
Abstract
Human metapneumovirus (hMPV) has been recognized as an important respiratory pathogen. Due to its relatively recent discovery, only limited information is available on the relationship between hMPV and type I interferons (IFN). This study was designed to determine whether in vitro hMPV is sensitive to the antiviral activity of IFN-β, leukocyte IFN-α, and several IFN-α subtypes in a human Hep-2 cell line. The results showed that 50% inhibitory concentration values against hMPV for the various type I IFN preparations were significantly higher than those against the IFN-sensitive vesicular stomatitis virus, and some IFN-α subtypes appeared to be more active against hMPV than others, with IFN-α subtypes 5, 6, 8, and 10 being the most potent, and IFN-α2, 17, and 21 the least potent. The results show that hMPV grown in Hep-2 is partially resistant to the antiviral activity of type I IFNs. Additional studies are required to understand whether and to what extent the relatively low sensitivity of hMPV to IFNs influences the clinical outcomes of infected individuals.
Introduction
As with respiratory syncytial virus (RSV), airway epithelial cells are the primary target for hMPV infection. However, to date little is known about hMPV pathogenesis and the activation of the innate immune response triggered by infection, and more specifically, about the relationship between type I interferons (IFN) and hMPV.
Previous studies have shown that infection of A549 cells (human lung adenocarcinoma epithelial cell line) with hMPV results in significant changes in type I IFN and IFN-stimulated gene transcription (2). It has also been reported that hMPV is not sensitive to IFN-α-mediated clearance, and that IFN-α-mediated gene expression is inhibited by hMPV infection (6). However, other studies have suggested that hMPV is more susceptible to IFN-α than is RSV, although both viruses appear to cause similar clinical manifestations and have similar epidemiological features (2,12). In addition, recently it has been shown that type I IFN renders epithelial cells of the respiratory tract resistant to viral infections (18). Hence, it is important to investigate hMPV interactions following infection, particularly the mechanism by which the host is capable of clearing the virus.
In light of these facts, we investigated whether in vitro hMPV is sensitive to the antiviral activity of IFN-β, leukocyte IFN-α, and several IFN-α subtypes. The sensitivity of hMPV was compared with that of vesicular stomatitis virus (VSV), which is known to be highly sensitive to the action of all types of IFN.
Materials and Methods
The hMPV [strain NL-001, group A1 (27)] was propagated and titrated in monkey kidney LLC-MK2 cells, while VSV (Indiana strain) was propagated and titrated in L929 mouse fibroblasts. To enable careful examination of the effects of IFNs on hMPV, initial experiments were conducted to characterize the growth of hMPV in a human Hep-2 cell line. The infection was initiated by inoculation of the stock virus at an input multiplicity of 0.1 tissue culture infection dose 50% (TCID50)/cell, both in the presence and absence of exogenous trypsin. The low multiplicity of infection (MOI) is due to the fact that hMPV propagated in LLC-MK2 cells replicates to a titer of 106 TCID50/mL, either in the presence or absence of exogenous trypsin at a concentration of 2 μg/mL, as suggested by Tollefson et al. (25).
After 1 h of adsorption, the hMPV inoculum was washed away and the cells were cultured. Trypsin was added during infection and every other day at a concentration of 0.5 μg/mL because we observed that at concentrations of 1 μg/mL or more it was toxic to the Hep-2 cell line. Indeed, at a trypsin concentration of 1 μg/mL the Hep-2 cells detached from the plate, as observed by Tollefson et al. (25).
Results and Discussion
Aliquots of the cells and the cell-culture fluid were collected at the time of virus absorption (i.e., 1 h post-infection), and at 2, 4, 8, 24, 48, 72, and 96 h post-virus infection, for experiments performed both with and without trypsin, and were titrated in LLC-MK2 cells by determination of the TCID50/mL using Reed and Muench's method (20). The kinetics of hMPV growth in Hep-2 cells in the above conditions are shown in Fig. 1. Each point represents the mean value of two separate experiments run in duplicate, with a standard deviation never exceeding log 0.3. It shows that in the experiments in which trypsin was not added, an hMPV virus titer of 103 TCID50/mL was present in the extracellular fluid after 24 h of Hep-2 cell-infection, which peaked 24 h later at approximately 103.43 TCID50/mL. The hMPV titer then slowly declined over the next 2 d. Conversely, the cell-associated virus titer was 102.2 TCID50/mL 4 h after the absorption period, and rose rapidly to 104.4 TCID50/mL between 8 and 24 h. The cell-associated virus titer remained relatively stable for about 24 h, and then declined over the next 2 d. During the productive phase of replication the cell-associated virus titer was approximately log 1 higher than the titer of the virus released in the medium. No differences in the kinetics and titers of hMPV growth in Hep-2 cells were observed when the same procedures were undertaken during infection, and every other day trypsin was added at a concentration of 0.5 μg/mL.
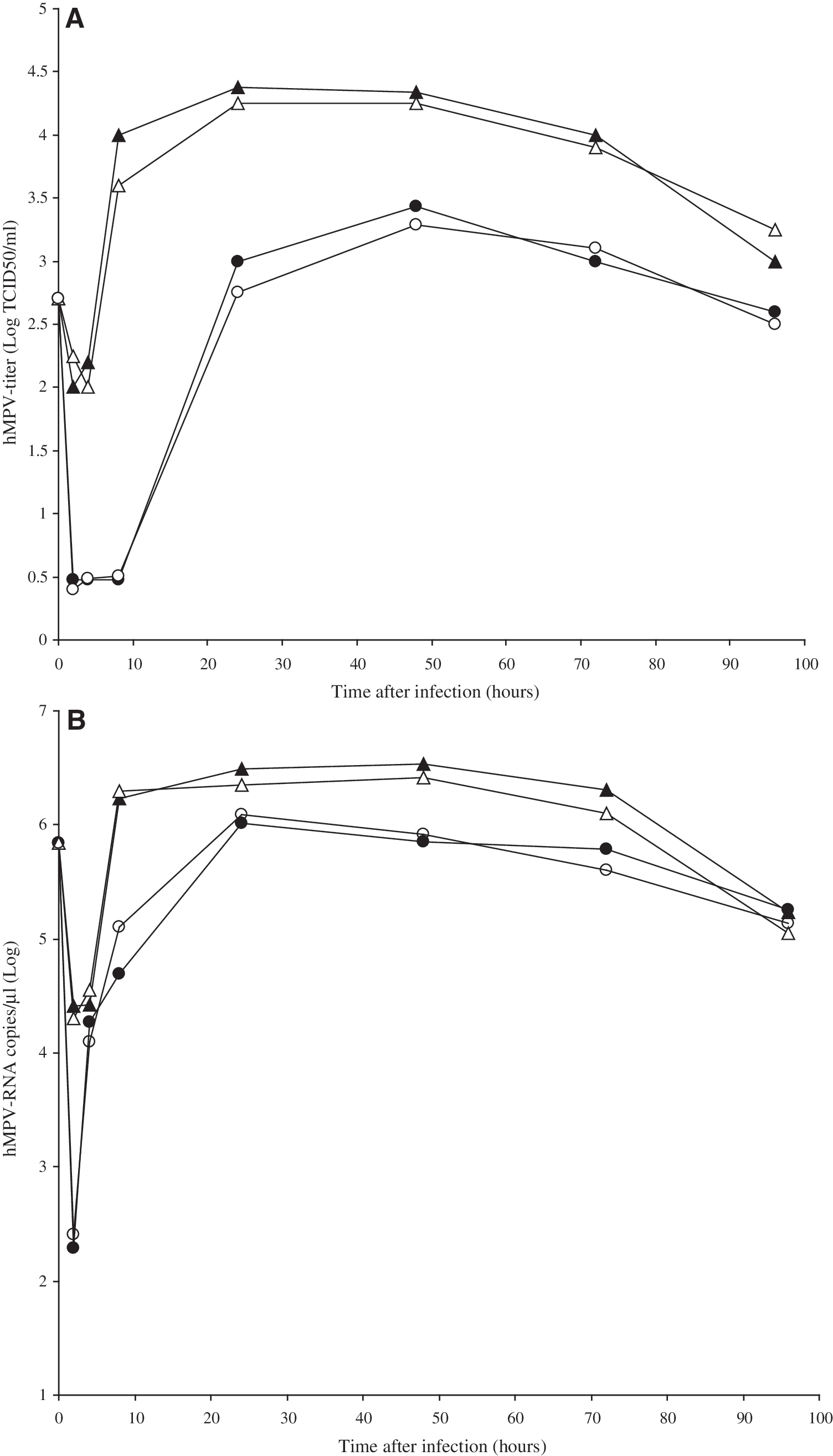
The human metapneumovirus (hMPV) replication cycle. Human Hep-2 cell lines were infected with hMPV at a multiplicity of infection of 0.1 tissue culture infection dose 50% (TCID50)/cell in the presence or absence of exogenous trypsin (0.5 μg/mL), and at the time of virus absorption and at 2, 4, 8, 24, 48, 72, and 96 h post-virus infection, pelleted cells (black triangles = without trypsin, white triangles = with trypsin), or supernatants (black circles = without trypsin, white circles =with trypsin) were separately collected and titrated for virus content using either yield assay (panel
Similar results were also obtained when levels of hMPV-RNA were measured using reverse transcriptase (RT) real-time PCR after the following experimental procedure. Viral RNA was extracted from aliquots of the cells or supernatants collected at the time of hMPV absorption, and at 2, 4, 8, 24, 48, 72, and 96 h post-virus infection using a QIAamp viral RNA Mini Kit (Qiagen Spa, Milan, Italy), in experiments undertaken with or without trypsin as described above. The RNA was dissolved in RNAse-free water, and hMPV quantification using real-time PCR (TaqMan assay) was undertaken after generation of cDNA using a high-capacity cDNA Archive Kit (Applied Biosystems, Monza, Italy). The primers and probe for the N gene of hMPV (16) were added to the universal PCR master mix (Applied Biosystems) at 300 and 100 nM, respectively, in a final volume of 50 μL. The standards were obtained by cloning the 212 bp of the viral N gene into the pCR2.1 plasmid using a TOPO TA cloning kit (Invitrogen Corp., Carlsbad, CA). A linear distribution (r = 0.99) between 101 and 108 copies of hMPV standard was obtained.
The results (Fig. 1B) showed that in the Hep-2 cell line without trypsin, an hMPV-RNA level of 104.3 copies/μL was measurable in extracellular fluid at 4 h post-infection, peaking 20 h later, and then slowly declining over the next 3 d. In contrast, production of cell-associated hMPV-RNA took place relatively quickly, reaching a titer of 106.5 copies/μL in about 8 h, with the highest titer seen 24 h after Hep-2 cell infection. Again, no difference in kinetics and titers of hMPV growth in Hep-2 cells, measured as the number of copies of hMPV RNA/mL, were observed when the same procedures were undertaken using trypsin after infection and every other day at a concentration of 0.5 μg/mL.
Having established that hMPV replication in Hep-2 cells was completed in about 48 h, and that after 24 h the peak yield was reached, regardless of the use of trypsin under experimental conditions, the latter time point was chosen to test the antiviral activities of the interferons. We tested the effects on hMPV replication of IFN-β (IFN-β1a, specific activity 270 million international units [MIU]/mg; Serono Europe Limited, London, U.K.), leukocyte IFN-α (specific activity 200 MIU/mg; Alfa Wassermann, Bologna, Italy), and different subtypes of IFN-α, namely IFN-α1, 2, 5, 6, 7, 8, 10, 14, 17, and 21 (specific activities 50–250 MIU/mg; all supplied by PBL Biomedical Laboratories, Piscataway, NJ). All experiments assessing the sensitivity to IFNs were performed without adding trypsin during or after hMPV infection of Hep-2 cells on the basis of the results of the characterization of the replication cycle of hMPV in Hep-2 cells. Furthermore, a single nucleotide change was found in our NL-001 strain, that encoded the amino acid substitution S101P at the putative cleavage site of the fusion glycoprotein that has been reported to be a major determinant of trypsin-independent growth of hMPV in tissue culture (24) (data not shown).
Briefly, Hep-2 cells were seeded at 3 × 104 cells per well in 96-well plates, and after 24 h cell triplicate cultures were incubated with serial dilutions (range 0–500,000 IU/mL) of each of the IFNs for 24 h. In each test, 12 wells were filled with 0.1 mL of medium to serve as both virus and cell control. Hep-2 cell monolayers were then washed with MEM, and both IFN-treated and control viruses were inoculated with hMPV at an MOI of 0.1, or 0.01 TCID50/cell. After adsorption at 37°C for 1 h, the excess virus inoculum was removed, the cell monolayers were washed with PBS, and the wells were topped up with complete medium to 0.1 mL/well. Infection with hMPV took place over 24 h at 37°C. Culture supernatants were then collected, and the cells were subjected to three consecutive freeze-thaw cycles at −80°C. The culture supernatant was then added to the cell lysate and titrated for hMPV yield in LLC-MK2 cells by determination of the TCID50/mL. The concentration that inhibited 50% of the hMPV yield (i.e., the IC50) was calculated from a dose–response curve for which the viral yields obtained in IFN-treated cells were expressed in terms of percentage inhibition with respect to the yield from virus-infected control cells. The results of the evaluation of hMPV type I IFN sensitivity are shown in Table 1. The IFNs demonstrated substantially different capacities for inhibiting the hMPV yield in Hep-2 cells. Specifically, in Hep-2 cells pre-treated with IFNs and infected with hMPV (MOI = 0.01 TCID50/cell) for 24 h, leukocyte IFN-α was less potent than IFN-β, with 45 IU/mL of the latter inhibiting 50% of hMPV replication. Higher concentrations (19.5-fold) of leukocyte IFN-α were required to inhibit the virus yield to the same extent. Thus, one can speculate that some IFN-α subtypes present in the mixture represented by leukocyte IFN-α may be more active than others.
hMPV (MOI = 0.1) versus hMPV (MOI = 0.01): p < 0.05, with the exception of IFN-α2, 14, and 21, by Student's t-test.
hMPV (MOI = 0.1) versus VSV (MOI = 0.1): p < 0.05 by Student's t-test.
The analyses were performed using SPSS v.13.0 for Windows (SPSS Inc., Chicago, IL).
The data, expressed in terms of 50% inhibitory concentrations (IC50) are the means ± standard deviations of three separate experiments.
MOI, multiplicity of infection; IU, international unit.
The antiviral actions of the different subtypes of leukocyte IFN-α, namely IFN-α1, 2, 5, 6, 7, 8, 10, 14, 17, and 21, against hMPV were then considered. The results indicate that all the IFN-α subtypes investigated were active against hMPV replication in Hep-2 cells to differing degrees (Table 1). Four subtypes appeared to have the greatest activity against hMPV: IFN-α5, 6, 8, and 10 (IC50 ≤88 IU/mL). Other subtypes were intermediately active (IFN-α1, 2, 7, and 14: IC50 range 105–820 IU/mL), or had only modest effects (IFN-α17 and 21: IC50 >1000 IU/mL) on hMPV replication. The effect of IFN concentration on hMPV yield at a 10-fold higher MOI was also evaluated. The results again indicated that hMPV is more sensitive to the action of IFN-β than leukocyte IFN-α. Similar results, with some minor differences, were also obtained using IFN-α subtypes. Values of IC50 ranged from 91 IU/mL (IFN-α5) to 15,000 IU/mL (IFN-α17). All findings were relatively similar when the specific activities of the different IFN preparations were considered (i.e., when the results were expressed in terms of IC50 in ng/mL; Table 1).
In order to validate these results and to characterize the human Hep-2 cell line from the point of view of IFN sensitivity, experiments designed to assess the antiviral effects of the same IFN preparations on VSV replication were performed on the same cells, as previously reported (22). As expected (Table 1), the results indicated that VSV is sensitive to all type I IFN preparations tested.
The results clearly indicated that IC50 values for the different type I IFN preparations against VSV were significantly lower than those obtained when Hep-2 cells were infected with hMPV at an MOI of 0.1 TCID50/cell (p < 0.05 using Student's t-test). Our results indicate that hMPV is partially resistant to the antiviral activity of IFN-β and different IFN-α subtypes, unlike a well-known IFN-sensitive virus, VSV. These results are in agreement with those of Dinwiddie and Harrod, which showed that hMPV is not sensitive to IFN-α-mediated viral clearance (6). In contrast, other studies, performed in vitro and in a mouse model, indicated that IFN-α is able to inhibit hMPV replication (2,12). The reasons for these discrepancies are unclear, but may be related to the different methods employed to determine hMPV's sensitivity to IFN, or to differences in the hMPV strain or cell type used. We also demonstrated, for the first time to our knowledge, that IFN-α subtypes have distinctive antiviral activities against hMPV, with the IFN-α5, 6, 8, and 10 subtypes being most effective, and IFN-α2, 17, and 21 being the least effective at inhibiting hMPV replication. Thus our findings may indicate an escape mechanism developed by hMPV to evade the antiviral activities of specific IFN-α subtypes. Indeed, several members of the Paramyxoviridae family have been shown to directly target STAT signaling through distinct mechanisms, including proteasomal degradation, sequestration in high-molecular-weight complexes, and inhibition of nuclear localization of STAT proteins (9,10,19). It has also recently been demonstrated that hMPV can inhibit type I IFN responses through regulation of STAT phosphorylation (6).
The differences in antiviral activity of the individual IFN-α subtypes against hMPV may correlate with their relative binding affinities for the receptors. Indeed, despite strong sequence and structural homology and a common cell surface receptor, the IFN-α subtypes differ in the extent of their antiviral responses, depending on their relative affinity for their receptor (7,14,26,28). In addition, studies comparing the differential effects of IFN subtypes on the activation of signaling molecules demonstrate differential activation of STAT, as well as mitogen-activated protein kinase (MAPK), following IFN-α subtype treatment (4). Thus it appears that there are distinct signaling pathways for the different subtypes, which may explain their differing activities.
In order to delineate a tentative characterization of the different effects of type I IFNs against hMPV in Hep-2 cell lines, we tested the expression of two well-known IFN-stimulated genes: protein kinase R (PKR) and myxovirus resistance protein A (MxA) (21).
The mRNA copy contents of their genes were measured in untreated versus IFN-treated Hep-2 cells. Briefly, Hep-2 cells (2.5 × 106) were exposed to 5 ng/mL of each type I IFN preparation for 24 h. After 24 h of type I IFN exposure, the cells were harvested and RNA was isolated. A TaqMan RT polymerase chain reaction was used to quantify PKR and MxA mRNA expression as previously described (22,23). Data, normalized to the expression of the glucuronidase gene (GUS, Assay-On-Demand, Hs99999908_mL; Applied Biosystems), were calculated using the arithmetic formula 2-ΔΔthreshold cycle(Ct), according to Applied Biosystems guidelines. In particular, the fold change in IFN-induced PKR or MxA expression compared with untreated cells was estimated using the formula, 2−(ΔΔCt), where ΔΔCt = [Ct PKR or MxA (+IFN) − Ct GUS (+IFN)] − [Ct PKR or MxA (−IFN) − Ct GUS (−IFN)].
The results showed that significant increases, relative to untreated Hep-2 cells, were observed for MxA- and PKR-mRNAs after in-vitro treatment of Hep-2 cells with all type I IFN preparations (p < 0.05 by Student's t-test; Fig. 2). However, no specific association was found between the expression of MxA or PKR mRNAs and the different antiviral activities of the type I IFNs against hMPV. Although the protein levels of MxA and PKR were not measured, and taking into consideration that in several studies researchers found a correlation between mRNA-levels and protein changes (8,11,13,15), these data suggest that these two proteins are not responsible for the differing antiviral activities displayed in these cells by the different type I IFNs.
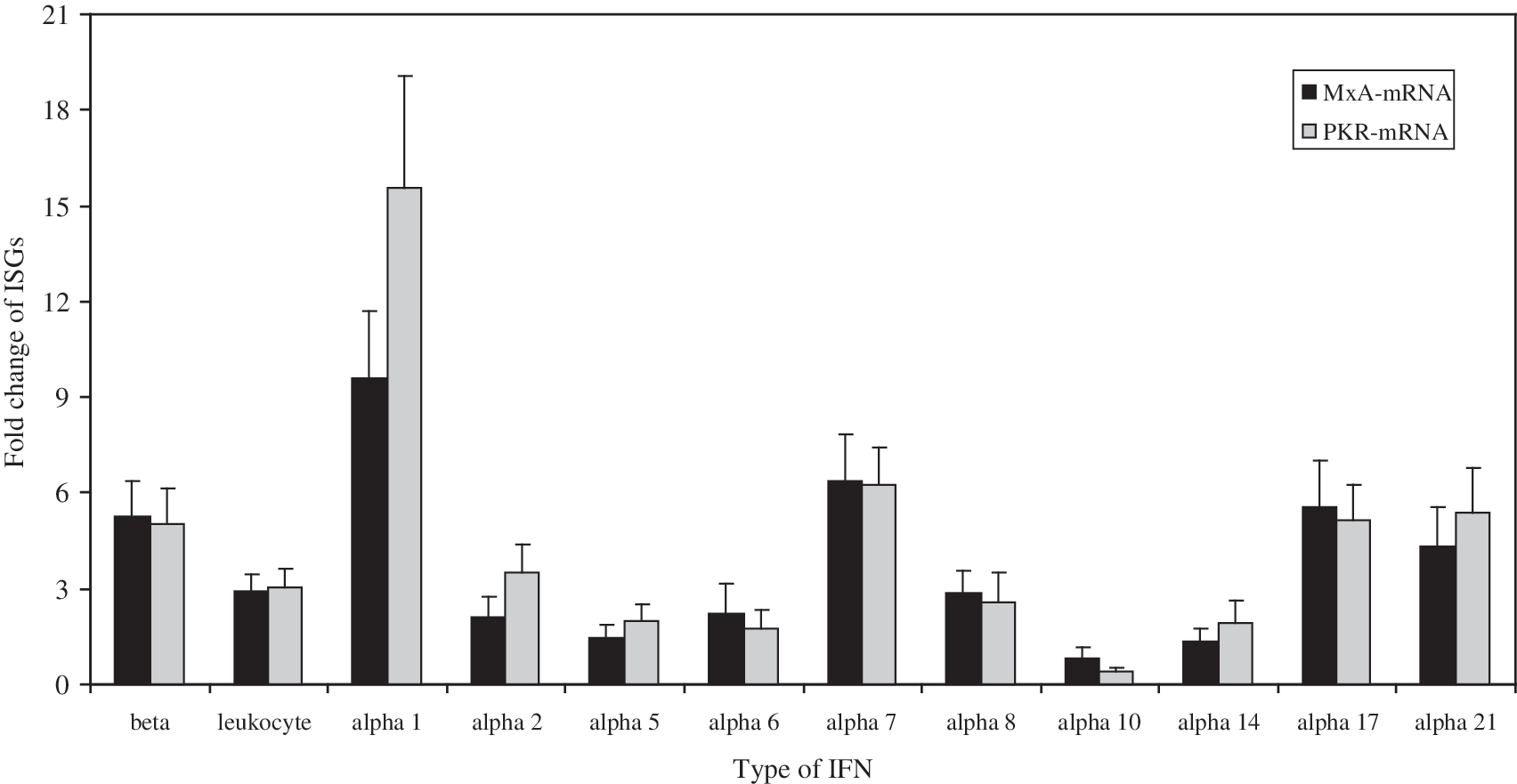
Expression of interferon (IFN)-stimulated genes (ISGs) (protein kinase R [PKR] and myxovirus resistance protein A [MxA]) in human Hep-2 cell lines treated in vitro with different type I IFN preparations. Hep-2 cells (2.5 × 106) were treated in vitro with different type I IFN preparations (5 ng/mL), and the levels of MxA- and PKR-mRNAs were measured 24 h later by TaqMan reverse transcriptase polymerase chain reaction. Data, normalized to glucuronidase gene expression, were calculated using the arithmetic formula 2-ΔΔCt, according to Applied Biosystems' guidelines. Each bar represents the mean of three separate experiments ± standard deviation. Significant increases, relative to the untreated Hep-2 cells, were observed for MxA- and PKR-mRNAs after in-vitro treatment of Hep-2 cells with all type I IFN preparations (p < 0.05 by Student's t-test).
Thus the reasons for the low and differing antiviral activities of the IFN-α subtypes against hMPV remain unclear, and this requires further investigation.
Conclusions
In conclusion, this study provides evidence that hMPV is partially resistant to the antiviral activity of IFN type I, although some IFN-α subtypes appear to be more active than others. Additional studies of how hMPV inhibits type I IFN pathways, and the consequences for the magnitude and quality of innate and acquired immunity, will be crucial in understanding the mechanisms behind hMPV pathogenesis.
Footnotes
Acknowledgments
This work was supported by grants to G.A. from the Pasteur Institute (Cenci Bolognetti Foundation; title of the project: “Molecular characterization of viruses causing bronchiolitis and study of viral and host factors affecting type I IFN antiviral response induced by respiratory viruses”), PRIN 2008 (no. 20085JWPK3), and to A.P. from “Sapienza” Università di Roma (Fondi ricerche Universitarie), and the Italian Ministry of Health (Ricerca Finalizzata conv. no. 88).
Author Disclosure Statement
No competing financial interests exist.