
Abstract
Combining results from previous studies, a multi-epitope antigen PCXZ against the hepatitis C virus was synthesized in this study. The antigenic specificity of PCXZ was determined by recognizing antibodies in serum samples from hepatitis C virus patients, but not from healthy subjects or subjects who had the hepatitis B virus. The characteristics of PCXZ immunogenicity were evaluated in BALB/c mice. Strong antibody responses were generated in mice immunized with either naked PCXZ or PCXZ in Freund's adjuvant. As for the T-cell responses, Freund's adjuvant significantly increased interferon-γ secretion and enhanced the lytic activity of cytotoxic T lymphocytes. The epitope Pa, one component of PCXZ, made the most significant contribution to specific CTL lysis; this epitope was also a B-cell epitope and was able to induce high IgG titers. In summary, PCXZ was found to be highly immunogenic, and elicited both humoral and cellular immune responses in mice.
Introduction
Previous studies using the synthetic antigen GZ-PCX showed that this antigen possessed a promising capability to induce specific antiviral immunity in mice and rabbits (14). GZ-PCX consists of β-galactosidase, five linear epitopes derived from several HCV proteins, and the T epitope from tetanus toxoid (TT) protein. To improve the strength of the immune response against HCV, Chen et al. (6) modified the vaccine sequence by removing the gz (β-galactosidase) gene, and linking three tandem repeats of the pcx gene, to form the pcx3 gene, which codes for a linear polypeptide. The modified vaccine achieved an exciting result that, by injection the HCV-positive serum from patients which were inoculated with the mouse anti-PCX3 serum previously, three of five transgenic mice with chimeric human liver eliminated the HCV infection 2 wk later. Actually, the aforementioned vaccines induced antiviral responses mainly by humoral immunity. However, immune responses mediated by helper T cells and cytotoxic T cells are necessary for viral clearance and protective efficacy against chronic infection (32). In this study, to enhance T-cell immune responses, three other identified epitopes that are recognized by Th cells or CTLs (Table 1) were fused to the pcx gene to construct a synthetic multi-epitope gene, pcxz. After immunization with the synthetic multi-epitope antigen PCXZ, potential antiviral immune responses, including increased secretion of specific IgG against the antigen or epitope, specific lysis reactions mediated by CTLs, and increased secretion of the proinflammatory cytokine interferon-γ (IFN-γ), were determined to assess the protective effects of the synthesized multi-epitope vaccine.
Materials and Methods
Multi-epitope gene construction
On the basis of the composition of the pcx gene (6,14), three epitope genes that were derived from HCV structural and nonstructural proteins (Table 1) were selected, and their sequences were optimized for better translation in E. coli by computer software. Synthetic nucleotide sequences corresponding to the three epitope genes were synthesized by Sangon (Shanghai, China), and inserted between the Hind III and Xho I sites of the plasmid pET28b-pcx (6), to construct the multi-epitope gene pcxz (Fig. 1).

Map of the pcxz multi-epitope gene. The pcx gene was constructed by Huang et al. (14). Additional epitope genes (from Pb to Pa) were produced by Sangon. The linker is a polypeptide (glycine-proline-glycine) between two epitopes.
Each epitope peptide (Table 1) was also synthesized at a 98% purity by California Bioscience (Coachella, CA). For ELISA experiments, the peptides were cross-linked with BSA.
Protein expression and purification
The aforementioned recombinant plasmids were transformed into E. coli BL21 (DE3) cells and expressed in Terrific broth (TB) medium supplemented with 100 μg/mL kanamycin at 37°C for 8 h with 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) induction. Cells were harvested by centrifugation at 3000 rpm for 10 min at 4°C. The harvested cells were lysed by ultrasonication. The sonicated solution was centrifuged at 10,000 rpm for 30 min, and the pellet containing chimeric protein was dissolved in 8 M urea. The expressed proteins were analyzed by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
Dissolved protein was incubated with Ni2+-nitrilotriacetate (NTA)-agarose (Qiagen Inc., Valencia, CA) according to the manufacturer's instructions. The purified protein in 8 M urea was concentrated with PEG 20,000 and further dialyzed in a urea gradient (from 8 M to 0 M) in TGE solution (50 mM Tris [pH 7.9], 0.5 mM EDTA, 50 mM NaCl, and 5% glycerol), and finally in 10 mM phosphate-buffered saline (PBS) (pH 7.4) for desalination and renaturation. The concentrations of the expressed proteins were determined by Lowry's method (30).
Antigenicity identification
Purified PCXZ antigen was incubated with human serum samples from hepatitis C patients. A total of 30 HCV-positive, 7 HBV-positive HCV-negative, and 22 healthy subject serum samples were collected in Shanghai, China, and screened for the presence of antibodies to PCXZ. Sera were diluted 1:100 and incubated with PCXZ in coated plates, followed by incubation with a HRP-conjugated anti-human IgG antibody (Bethyl Laboratories, Montgomery, TX) as a secondary antibody, and o-phenylenediamine (OPD) as an enzyme substrate. A positive result was defined as an absorbance value greater than twice the absorbance of the negative control, with a cut-off value of 0.2.
Western blotting was also performed to further identify the antigenicity of PCXZ. Purified PCXZ protein was separated by SDS-PAGE using 12% polyacrylamide gels and transferred to nitrocellulose membranes. After incubation in 5% non-fat milk in TBS containing 1 g/L Tween-20 for 1 h at room temperature, the membrane was incubated with human anti-HCV-positive or -negative sera that was diluted 1:100. HRP-goat anti-human IgG was used as a secondary antibody.
Antigenic immunization
Male BALB/c mice that were 6–8 wk old were purchased from Shanghai Laboratory Animal Center (Shanghai, China), and were randomized into three groups, with each group containing eight mice. At week 0, the animals were subcutaneously injected with 50 μg of purified antigen, either emulsified in Freund's complete adjuvant (Sigma-Aldrich, St. Louis, MO), or untreated. At weeks 3 and 5, the same antigens alone or with Freund's incomplete adjuvant were delivered, respectively, by IP injection. For the last immunization at week 7, 10 μg of the corresponding antigen, dissolved in endotoxin-free PBS, was administered via tail vein injection. Mice in the control group were administered endotoxin-free PBS with adjuvant. Blood samples were taken at week 0 (pre-immunization), and at weeks 3, 5, 7, and 9, via tail bleeds. All sera were stored at −20°C.
Enzyme-linked immunosorbent assay
To analyze the antibody levels in murine sera after immunization, 96-well Microlon plates (Grener, Germany) were coated with purified PCXZ protein at 100 ng/well in a carbonate-bicarbonate buffer (pH 9.6) overnight at 4°C. Serially diluted murine sera were added to each well and incubated for 2 h at room temperature. To determine total IgG levels, HRP-conjugated goat anti-mouse IgG antibody (Millipore, Billerica, MA) was incubated at a dilution of 1:5000 to detect bound IgG. To determine antigen-specific IgG1 and IgG2a antibody levels in murine sera, bound antibodies were incubated with HRP-conjugated goat anti-mouse IgG1 or IgG2a antibodies (Southern Biotechnology Associates, Birmingham, AL), at dilutions of 1:5000 each. The reactions were developed by adding the TMB peroxidase substrate system (KPL Inc., Gaithersburg, MD), at 100 μL/well, and the optical density was recorded at 450 nm.
To verify that the specific antibodies induced by PCXZ could be recognized by each epitope of PCXZ, synthetic peptides that were cross-linked to BSA were coated in 96-well Microlon plates with 1 μg in each well. The PCXZ and PCX (6) recombinant proteins were detected at the same time. Mouse sera were serially diluted.
Enzyme-linked immunospot assay
The ELISPOT assay was performed to determine the numbers of IFN-γ-secreting cells in the population of mouse splenocytes after vaccination (23). The assay was carried out according to the manufacturer's instructions with the mouse IFN-γ ELISPOT kit (U-CyTech Biosciences, Utrecht, The Netherlands). Two weeks after the last immunization, murine splenocytes (2 × 105 per well) were separated and cultured in triplicate wells for 40 h in Multiscreen plates (Millipore) coated with anti-mouse IFN-γ monoclonal antibody in complete RPMI 1640 culture medium (Invitrogen, Carlsbad, CA), in the presence of 10 U/mL recombinant IL-2 (PeproTech EC Ltd., London, U.K.) as negative control, 10 μg/mL PCXZ, 10 μg/mL BSA as irrelevant stimulation, 10 μM of synthetic peptide, or 5 μg/mL of concanavalin A as a positive control. The number of spots in the negative control was subtracted from the number of spots in the experimental wells containing specific proteins or peptides. Each spot represented an individual IFN-γ-producing cell. Results are shown as the mean values obtained for triplicate wells.
Cytotoxic T lymphocyte assay
CTL assays were performed based on the evaluation of cytoplasmic lactate dehydrogenase (LDH) activity following cell lysis, as previously described (17). In brief, aliquots of 5 × 106 splenocytes were stimulated with 10 μg/mL PCXZ and recombinant IL-2 (30 U/mL) in complete RPMI 1640 for approximately 7 d, and re-stimulated on day 5 using irradiated naïve splenocytes loaded with the selected antigen protein (10 μg/mL). On day 7, re-stimulated splenocytes were used as effector cells in the Cytotox 96 non-radioactive cytotoxicity assay (Promega Corp., Madison, WI) against P815 target cells, which were either untreated or loaded with 10 μg/mL of the selected protein, BSA, or 10 μM of synthetic peptide. Different effector:target cell ratios were tested. After 4 h of incubation, the activity of LDH released upon cell lysis was measured. The lysis percentage was calculated according to the following formula: [(experimental release – spontaneous release)/(maximal release – spontaneous release)] × 100. For each effector:target cell ratio, the results are shown as the mean percentage of specific lysis obtained from duplicate experiments.
Statistical analysis
Data from multiple experiments are presented as means ± standard error (SE). The Student's t-test was used to compare differences between two groups, and one-way analysis of variance (ANOVA) was used for more than two groups. p Values <0.05 were considered statistically significant. Statistical analyses were performed using SPSS 12.0 software with Windows XP (SPSS Inc., Chicago, IL).
Results
Design of the synthetic multi-epitope gene
Previously, we constructed a multi-epitope gene, pcx, that contained five linear epitopes derived from several HCV proteins: core (1 –17, 132–140), E1 (317–325), NS3 (1445–1453), and NS5B (2781–2788), and the T epitope derived from TT protein (14). In this study, three other identified epitopes (Table 1) were added to enhance T-cell immune responses. Considering the genetic variability of HCV, the selected epitopes were highly conserved (Table 2). The order of the epitopes was arranged to prevent the formation of complex secondary structures and transmembrane domains. A linker (glycine-proline-glycine) was inserted into the gap between epitopes (Fig. 1). The synthetic multi-epitope gene pcxz was cloned into the bacterial expression vector pET-28b (+), and the sequence was confirmed by DNA sequencing.
Variations are underlined.
Recombinant expression of the pcxz recombinant gene
The recombinant plasmids pET28b-pcxz were transformed into competent E. coli BL21 (DE3) cells, and protein expression was induced. The recombinant proteins were expressed in inclusion bodies that formed in the transformed cells. The expressed protein PCXZ had a molecular weight of approximately 21.1 kDa, as expected (Fig. 2).
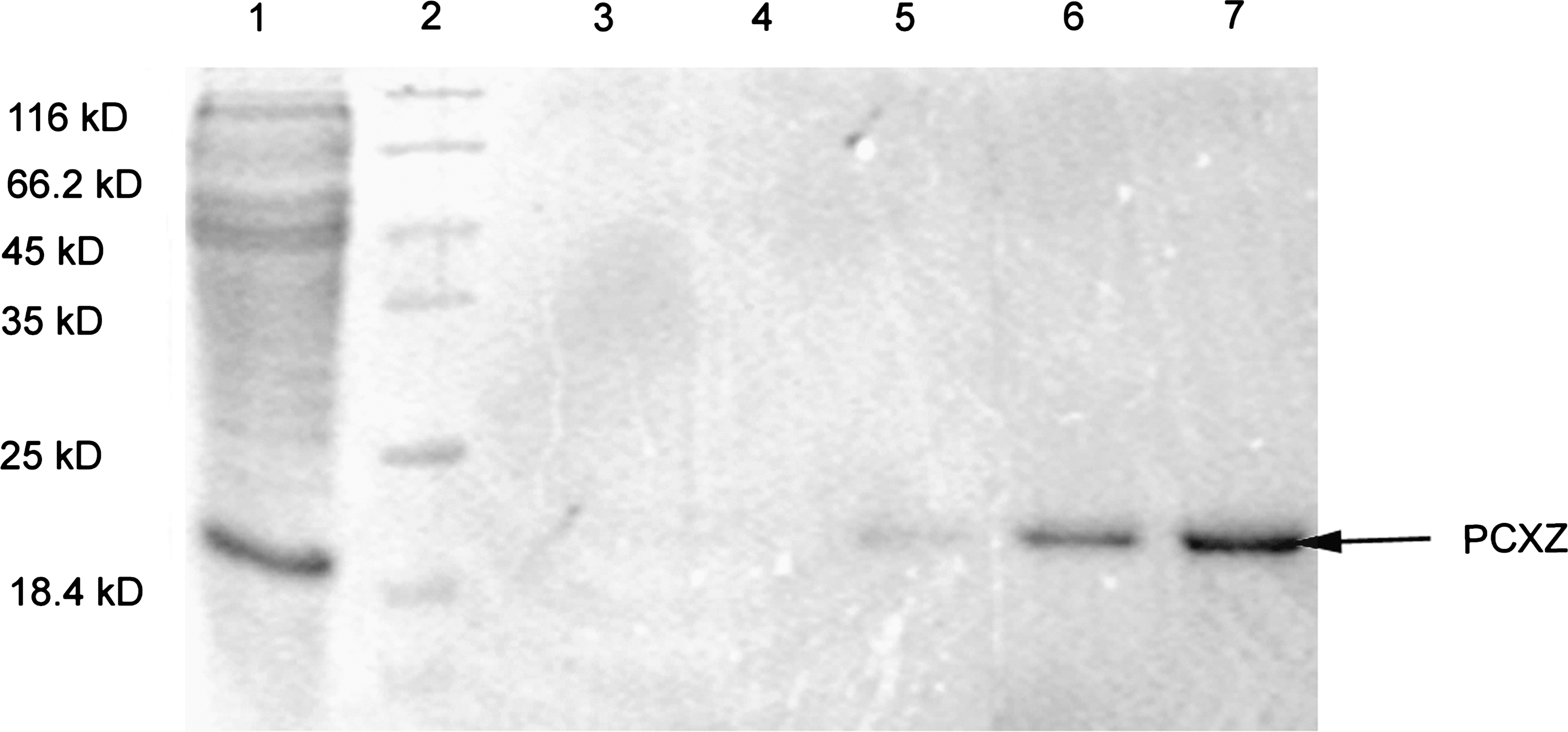
Expression and purification of the PCXZ protein in E. coli. High-level expression of PCXZ (approximately 21.1 kDa) was induced with 1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) in E. coli BL21 (DE3) cells. Purification of PCXZ was performed under denaturing conditions (lane 1, cells induced with IPTG; lane 2, protein molecular weight markers; lanes 3–5, flow-through; lane 6, eluate; lane 7, purified PCXZ).
Specific antigenicity of the PCXZ recombinant protein
The immunological reactivities of the synthetic multi-epitope antigens were detected by ELISA and Western blot. Using the synthetic proteins as coating antigens, 30 serum samples collected from hepatitis C patients, 7 serum samples collected from hepatitis B patients (anti-HBs-positive), and 22 serum samples from normal healthy subjects were examined by ELISA. The results showed that the synthetic proteins carrying the HCV epitopes specifically reacted with anti-HCV antibodies (Fig. 3). In total, 25 of the 30 tested serum samples from HCV patients reacted with the PCXZ protein, and only 1 of the 22 serum samples from normal healthy subjects showed slight detectable immunoreactivity with PCXZ.
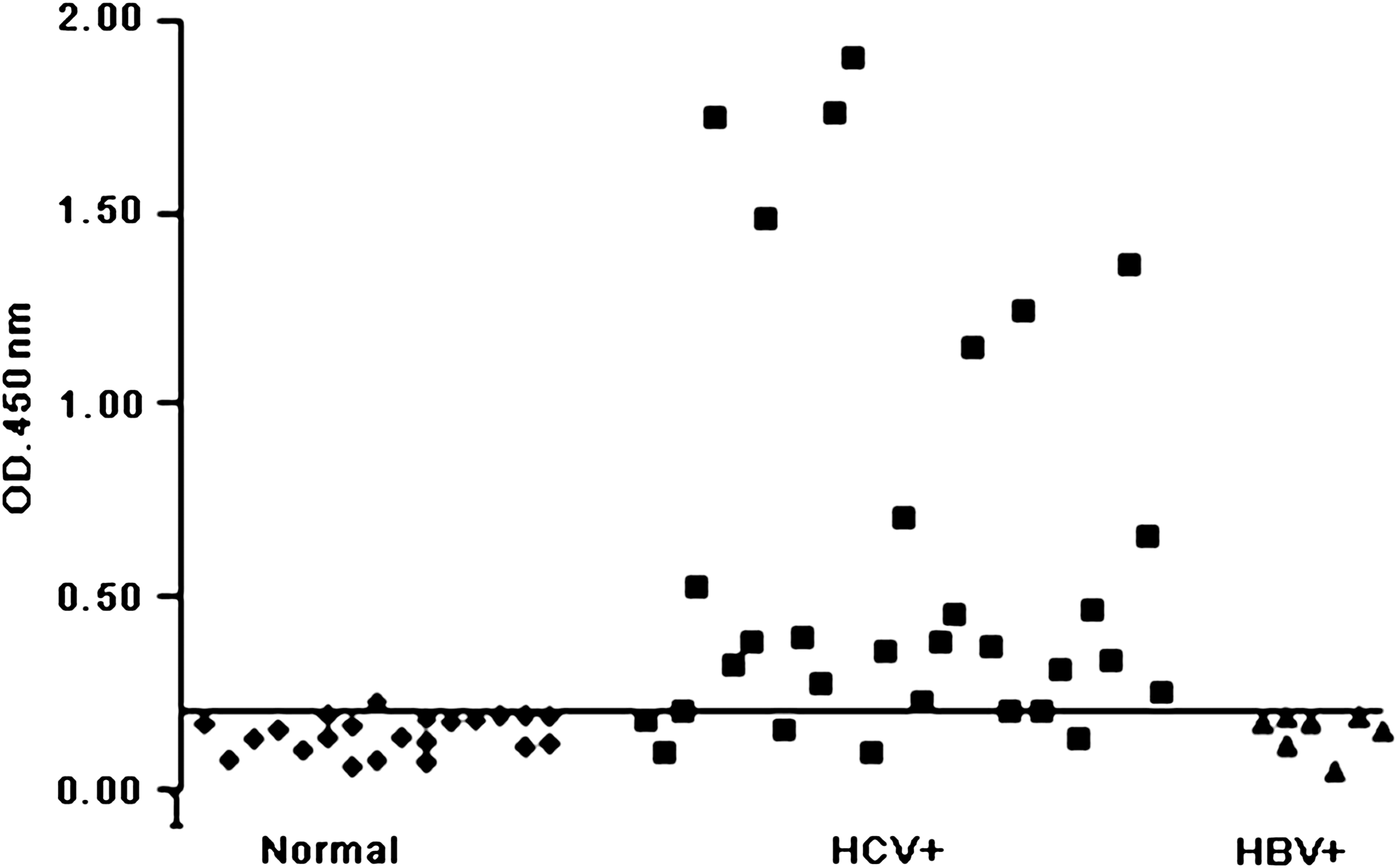
Reactions of the synthetic multi-epitope antigen as a coating antigen with anti-HCV antibodies in sera from hepatitis C patients (HCV+), hepatitis B patients (HBV+), or normal healthy subjects, by ELISA. A positive result was defined as an absorbance value greater than twice the absorbance of the negative control, with a cut-off value of 0.20.
Western blot test results showed that the synthetic proteins that carried HCV epitopes could be specifically recognized by an anti-HCV antibody in serum samples from hepatitis C patients, but not in serum samples from healthy subjects (Fig. 4).
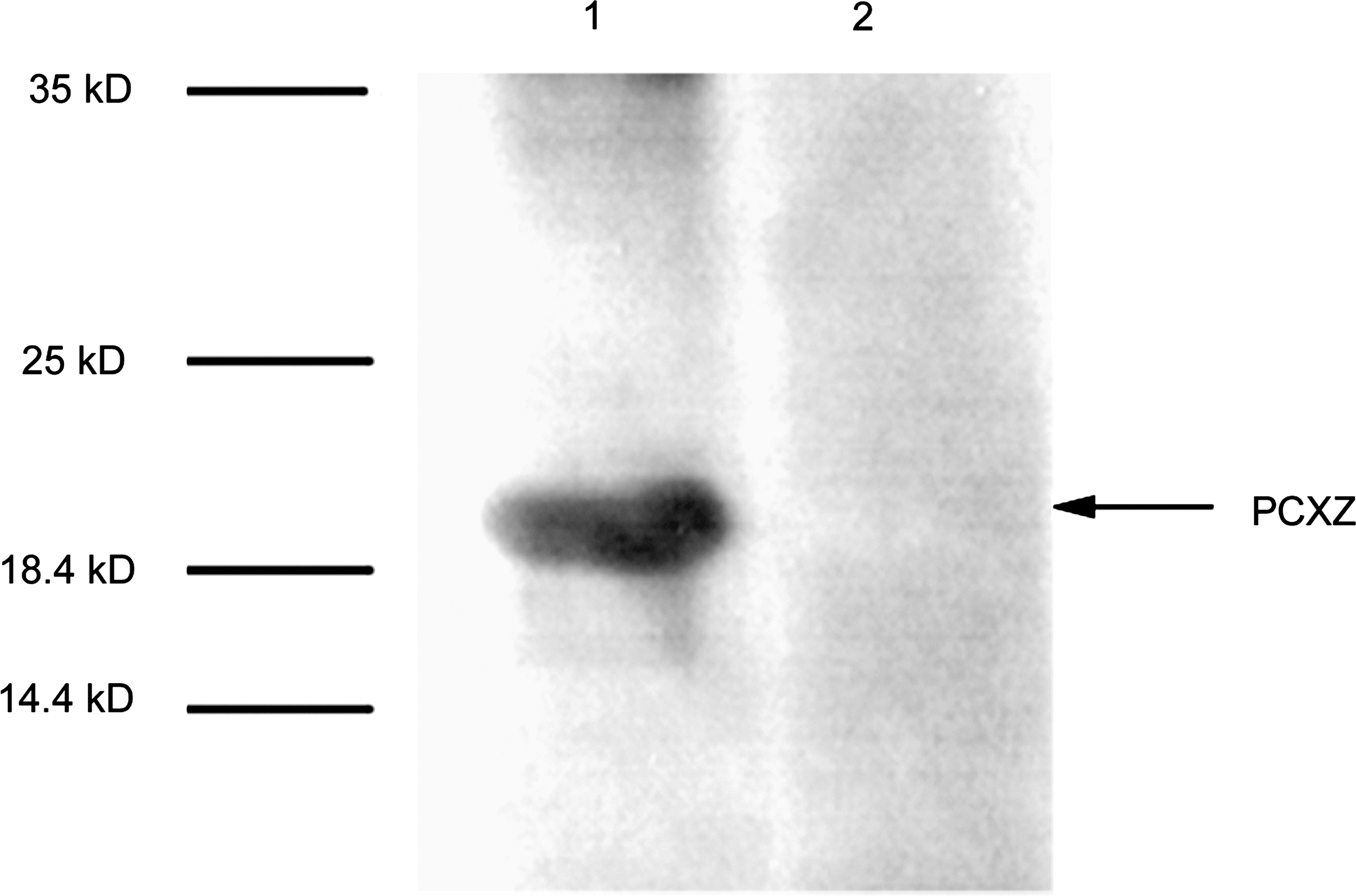
Detection of synthetic multi-epitope proteins with antibodies in serum from a hepatitis C patient or healthy control by Western blot (lane 1, serum from a hepatitis C patient; lane 2, serum from a healthy control).
Antibody responses to the synthetic multi-epitope vaccine in mice
To determine whether the synthetic multi-epitope vaccine PCXZ elicited antibody responses in mice, mouse sera collected after vaccination were analyzed for antigen-specific IgG titers by ELISA assays. As shown in Fig. 5, from the fifth week on, all mice vaccinated with PCXZ developed PCXZ-specific IgG antibodies, with a significant difference compared to the control group that was immunized with PBS and adjuvant. The antibody IgG titers of the mice of group B, which was immunized with PCXZ in the absence of adjuvant, rapidly increased at week 5 compared to those of group A, which were immunized with PCXZ and Freund's adjuvant (*p < 0.05). At weeks 7 and 9, however, there was no significant difference between the two groups. The average maximum IgG titer was approximately 6.56 (±1.25) × 105 and 2.81 (±1.25) × 105 for groups A and B, respectively, at week 9.
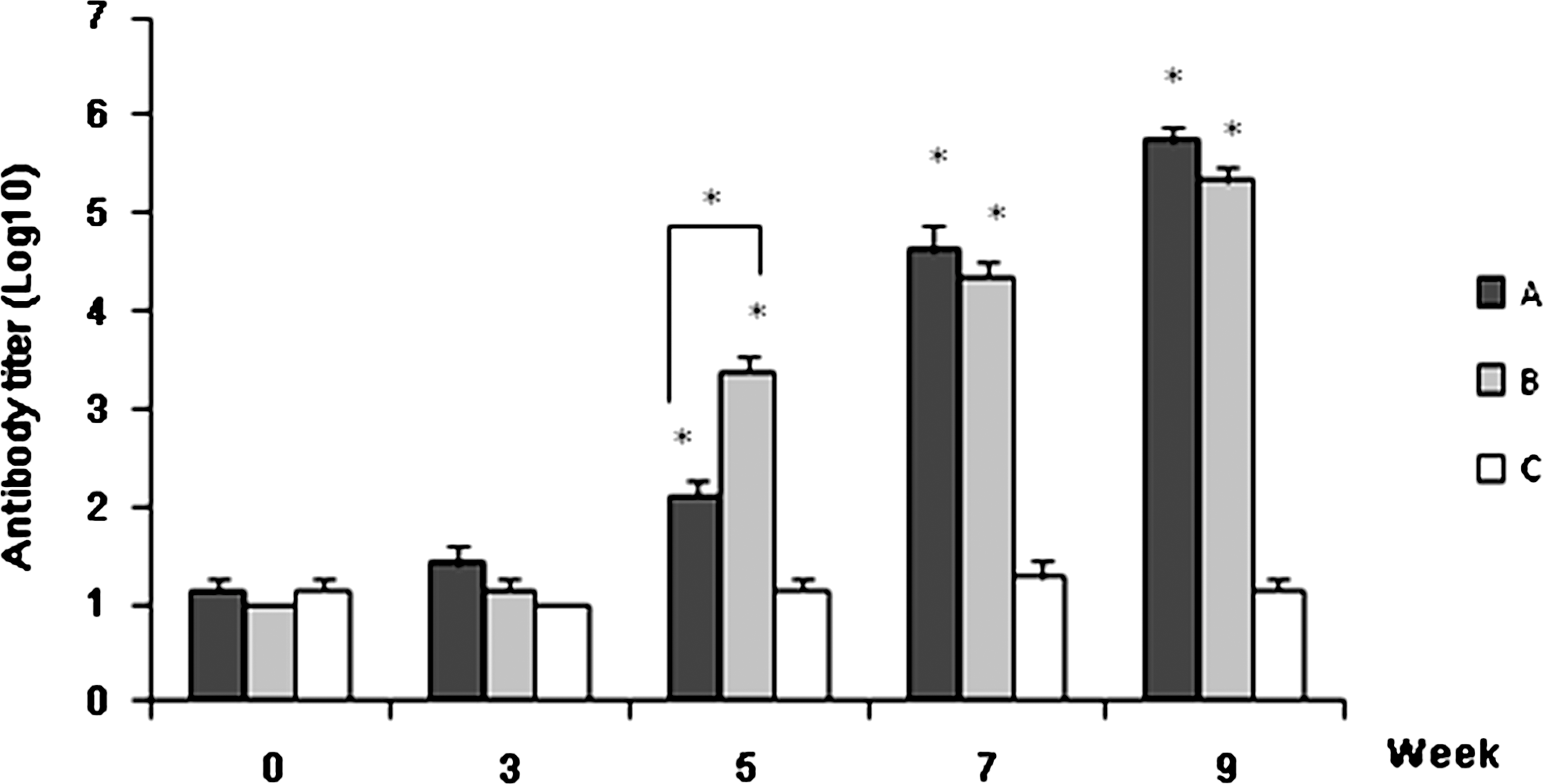
ELISA titers of antibodies against sera of mice immunized with the multi-epitope antigen PCXZ. Antibody responses were measured at several time points. From week 5, compared with the control group (group C), the differences were statistically significant for groups A and B. There was no significant difference between groups A and B except at week 5 (A, mice immunized with PCXZ emulsified in Freund's adjuvant; B, mice immunized with PCXZ; C, mice immunized with PBS and adjuvant). Similar results were obtained in three independent experiments. (*p < 0.05).
Individual epitopes in the multi-epitope antigen showed different reactivities in serum samples from vaccinated mice (Fig. 6). In sera from groups A and B, antibody titers against epitope Pa showed a significant difference compared to those against the irrelevant protein BSA. However, epitope Pb could not effectively induce a B-cell reaction. Thus, peptide Pa appeared to have an enhanced ability to elicit a B-cell reaction compared to epitopes Pb and Pc.

Synthetic peptides reacted with mouse anti-PCXZ sera. The peptides Pa, Pb, and Pc, cross-linked to BSA, were coated separately at concentrations of 1 μg/well, and the recombinant proteins PCXZ and PCX (ref. 6) were coated at concentrations of 0.1 μg/well. In addition, the irrelevant protein BSA was also coated as a control. Sera from immunized mice were collected at week 9. In sera from groups A and B, antibody titers against epitope Pa showed a significant difference compared to those against the irrelevant protein BSA (group A, mice immunized with PCXZ emulsified in Freund's adjuvant; group B, mice immunized with PCXZ; group C, mice immunized with PBS and adjuvant). Experiments were repeated three times, and similar results were obtained for each experiment (*p < 0.05 versus BSA).
As an indicator for evaluating the quality of the immune response (1,21), the relative levels of IgG subclasses (IgG1 and IgG2a) were determined (Fig. 7). The multi-epitope antigen induced IgG1 to a greater extent than IgG2a, and the ratios of IgG2a to IgG1 for groups A and B were 0.05 and 0.01, respectively, which indicated T-helper-2 (Th-2)-based responses.
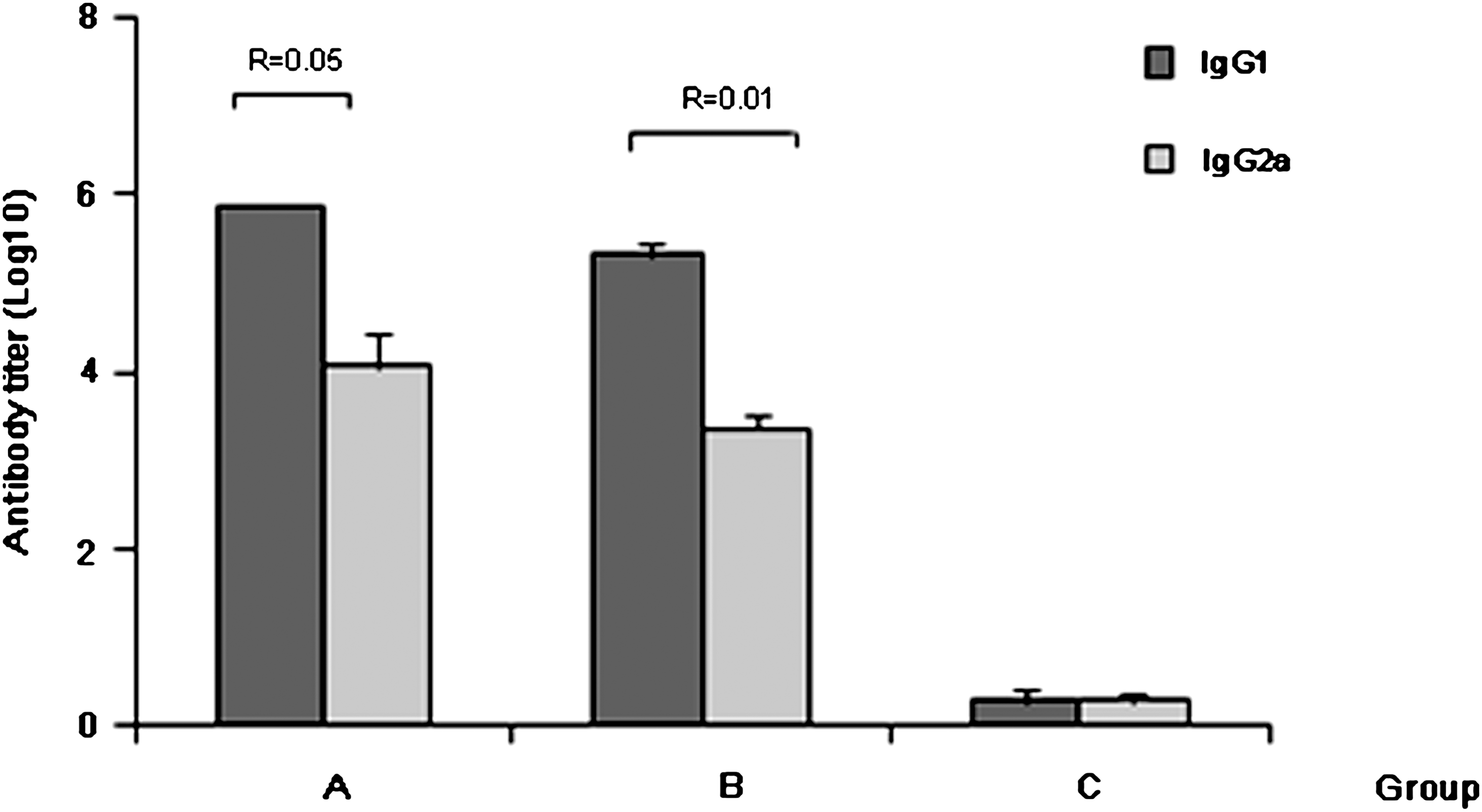
Antibody levels of IgG subclasses (IgG1 and IgG2a) in sera of mice immunized with PCXZ. Sera were collected at week 9 (group A, mice immunized with PCXZ emulsified in Freund's adjuvant; group B, mice immunized with PCXZ; group C, mice immunized with PBS and adjuvant; R, ratio of IgG2a to IgG1).
Cell-mediated responses to the synthetic multi-epitope vaccine in mice
To determine whether the synthetic multi-epitope vaccine was able to induce cell-mediated immune responses in mice, after vaccination, mouse splenocytes were separated, and IFN-γ secretion levels upon antigen re-stimulation in vitro were analyzed. The frequencies of IFN-γ-producing T cells targeting the synthetic multi-epitope protein PCXZ were 608 ± 77 and 166 ± 50 spots per 106 cells for the PCXZ-vaccinated groups A and B, respectively. Freund's adjuvant significantly enhanced the specific secretion of IFN-γ, as shown by comparing the results of group A with those of group B. As a fragment of the full synthetic multi-epitope protein PCXZ, the synthetic protein PCX also induced the proliferation of specific IFN-γ-producing T cells, with 128 ± 32 spots per 106 cells in mice from group A (Fig. 8).

Induction of specific IFN-γ-producing T cells from mice immunized with PCXZ. Two weeks after the last immunization, splenocytes were separated and cultured for 40 h in the presence of PCXZ or PCX or the irrelevant protein BSA. The number of IFN-γ-producing T cells specific to each epitope or protein was assessed by ELISPOT assay, as described in the materials and methods section (A, mice immunized with PCXZ emulsified in Freund's adjuvant; B, mice immunized with PCXZ; C, mice immunized with PBS and adjuvant). Similar results were obtained in three independent experiments (*p < 0.05).
Because CTLs are one of the major effectors of cell-mediated immune responses, we also determined the presence of specific CTLs against PCXZ/PCX and each individual epitope after vaccination. Mice immunized with PCXZ had induced cytolytic responses that were specific to PCXZ (25–41% for group A and 27–35% for group B), with the corresponding effector-to-target ratios (E:T) ranging from 6.25:1 to 25:1 (Fig. 9a). Comparison of the targeting antigens PCXZ and PCX revealed that a significantly lower level of cytolysis was found for PCX (Fig. 9b). For peptide Pa, specific CTLs displaying a high percentage of lysis were induced 40% and 34% for groups A and B, respectively, at an E:T ratio of 25:1 (Fig. 9d). For peptide Pc, only weak cytolytic activity was detected for groups A and B (25% and 24%, respectively), with an E:T ratio of 25:1 (Fig. 9f). For peptide Pb, lytic activity was nearly undetectable (Fig. 9e). No specific lysis was detected with the irrelevant protein BSA (Fig. 9c).
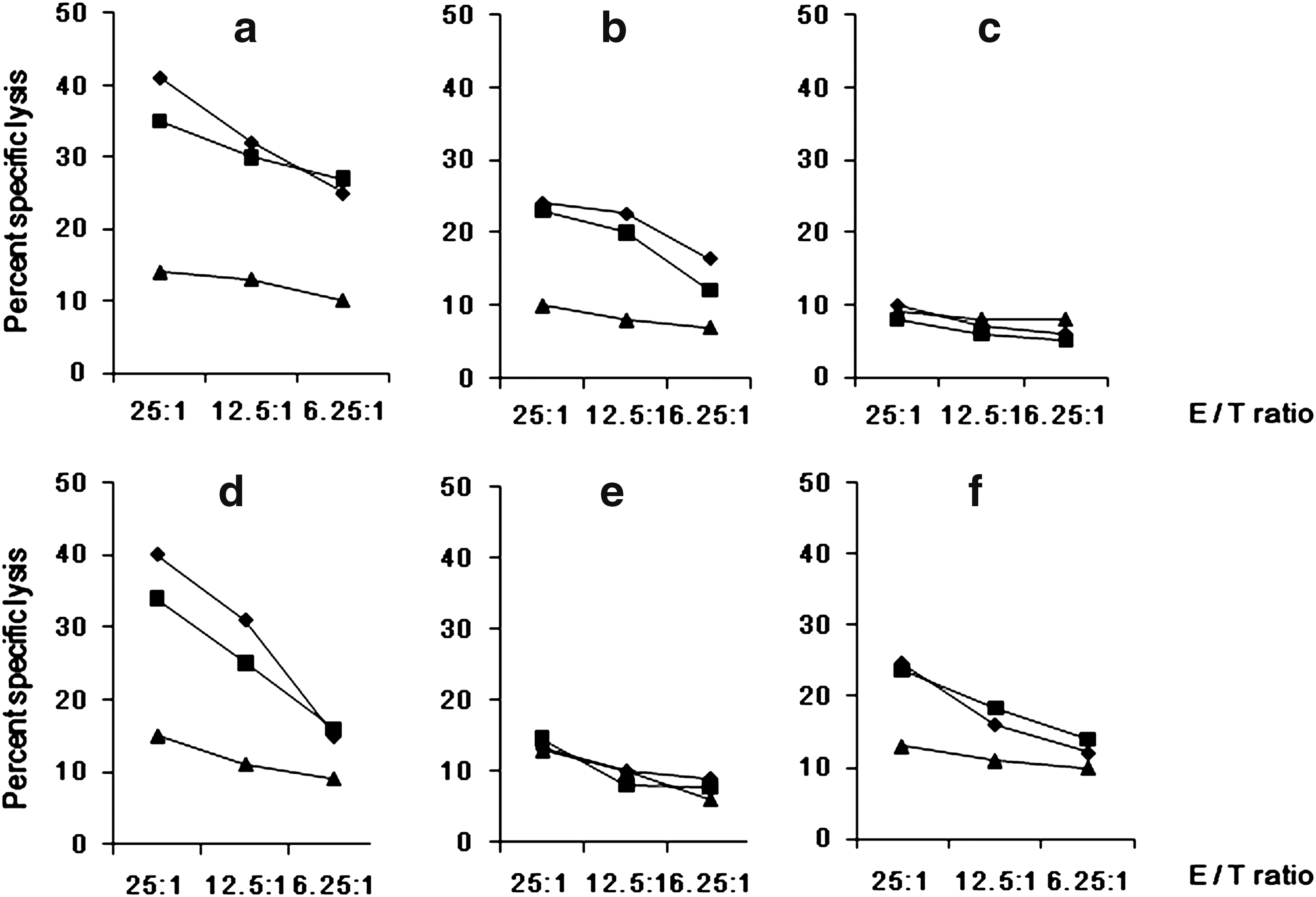
CTL responses of mice immunized with the PCXZ antigen against a synthetic epitope peptide, PCXZ or PCX. Induced cytolytic responses specifically targeting PCXZ (
Discussion
On the basis of synthetic multi-epitope vaccines using the GZ-PCX (14) or PCX3 (6) antigens, three additional linear epitopes contained in HCV, E2, NS3, and NS5B, were added to construct the synthetic multi-epitope vaccine PCXZ. In BALB/c mice, immunization with the PCXZ vaccine induced the production of high-titer IgG, and induced lytic activity of cytotoxic CD8+ T cells and IFN-γ secretion by CD4+ T cells. The immune response was mainly a Th-2-based response. One of the three added linear epitopes, Pa, appeared to be competent to induce significant humoral and cellular immune responses.
The antigenic specificity of a synthetic multi-epitope antigen has been the most important issue in vaccine development (29). In this study, the synthetic multi-epitope antigen PCXZ was shown to be specifically recognized by sera collected from HCV patients, but not by sera collected from healthy subjects (Fig. 4). The existence of rapid and frequent mutations in HCV has frustrated clinical diagnosis and vaccine design. Because of this recognized problem, the multi-epitope antigen PCXZ was based on conserved epitopes (Table 2). To demonstrate specific identification of HCV, serum samples from 30 HCV-positive patients were screened. The results showed that 25 out of 30 tested serum samples from HCV patients presented antigen-antibody reactions with the recombinant PCXZ protein (Fig. 3), which indicates that PCXZ epitopes can be recognized by antibodies of different HCV-infected individuals in China.
The synthetic multi-epitope vaccine PCXZ could specifically induce strong B-cell responses, and significant CD4+ and CD8+ T-cell responses in BALB/c mice. Previous studies have indicated that the synthetic multi-epitope antigens GZ-PCX (14) and PCX3 (6) could evoke strong humoral immune responses, and sera from mice immunized with PCX3 were able to diminish and even eliminate viral loads in transgenic mice with chimeric human livers. However, based on current knowledge of the mechanisms involved in the control of viral infections, a vaccine should induce a multi-specific and vigorous cellular host response, involving both CD4+ and CD8+ T cells, and a strong and cross-neutralizing antibody response. There is little evidence that any vaccine achieves protection through antibodies alone or prevents initial infections entirely; the same holds true for T-cell responses (12,15,20,32,37). An important potential advantage of multi-epitope–based vaccines is the elicitation of a number of different responses simultaneously (29). Kaba et al. (18) developed a multi-stage multi-valent antigen, FALVAC-1A, which consists of 21 different B-cell, T-cell, and CTL epitopes from seven different antigens of P. falciparum, and demonstrated that the synthetic antigen was a highly immunogenic vaccine candidate antigen, that was capable of eliciting humoral and cellular responses in diverse genetic backgrounds. In this study, T-cell responses were enhanced because additional T-cell-specific epitopes were added to the multi-epitope antigen pcx gene. After immunization in mice with the synthetic antigen PCXZ, for example, the frequencies of IFN-γ-producing CD4+ T cells increased from 128 ± 32 to 608 ± 77 per 106 splenocytes when stimulated by PCX or PCXZ in vitro, respectively (Fig. 8). CTL lysis of CD8+ T cells increased from 23% (for targeting PCX) to 41% (for targeting PCXZ; Fig. 9a and 9b).
The epitope Pa, one component of PCXZ, which corresponds to NS31248–1261, appeared to be competent to induce significant humoral and cellular immune responses, and seemed to be a promising epitope for multi-epitope or subunit vaccine development. The nonstructural protein 3 (NS3) was among the HCV antigens so far described that contains both CD4+- and CD8+-restricted epitopes (4), and it seemed to emerge as a mandatory immunogen for T-cell responses (32). In this study, the epitope Pa (NS31248–1261), which was demonstrated to dominate the immune response with spontaneous viral clearance (8), was also selected. After immunization with multi-epitope antigen PCXZ in BALB/c mice, our results showed that both B-cell responses and cytolytic CD8+ T-cell responses were induced targeting Pa epitope. CD4+-derived IFN-γ-producing T cells were undetectable using the ELISPOT technique. It is possible, however, that other cytokines might have been induced (e.g., a Th-2 type of cytokine). Our results are consistent with observations made by Brinster et al. (3). Several sets of researchers (9,13,35) found that regions of E2 were involved in the CD81 interaction. Owsianka et al.(24) demonstrated that the mAb3/11-targeting epitope E2412–423 could inhibit binding to CD81 of HCV-like particles, indicating that the epitope E2412–423 is an important neutralization epitope. In this study, the corresponding epitope, Pb (epitope E2412–423), of the synthetic multi-epitope antigen PCXZ did not efficiently induce B-cell responses. In addition, the epitope Pc of PCXZ, corresponding to NS5B2578–2602, is HLA-A2.1-restricted (11,31,36,38), which might be the reason why the immune responses targeting this epitope were not sufficiently strong in BALB/c mice.
Immunological adjuvants play an important role in immune responses, especially for purified recombinant molecules or subunits of pathogen vaccines (10). Freund's adjuvant was used in this study, and the results indicated that T-cell responses, particularly the frequency of IFN-γ-producing T cells, were enhanced by the use of adjuvant; however, the ratio of IgG2a to IgG1 subclasses indicated that the immune response was mainly a Th-2-based response. Several reports have indicated that CpG ODN is an excellent immune adjuvant, switching on T-helper 1 (Th-1) immune responses (7,19,26). Jiao et al. (16) demonstrated that liposomes co-encapsulating HCV NS3 and CpG are a good candidate vaccine to induce strong Th-1 immune responses against hepatitis C viruses. Lin et al. (22) found that NS345 core polyprotein adjuvanted with Iscomatrix plus CpG was optimal for eliciting Th-1-type CD4+ T-cell responses in mice.
Footnotes
Acknowledgment
This work was support by the National High Technology Research and Development Program (“863” Program) of China (grant no. 2006AAD2A228).
Author Disclosure Statement
No competing financial interests exist.