acute hepatitis B patients was indeed statistically significant (p < 0.01; Table 3).
No significant effect of treatment was seen in chronic hepatitis B patients with respect to the phenotypic markers analyzed here. Similarly, no significant difference was observed when comparing patients with different therapy outcomes (data not shown).
Expression of CCR7 and perforin in CD8+ T lymphocytes in healthy controls and chronic hepatitis B patients
The finding of significantly higher percentages of CD8+ T cells expressing the naïve phenotype in chronic hepatitis B patients led us to also study the expression of CCR7 in a smaller group of 12 such patients, and to compare them with 7 healthy controls. The data are summarized in Table 4. Even though no high statistical significance could be demonstrated, a trend toward higher expression of CCR7 on CD8+ T cells from chronic hepatitis B patients was found. In both chronic hepatitis B patients and in healthy controls, CCR7 was predominantly expressed in the early stages of T-cell maturation, as characterized by the presence of CD28. In particular, the naïve CD45RA+CD28+ subset appeared to express the highest amount of CCR7. Conversely, CCR7 was virtually absent on recently-activated intermediate and late (CD45RA−CD28−) CD8+ T cells. The early activated (CD45RA−CD28+), and the late quiescent (CD45RA+CD28−) CD8+ T cells, showed some degree of CCR7 expression, in a greater extent in chronic hepatitis B patients than in controls.
In agreement with the representative pattern shown in Fig. 2, the fraction of perforin-containing CD8+ T cells was inversely correlated with the expression of CCR7 along the differentiation phases. As for CCR7 expression, no significant difference was observed comparing these subgroups of chronic hepatitis B patients to healthy controls (Table 5).
Discussion
Skewing of lymphocyte subpopulations, often associated with impaired function, is a hallmark of immune senescence (20). Latent CMV infection, a virus known to reactivate often, and thus to challenge the immune system repeatedly, is a paradigm example of a pathogen that shapes the adaptive immunity to the point of compromising the response to other infectious agents (14,20). Hypothetically, the ability to react to infections could thus be inferred by the study of lymphocyte markers known to be associated with differing functions and stages of maturation.
The association between expansion of some lymphocyte subsets and latently or actively persisting infections needs to be tested in various circumstances before extrapolating that all persisting viruses lead to accumulation of lymphocytes at specific stages of differentiation. The present study shows that chronic active infection with HBV does not result in the accumulation of CD8+ T lymphocytes at a given maturation state. In contrast, together with a lack of markers of recent activation, CD8+ T lymphocytes from chronic hepatitis B patients show a naïve phenotype at a greater percentage than that seen in healthy controls or convalescent acute hepatitis B patients. This appears to be associated with a relative reduction of intermediately-differentiated effector-memory cells. These findings appear to be in contrast to the continuous exposure to large amounts of viral antigen typical of chronic infection with HBV. One possible interpretation could be provided by a mouse model of chronic polyomavirus infection, in which in the face of ongoing re-encounter with antigen, continuous recruitment of naïve cells has been demonstrated (25). Assuming that continuous activation of CD8+ T lymphocytes occurs in response to the rapidly-replicating virus, the expansion of the naïve phenotype suggests a rapid elimination of activated cells.
The relationship between the percentages of lymphocytes bearing defined phenotypes and the amount of lymphocytes specific for given pathogen-associated antigens is not fully understood. The allocation of memory and effector T cells in relation to infections have mostly been derived from the characterization of virus-specific CD8+ T cells. Although major expansions of virus-specific T lymphocytes have been demonstrated during acute infections, the percentage of cells specific for a given epitope can hardly explain the significant changes seen in lymphocyte subsets. The currently used MHC tetramer-peptide complexes have indeed allowed us to elucidate fine mechanisms of antigen-specific immune responses, but they fail by their very nature to provide a complete picture of the reacting immune system. By concentrating attention on those responses characterized by the highest avidity of the receptor involved, the participation of large numbers of lymphocytes exhibiting weaker reactions goes unchecked. Those less stringent, probably cross-reactive, lymphocytes may in turn show faster and more efficient patterns of reaction, due to previous exposure to other unrelated antigens. The present data suggest that during chronic infection with HBV, the skewing of CD8+ T lymphocytes toward the naïve phenotype may be driven by the elimination, or compartmentalization, of cells activated as a result of the persisting virus. This contrasts with the accumulation of lymphocytes in subsets corresponding to the phenotype of the virus-specific cells. HBV-specific CD8+ T cells are readily detected in the circulation of subjects spontaneously resolving acute hepatitis B infection (9,12,21,26). These cells show a phenotype similar to that of hepatitis C virus-specific CD8+ T cells during acute infection with this virus (24). This finding suggests that HBV is perfectly capable of inducing a vigorous virus-specific response. In contrast, chronic infection with HBV is characterized by limited and weak virus-specific T-cell responses, and HBV-specific CD8+ T cells in chronically-infected subjects are hardly detectable directly ex vivo (4), requiring in vitro expansion to be visualized (6,26). For this reason, it has been practically impossible to establish the phenotype of the fully differentiated HBV-specific CD8+ T lymphocytes seen during chronic infection, in contrast to what has been extensively reported for other viral infections (22).
A factor that could play a role in this scenario is the migration of HBV-specific lymphocytes to the liver. HBV-specific CTL have been demonstrated infiltrating the livers of chronic hepatitis B patients. The increase in infiltrating lymphocytes during flare-ups of hepatitis, however, appears to be mainly due to an influx of non-HBV-specific lymphocytes, as assessed by MHC-tetramer complexes, rather than expansion of the specific ones (7,18).
The low percentage of CD27+CD28− CD8+ T lymphocytes found in chronic hepatitis B patients, as hypothesized above, can result from increased influx of naïve T cells, or less likely, the accumulation of terminally-differentiated cells. However, either because of active replenishing or a passive relative increase of naïve cells, continuous loss of CD8+ T cells in the intermediately-differentiated subset seems to be the most plausible explanation for the observed phenotype distribution. The early and intermediate subsets, characterized by the maintenance of CD27 expression, appear to include the effector and memory cells generated during most viral infections, either acutely resolved or persisting (22). Consistent with these findings, the healthy controls and patients acutely infected with HBV examined in the present study showed similar percentages of circulating CD27+CD28−CD8+ T cells. The only difference observed resides in the CD45RA− fraction, which was larger in the acutely HBV-infected patients, most likely reflecting recent activation.
In conclusion, the results of the present study suggest that the persistence of HBV is not associated with exhaustion or early senescence of the lymphocyte pool. However, patients chronically infected with HBV fail to generate a steady pool of intermediately-differentiated memory CD8+ T lymphocytes. Further studies are warranted to establish whether lymphocyte subset patterns are associated with clinically relevant events in these patients.
Footnotes
Acknowledgments
The authors would like to acknowledge the precious collaboration of the Department of Infectious Diseases of the Rotterdam Municipal Health Service and the Department of Gastroenterology of the ErasmusMC University Hospital for the access to blood samples from patients with acute and chronic infection with HBV, respectively.
Financial support was obtained from the Landsteiner Stichting voor Bloedtransfusie Research (Amsterdam, The Netherlands).
Author Disclosure Statement
No competing financial interests exist.