
Abstract
In Graves' disease, the IgG class autoantibody against thyrotropin receptor (TRAb) is produced excessively and induces hyperthyroidism. Epstein-Barr virus (EBV) is one of the human herpesviruses that persists for life, mainly in B lymphocytes, and is occasionally reactivated. Therefore, EBV may affect the antibody production of B lymphocytes that would normally produce TRAb. The purpose of the present study was to evaluate the association of EBV reactivation with the etiology of Graves' disease. Serum levels of EBV antibodies and IgE were determined by ELISA. TRAb levels were determined by radioreceptor assay. We performed in-situ hybridization (ISH) of EBV-encoded small RNA (EBER)1 on the thyroid tissue of one of our patients. In Graves' disease patients with TRAb levels ≥10%, EA antibody levels, which indicate EBV reactivation, were moderately but significantly correlated with the levels of TRAb, and weakly but significantly correlated with IgE. EBER1-ISH revealed that one of our patients had EBV-infected lymphocytes infiltrating the thyroid gland. EBV reactivation may stimulate antibody-producing B lymphocytes predisposed to make TRAb, and this may contribute to or exacerbate the disease.
Introduction
Epstein-Barr virus (EBV) is a human γ-herpesvirus and its genome consists of double-stranded DNA approximately 170 kbp in size (15). EBV is widespread in all human populations, with more than 90% of adults positive for serum antibodies. EBV causes infectious mononucleosis during its primary infection, and it also is the human tumor virus that is detected in subsets of Burkitt's lymphoma, Hodgkin's lymphoma, nasopharyngeal carcinoma, and gastric cancer or other malignancies (23).
EBV persists, mainly in human B lymphocytes, for life, and is occasionally reactivated. Therefore viral latency or intensive reactivation may have effects on the antibody production of B lymphocytes, and a contribution of EBV to various autoimmune diseases has been suggested (2,4,5,9,14,17 –20,22,25,30 –32,34). For example, in multiple sclerosis (MS), and systemic lupus erythematosus (SLE), EBV infection is thought to be an important environmental trigger (4,22,34). In SLE, it has been reported that EBV early antigen (EA) antibody is high in patients with cutaneous symptoms or increased anti-Ro antibody, and viral capsid antigen (VCA) antibody is high in patients with joint involvement (34).
We found several cases of exacerbation of Graves' disease accompanied by high levels of EA antibody. EA is the product of the BMRF1 gene in the EBV genome (6,10,21). BMRF1 protein is expressed in the early phase of EBV reactivation. We thus sought to study the role of EBV reactivation in Graves' disease.
In in-vitro experiments, it has been shown that EBV can induce Ig production in infected B lymphocytes (13,27,29). Therefore, EBV may be related to Graves' disease through stimulation of autoantibody production in B lymphocytes attracted to TRAb.
In this study, we hypothesized that TRAb production might be affected by EBV reactivation, and examined serum levels of TRAb, anti-EBV antibodies, and IgE, the most enhanced isotype. We examined whether EBV-positive lymphocytes actually existed in the thyroid tissue of a Graves' disease patient, using EBV-encoded small RNA (EBER)1 in-situ hybridization (ISH).
Materials and Methods
Subjects
Sixty-six Graves' disease patients and 32 healthy control subjects participated in the study. They all gave informed written consent, and the study protocol was approved by the Medical Ethics Committee for Human Subject Research at the Faculty of Medicine, Tottori University, Yonago, Japan.
At the time of diagnosis, the patients had symptoms including at least one of the following: (1) signs of thyrotoxicosis such as tachycardia, weight loss, finger tremor, and sweating, (2) diffuse enlargement of the thyroid gland, (3) exophthalmos and/or specific ophthalmopathy, and all patients also met three laboratory-determined criteria: (1) elevation in serum free T4 and/or free T3 levels, (2) suppression of serum thyroid-stimulating hormone (TSH) (<0.1 μU/mL), and (3) positive for TRAb or thyroid-stimulating antibody (TSAb). Fifty-four patients showed elevated radioactive iodine uptake by the thyroid gland, and in 12 patients the iodine uptake was not tested.
Twenty-two patients had had no treatment, and 44 patients were undergoing treatment with antithyroid drugs (methimazole or propylthiouracil).
Control subjects were selected from healthy laboratory staff. Their thyroid function was normal, and subjects that had any familial history of thyroid disease were excluded.
The mean ages (±SD) of the subjects were 41.0 ± 13.5 y for Graves' disease patients, 40.3 ± 14.3 y for those with no treatment, 41.4 ± 13.2 y for those under treatment, and 34.7 ± 16.9 y for normal subjects. The clinical backgrounds of the study subjects are shown in Table 1. There were no subjects with clinically obvious EBV primary infection at the time of sampling.
Sample analysis
Peripheral blood samples were obtained from patients and control subjects. The sera were separated and stored at −80°, and sample analyses were performed soon after collection.
Levels of anti-EBV IgG antibodies against EA, viral capsid antigen (VCA), and EBV nuclear antigen (EBNA) in sera were determined by ELISA (EA, Vircell, Granada, Spain; VCA and EBNA, Panbio, Brisbane, Queensland, Australia). For EA antibody, intra-assay coefficient of variation (CV) was 9.3%, and inter-assay CV was 9.6%. For VCA antibody, intra-assay CV was 3.6%, and inter-assay CV was 1.3%. For EBNA antibody, intra-assay CV was 7.8%, and inter-assay CV was 1.7%.
Anti-EBV antibodies were tested for early antigen-diffuse type (EA-D, a BMRF1 gene product), VCA-p18 (a BFRF3 gene product), and EBNA1 (a BKRF1 gene product).
Serum TRAb levels were determined by radioreceptor assay (SRL, Inc, Tokyo, Japan). Serum samples were incubated with solubilized porcine TRH receptors for 15 min at room temperature. 125I-labeled bovine TSH was added, and was incubated 2 h more at room temperature. After adding polyethylene glycol, the mixture was centrifuged at 1500 × g for 30 min to precipitate the receptor-tracer conjugates. The radioactivity in each pellet was measured (24). Intra-assay CV was 1.35%, and inter-assay CV was 1.05%. We adopted 10% as the clinical cut-off level of TRAb.
Serum IgE levels were determined by ELISA with human IgE antibody (Bethyl Laboratories, Inc., Montgomery, TX) as capture antibody, and HRP-conjugated human IgE antibody (Bethyl Laboratories) as the detection antibody. Intra-assay CV was 2.5%, and inter-assay CV was 5.6%.
According to the manufacturer's instructions, we measured each control serum, and determined the cut-off values for EA, VCA, and EBNA antibodies as 0.8, 1.0, 0.9 ??? (OD 450 nm), respectively. The clinical cut-off value for IgE was 170 IU/mL, so we adopted 408 ng/mL for our analysis (1 IU/mL = 2.4 ng/mL).
All sample sera were checked for the hepatitis C virus (HCV) antibody by qualitative analysis using the third-generation HCV antigen (Ortho Clinical Diagnostics, Tokyo, Japan) (Table 1).
In-situ hybridization of EBV-encoded small RNA1
We performed the EBER1 ISH on the thyroid tissues of one of our patients (under treatment, TRAb 73%, EA 1.27 ???), who underwent partial thyroidectomy. The section was deparaffinized, dehydrated, and treated with proteinase K (Dako, Tokyo Japan) for 10 min at room temperature. After washing, FITC-conjugated peptide nucleic acid (Dako) complementary to EBER1 was applied and hybridized overnight at 37°C, then washed with standard saline citrate. Detection was performed with alkaline phosphatase-labeled anti-FITC antibody (Dako) for 30 min at room temperature, and BCIP/NBT for 7 h at room temperature. Methyl green was used as the counter-stain.
Statistical analysis
Statistical analysis was performed using Stat View software (SAS Institute, Inc., Cary, NC). We adopted the non-parametric Spearman's rank correlation test for examination of the relationship between the two groups, and Fisher's exact test for comparison of the positive rates of antibody levels.
Results
Anti-EBV antibodies, IgE, and TRAb levels
The prevalence rates of EBV antibodies, IgE, and TRAb in the tested subjects are shown in Table 2. The rate of patients who tested positive was significantly higher than that of control subjects, for EA antibody (p < 0.0001), VCA antibody (p = 0.0127), EBNA antibody (p = 0.0015), and TRAb (p < 0.0001). However, for IgE, the rate of control subjects who tested positive was significantly higher than that of the patients.

*Statistically significant.
The prevalence of TRAb in the patients with no treatment was significantly higher than that seen in the patients under treatment (p = 0.0255).
Relationship between IgE levels and EA antibody levels
A weak but significant correlation (Spearman's rank correlation coefficient [rs] = 0.318, p = 0.0371, regression curve from x to y: y = 104.73 + 209.93 x) was detected in Graves' disease patients with TRAb levels ≥10% (Fig. 1a), but in the patients with TRAb levels <10%, we could not detect any relationship (rs = 0.015, p = 0.9443; Fig. 1b).
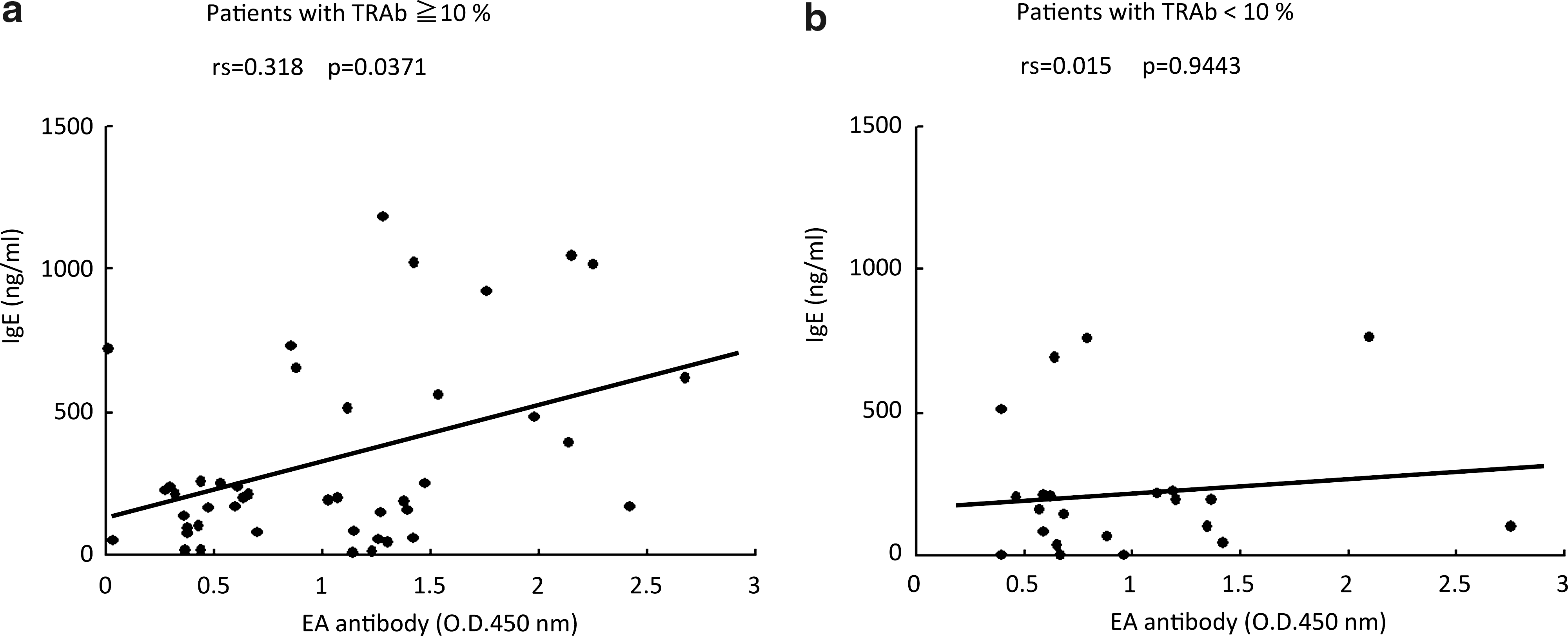
Distribution of early antigen (EA) and IgE antibody levels in Graves' disease patients: (
Relationships between TRAb and EA antibody levels
Though there was no correlation between TRAb and EA antibody levels of all Graves' disease patients, in the patients with increased TRAb (cut-off level: 10%), we detected a moderate but significant positive correlation (rs = 0.443, p = 0.0037, n = 44, regression curve from x to y: y = 23.717 +12.729 x; Fig. 2a). We also found a moderate positive correlation for patients under treatment, with increased TRAb levels (rs = 0.461, p = 0.0239, n = 25, regression curve from x to y: y = 19.934 + 15.236 x; Fig. 2b), and in patients with no treatment with increased TRAb levels (rs = 0.521, p = 0.0230, n = 19, regression curve from x to y: y = 24.877 + 11.756 x; Fig. 2c). In comparison to those relationships, in the normal controls increased EA antibody levels were not related to TRAb levels (Fig. 2d).

Distribution of anti-thyrotropin receptor antibody (TRAb) and early antigen (EA) antibody levels: (
Relationships between TRAb levels and EBNA or VCA antibody levels
We found no correlation between TRAb and EBNA (rs = 0.143, p = 0.2526), or VCA (rs = 0.127, p = 0.3088) antibodies (data not shown).
In-situ hybridization of EBV-encoded small RNA1
We could obtain only one sample for ISH, and we detected some EBER1-positive lymphocytes in the thyroid tissue from the Graves' disease patient (Fig. 3).
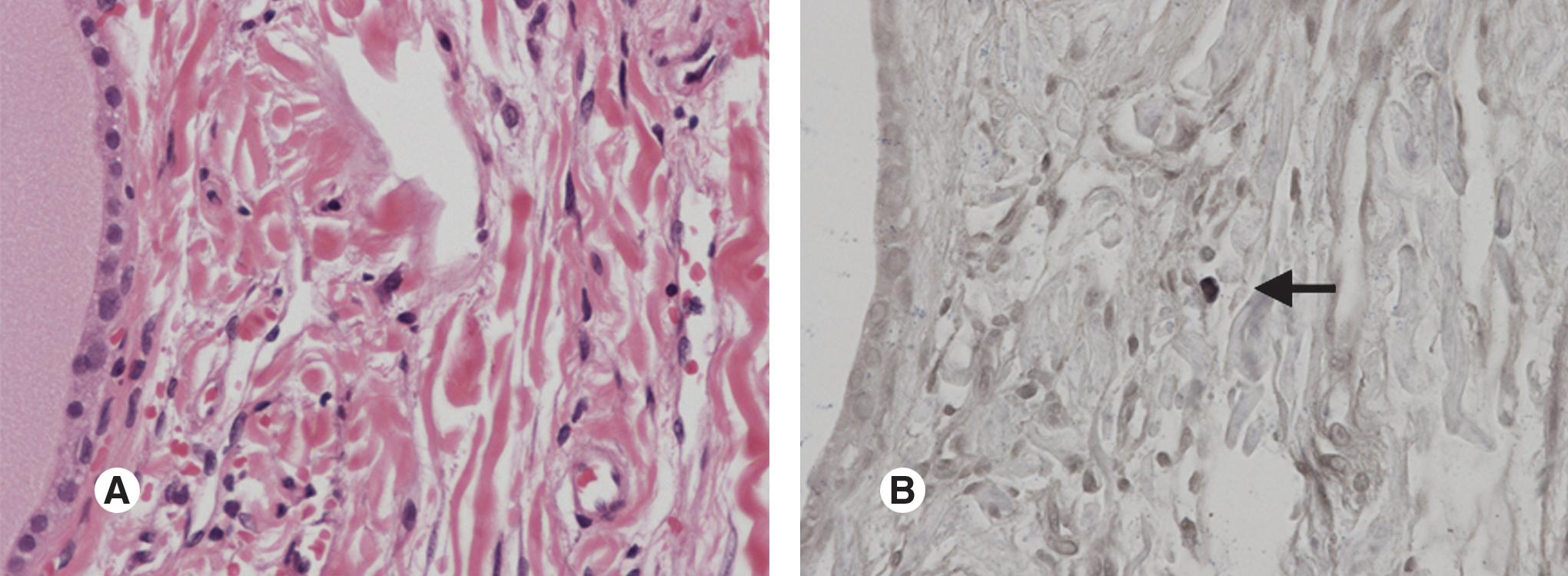
One of the EBV-encoded small RNA1 (EBER1)-positive lymphocytes infiltrating (arrow) the thyroid gland in a Graves' disease patient: (
Discussion
Several reports have shown the contribution of EBV to the antibody production of B lymphocytes, and EBV is thought to be a B-lymphocyte activating factor (13,27,29). When EBV-infected B lymphocytes are in the presence of interleukin-4, the production of every isotype of Ig is increased, and the most enhanced isotype is IgE (27). The class switch recombination is induced for IgE (13). Therefore, EBV could be a factor stimulating B lymphocytes to produce IgG with IgE.
Clinically, it has been reported that Graves' disease patients also often have allergic disease or high serum levels of IgE (1,11,27,29,33), and during anti-thyroid drug treatment the levels of TRAb decrease less, and the remission rate is lower in patients with IgE elevation (16).
Another report showed that B cells of Graves' disease patients were predisposed to the production of TRAb (8). When EBV is reactivated and stimulates B cells that are predisposed to TRAb production, both IgE and TRAb may be increased.
In this study, we measured the serum levels of EA antibody. EA is expressed during primary infection with EBV, or in the early phase of viral reactivation. In our study, the EA level represents the extent of EBV reactivation, because in all of the patients and the normal subjects we studied, none were in a state of EBV primary infection. Thus we determined serum IgE levels and TRAb levels in the same serum samples.
There are many factors that cause exacerbations of Graves' disease and allergic diseases, and if EBV stimulation induces both IgE and TRAb production, this is only one of many stimulating factors. Therefore, in those with low levels of TRAb production, other factors must be dominant, and ascertaining the precise relationship between EBV antigen and Ig production is difficult.
Our investigation showed a weak but significant correlation between serum EA antibody and IgE levels in Graves' disease patients with TRAb levels ≥10% (Fig. 1) suggesting that with increased TRAb production, serum IgE increases in proportion to the increase in EA antibody. On the other hand, in patients with low TRAb levels, we could not detect any relationship between IgE and EA antibody levels.
Also, for TRAb and EA antibody we found a moderate but significant correlation, only in patients with TRAb levels ≥10%. As shown in Fig. 2, there were some patients with increased TRAb in spite of low EA levels, because other factors affect increases in TRAb. However, there were also some patients with low TRAb and high EA levels, and they also had TRAb levels <10%. We could explore the relationship by excluding these data points.
One of the putative causes of low TRAb and high EA levels is the effect of therapy. Usually TRAb levels in Graves' patients are gradually reduced by treatment with anti-thyroid drugs. In our study, the patients shown in Fig. 2b (those under treatment), and 2c (those with no treatment) were separated. However, most patients with low TRAb and high EA levels were distributed among those with TRAb levels <10%, so during treatment the TRAb levels of these samples were below the cut-off level (10%), in spite of the fact that EA levels remained high. Another factor is the individual differences in the numbers of TRAb-producing cells. Serum TRAb levels correlate with the percentage of B cells committed to TRAb production, even though the stimulation by EBV is the same.
These results suggest that in a certain group of Graves' disease patients, EBV reactivation stimulates antibody-producing B cells to induce Ig production, and serum TRAb along with IgE levels increase.
We also measured VCA antibodies, but could not detect any associations with TRAb. VCA is the late protein of reactivation, so antibody against VCA increases in serum during the late phase of viral reactivation. However, once it is expressed, it remains in the serum for a long time, as Henle et al. indicated in their report (10). Therefore in adults, VCA reflects past infection rather than the extent of reactivation. VCA is neither related to the extent of reactivation nor to TRAb.
Viral reactivation results in the production of replicated viral particles. In some patients, especially those with high VCA antibody levels, the reactivation may progress to the late phase, and there may be free viral DNA or particles in the serum. Therefore, we tried to detect viral DNA in the serum samples, but we could not detect any EBV DNA (data not shown). These results suggest that there was no detectable free viral DNA or particles in the patients' sera.
On the other hand, one of our patients had EBV-infected lymphocytes infiltrating the thyroid gland (Fig. 3), which supports an influence of EBV on the thyroid gland, and suggests the possibility of EBV reactivation in the thyroid gland, although EBER is expressed in latently-infected cells. If we had detected reactivation proteins in the thyroid gland, this would have been stronger evidence of a connection, but unfortunately reactivation proteins such as ZEBRA were not detected (data not shown). This is because lytically-infected cells usually appear less frequently than latently-infected cells. However, it is important that EBV-infected lymphocytes were present in the patient's thyroid tissue.
It is unclear how virus reactivation causes antibody production. It may simply increase viral copy numbers, and consequently affect virus-infected cell numbers, or it may stimulate the antibody production system via some other pathway. There have been many reports (2,5,9,17 –20,25,29 –32,34) concerning which is more important for antibody production: latent infection or reactivation. For example, it has been reported that EBV-latent membrane protein (LMP)1 mimics CD40 signals, to induce B-lymphocyte activation and differentiation continuously (29), and the expression of the EBV reactivation proteins EA and VCA have been shown to be related to the symptoms of SLE (34). Positive EBV-latent protein EBNA is a known risk factor for MS (2,5,9,17,18,20,22,25,32), but the level of EBNA antibody was not related to the TRAb level in our study. An association of EBV with MS has long been suggested, and recently the EBV-infected autoreactive B-cell hypothesis was proposed (27). In genetically-susceptible individuals, EBV-infected autoreactive B cells seed the target organ, where they produce pathogenic autoantibodies and send survival signals to autoreactive T cells. This hypothesis is compatible with our hypothesis; in a subset of Graves' disease patients, EBV reactivation stimulates antibody-producing B cells predisposed to TRAb.
In addition to EBV, there are some other chronically-infecting viruses. Cytomegalovirus, herpes simplex virus (HCV)-1 and HCV-2, and human herpesviruses 6 and 7 were all reported to be unrelated to autoimmune thyroid disease (26). Hepatitis B virus is not thought to be related to thyroidal autoimmunity, since the levels of thyroidal autoantibodies in hepatitis B patients were lower than those of normal controls (12). JC virus is unlikely to cause thyroidal autoimmunity, because it has tropism to the central nervous system, not to the thyroid gland (3). Hepatitis C virus has been reported to be related to thyroidal autoimmunity (12,28). Of our subjects, two of the Graves' patients and none of normal controls tested positive for HCV-antibody, but this difference was not significant (p > 0.9999). Thus HCV may not be related to Graves' disease.
Graves' disease is a multi-factorial disease, and many studies are exploring it, both genetically and environmentally. It is likely that EBV is not itself the cause of Graves' disease. However, it influences TRAb production, and may thus affect the progression and exacerbation of Graves' disease. This issue requires more investigation, and it may lead to therapies that help prevent Graves' disease, as well as other autoimmune diseases.
Footnotes
Acknowledgments
The authors thank Drs. Chiaki Shigemasa, Shunsuke Meshitsuka, Kazuhide Ogino, and Shinichi Taniguchi, for their valuable advice and encouragement.
Author Disclosure Statement
No competing financial interests exist.