
Abstract
Expression of major histocompatibility complex (MHC) class I alleles such as B*57 and B*27 are associated with slow HIV disease progression. HIV-specific immune responses in slow progressors (SP) are characterized by a poly-functional profile. We previously observed within infected subjects that HIV peptide-specific responses could differ from each other in their functional composition. We investigate here whether responses restricted by MHC class I alleles associated with slow disease progression have a more poly-functional profile than responses restricted by other alleles. We stimulated peripheral blood mononuclear cells (PBMCs) isolated from 36 chronically HIV-infected individuals with a panel of optimal peptides restricted by the HLA alleles expressed by each subject, and assessed the contribution of single IL-2-, single IFN-γ-, and IFN-γ/IL-2-secreting lymphocytes to the total response measured using a dual color ELISPOT assay. The contribution of functional subsets to responses restricted by HLA B*57/B*27 was similar in SP and progressors. For responses restricted by other MHC class I alleles, dual IFN-γ/IL-2-secreting lymphocytes contributed significantly more to the total response in SP than progressors. Within SP subjects, peptides restricted by both B*57/B*27 and other alleles stimulated responses with similar functional profiles. In progressors, peptides restricted by B*57/B*27 stimulated responses composed of a significantly greater proportion of IFN-γ/IL-2-secreting cells than peptides restricted by other alleles. Within progressors, the contribution of IFN-γ/IL-2-secreting lymphocytes was greater to epitopes restricted by protective HLA alleles compared with responses restricted by other alleles. HLA haplotypes influence the relative functional composition of HIV-specific responses.
Introduction
The effect of host genetics on HIV disease progression is demonstrated in part by the association between the expression of certain major histocompatibility complex (MHC) class I alleles with the rate of disease progression (12,15). MHC class I alleles such as B*57 and B*27 are frequently expressed in individuals exhibiting slow disease progression, whereas the expression of B*07, B*08, and B*35 are associated with rapid rates of progression (11,22,28,33,35). Additionally, homozygosity at the MHC class I loci confers an unfavorable prognosis, potentially caused by a narrower spectrum of peptides presented to the immune system (11,49). The effect exerted by MHC class I (or HLA) alleles on disease progression supports a protective role for the HIV-specific CD8 arm of the immune response, although the functional mechanisms involved in slowing disease progression are still under investigation.
Both quantitative and qualitative aspects of the HIV-specific immune response have been comprehensively investigated in infected individuals, and parameters such as breadth, magnitude, specificity, and function for both HIV-specific CD4+ and CD8+ T lymphocytes have provided substantial insight about what constitutes protection. Several cohort studies have found that neither breadth nor magnitude of HIV-specific interferon-γ (IFN-γ)-secreting immune responses associate with disease progression parameters (1,14,16,29,43). However, recent reports suggest a role for specificity in controlling viral replication, and conclude that HIV-specific responses targeting Gag p24 determinants are associated with control of viral replication, and should therefore be considered in vaccination strategies (9,18,24,30,34,39,45,47,48). The functional profile of HIV-specific CD4+ and CD8+ lymphocytes are also associated with different HIV disease outcomes (5,20,42). Maintenance of a poly-functional repertoire is linked with protection from disease progression, and functions such as proliferation, secretion of interleukin-2 (IL-2), tumor necrosis factor-α (TNF-α), and perforin are preserved in individuals who control viral replication (21,23,32,36,37,41,44,52,53). In contrast, individuals who typically progress to AIDS have a functionally skewed population of HIV-specific cells in which only IFN-γ secretion persists.
Although clear genotypic and phenotypic evidence supports a protective role played by HIV-specific CD8 lymphocytes, it is less clear whether there is an association between the expression of protective alleles and poly-functional immune responses. We therefore sought to determine whether responses restricted to HLA-B*57 and/or B*27 preferentially stimulate and preserve IL-2 secretion compared to responses restricted to non-B*57 and B*27 alleles. We reasoned that if poly-functional responses exerted more effective control than mono-functional responses, it would be expected that responses restricted by alleles associated with slow progression would be more likely than responses restricted by alleles associated with neutral or rapid disease progression to be poly-functional. We addressed this question by analyzing the contribution of three functionally distinct subsets of HIV-specific lymphocytes (single IFN-γ-, single IL-2-, and dual IFN-γ/IL-2-secreting lymphocytes) to the total measured response in slow progressors (SP) and progressors expressing HLA-B*57, B*27, and other alleles following stimulation of peripheral blood mononuclear cells (PBMCs) with MHC class I restricted optimal epitopes.
Materials and Methods
Study population
The study population included 36 treatment-naïve HIV-infected subjects. Twenty three were SP from the Montreal SP cohort, and 13 were progressors from the Montreal Primary Infection (PI) cohort, and from the Immunodeficiency Treatment Centre of the McGill University Health Center. For participants in the Montreal PI cohort, samples from time points corresponding to the chronic phase of infection were analyzed. The median (range) time from infection for the progressor group was 20.4 (range 8.3–55.9) mo. The SP group all had CD4+ T-cell counts above 500 cells/mm3 and were infected at least 7 years (10). The median (range) CD4+ T-cell count and viral load (VL) for the SP group was 760 (520–2300) cells/mm3, and 2.68 (1.7–4.94) log10 HIV RNA copies/mL, respectively (Table 1). Progressors all had CD4+ T-cell counts <500 cells/mm3 at the time of ELISPOT evaluation. The median CD4+ T-cell count and VL for the progressor group was 388 (150–493) cells/mm3 and 4.74 (2.67–5.51) log10 HIV RNA copies/mL, respectively (Table 1). Classification criteria for the SP population were based on the observation that in untreated HIV infection, the median time from infection for CD4+ T-cell counts to fall below 500 cells/mm3 was 1.7 y (95% confidence intervals [CI] 1.29,1.89 y) (25). Fourteen of 23 (60.9%) and 8 of 13 (61.5%) SP and progressors expressed either HLA-B*57 and/or B*27, respectively (Table 1). As expected, CD4 counts were significantly higher and VL were lower in SP versus progressors (p < 0.0005 by unpaired t-test). Informed consent was obtained from all study subjects, and the research conformed to ethical guidelines of all participating institutions.
HLA, human leukocyte antigen.
Laboratory testing
Plasma viremia was measured using the Versant HIV-1 3.0 RNA assay (bDNA) (Bayer Diagnostics, Tarrytown, NY), with a detection limit of 50 HIV-1 RNA copies/mL of plasma.
Cells
Blood was obtained by either venipuncture into tubes containing EDTA anticoagulant, or leukapheresis as previously reported (6). PBMCs were isolated by density gradient centrifugation (Ficoll-Paque; Pharmacia, Uppsala, Sweden), and cryopreserved in 10% dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO) with 90% fetal bovine serum (FBS; Medicorps, Montreal, Quebec, Canada).
HLA genotyping
Subjects were typed for MHC class I antigen expression using the line probe assay (Innogenetics Inc., Alpharetta, GA). DNA for molecular HLA typing was prepared from either PBMCs or Epstein-Barr virus (EBV)-transformed B-cell lines using the QIAamp DNA blood kit (Qiagen Inc., Mississauga, Ontario, Canada).
Dual-color ELISPOT assay for single IFN-γ, single IL-2, and dual cytokine secretion
IFN-γ and IL-2 secretion by HIV-specific cells was quantified using the dual-color ELISPOT assay as previously described (7,44). PBMCs (150,000– 200,000) were plated and stimulated for 28 h and at 37°C with a panel of HIV clade B optimal epitopes restricted by the MHC class I alleles expressed by the individual being tested at a concentration of 4 μg/mL. The stimuli were drawn from 50 previously described optimal epitopes (Table 2) (8). Cytomegalovirus (CMV), EBV, and influenza (INF)-optimal epitopes restricted to common HLA alleles were also used as positive control peptide stimuli. Media alone was used as a negative control, and anti-CD3 antibody (clone CLB-T3/4.E; Research Diagnostics Inc., Flanders, NJ) was used as a positive control stimulus.
HLA, human leukocyte antigen.
The criteria used to identify positive single IFN-γ, single IL-2, and dual-cytokine-secreting responses were obtained by testing 8 low-risk HIV-uninfected individuals for reactivity to pools of HIV peptides covering the entire genome. Uninfected individuals produced a mean of 4.2 ± 10.6 IFN-γ SFC/106 PBMCs to the stimulatory peptide pools. A positive response was defined as being 3 standard deviations above that seen for uninfected subjects (i.e., >36 SFCs/106 PBMCs, and at least threefold greater than the autologous negative control wells. Using the same rationale, the thresholds for IL-2 and dual cytokine secretion were >32 SFC/106 PBMCs, and >21 SFC/106 PBMCs, respectively, and at least threefold greater than the autologous negative control wells. The relative contribution of each functional subset was determined by dividing the magnitude of each subset's response by the total response magnitude.
Statistical analysis
Statistical analysis and graphical presentation were performed using InStat 3.05 and Prism 4.0 (GraphPad Software, San Diego, CA). Unpaired t-tests and Mann-Whitney U tests were used to assess differences in the contribution of the three functional subsets measured in SP and progressors based on the distribution of the studied variable. A p value <0.05 was considered significant.
Results
Epitopes restricted by protective MHC class I alleles stimulate dual-cytokine-secreting lymphocytes in both SP and progressors
PBMCs isolated from 36 chronically infected individuals were stimulated with a panel of optimal peptides customized for each study subject based on their HLA type. A dual color ELISPOT assay was used to detect cells secreting single IL-2, single IFN-γ, and both IFN-γ and IL-2. The percentage contribution of each of these 3 functional subsets to the total of each epitope-specific response was calculated. While expression of HLA-B*57 and B*27 alleles was associated with slow progression, some individuals expressing these alleles did exhibit HIV disease progression (38). However, not all SP express HLA-B*57 or B*27 (38). When the functional distribution of responses to epitopes restricted by HLA-B*57 or B*27 was compared between the 14 SP and 8 progressors expressing at least 1 of these alleles, the between-group proportional contribution of the three functional subsets to the total response measured was similar (Fig. 1A, B and C).
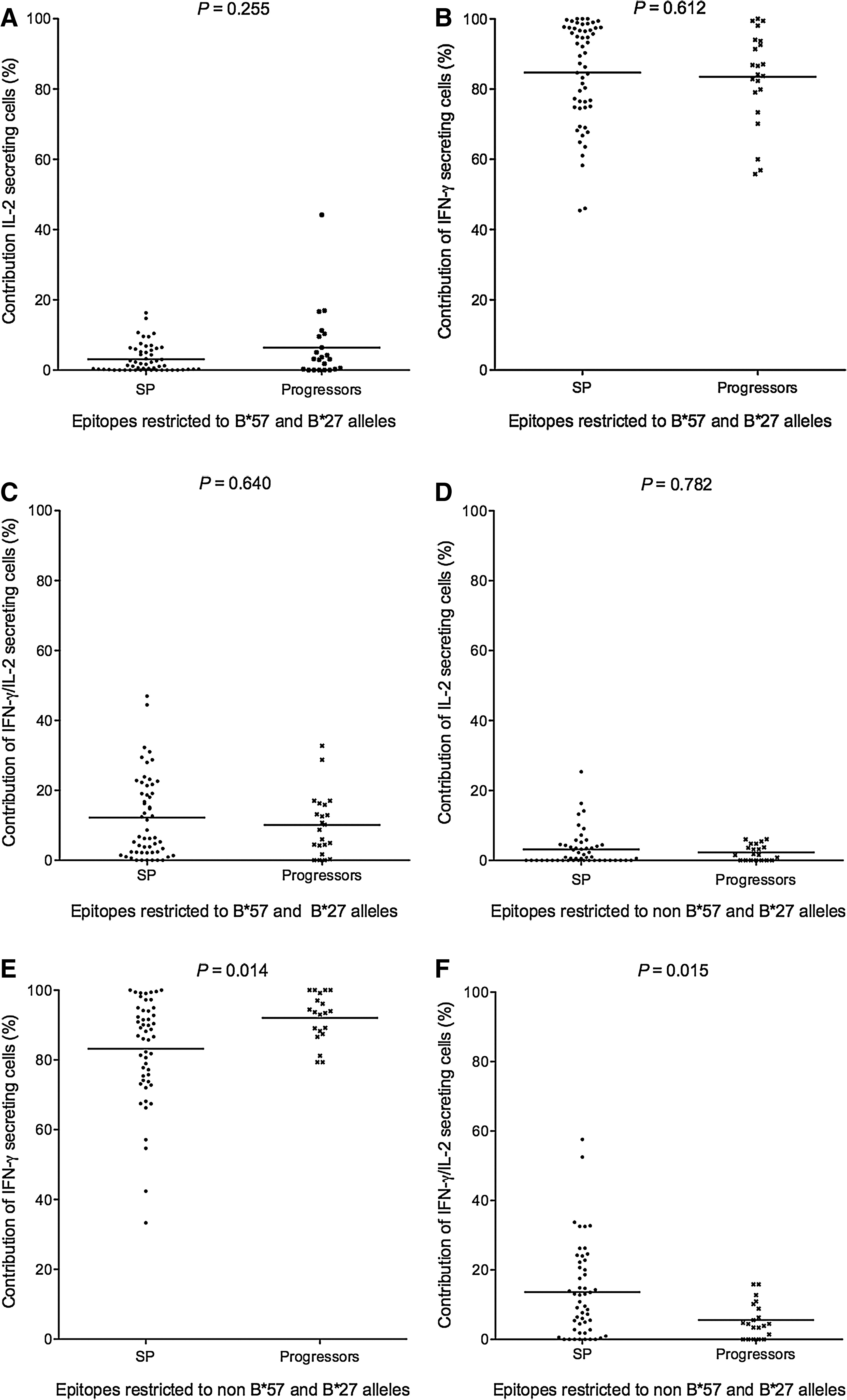
The contribution of three functional lymphocyte subsets to total HIV peptide-specific responses differs in slow progressors (SP) versus progressors, depending on the MHC class I alleles that restrict peptide recognition. Scatterplots illustrate the contribution of single IL-2-, single IFN-γ-, and dual IFN-γ/IL-2-secreting lymphocytes to the total magnitude of responses to individual stimulatory optimal epitopes restricted by HLA-B*57 and B*27 alleles in SP (n = 14) and progressors (n = 8) expressing these alleles (panels
The contribution of functional lymphocyte subsets to total HIV peptide-specific responses differs for responses to peptides restricted by protective versus other MHC class I alleles in cells from progressors, but not in SP subjects. Scatterplots illustrate the contribution of single IL-2-, single IFN-γ-, and dual IFN-γ/IL-2-secreting lymphocytes to the total magnitude of responses to individual stimulatory optimal epitopes restricted by HLA-B57 and B*27 versus other MHC class I alleles within SP (n = 14; panels
Next we compared the functional distribution of responses restricted by alleles other than HLA-B*57 and B*27 between all SP (n = 23) and progressor (n = 13) subjects expressing these MHC class I alleles or not. The between-group proportional contribution of single IL-2-secreting cells was similar, whereas single IFN-γ-secreting cells contributed less and dual IFN-γ/IL-2-secreting cells more to responses restricted by alleles other than HLA-B*57 and B*27 among SP (p = 0.014 and p = 0.015 by Mann-Whitney U test for single IFN-γ and IFN-γ/IL-2, respectively; Fig. 1D, E, and F). Together these results suggest that while epitopes restricted by HLA-B*57 and B*27 alleles maintain the potential to stimulate poly-functional immune responses in SP as well as in progressors, those restricted by other MHC class I alleles stimulate a response with more poly-functional characteristics in SP than in progressors.
Functional subsets are equally distributed for responses restricted by HLA-B*57 and B*27, versus other MHC class I alleles in SP, but not progressors
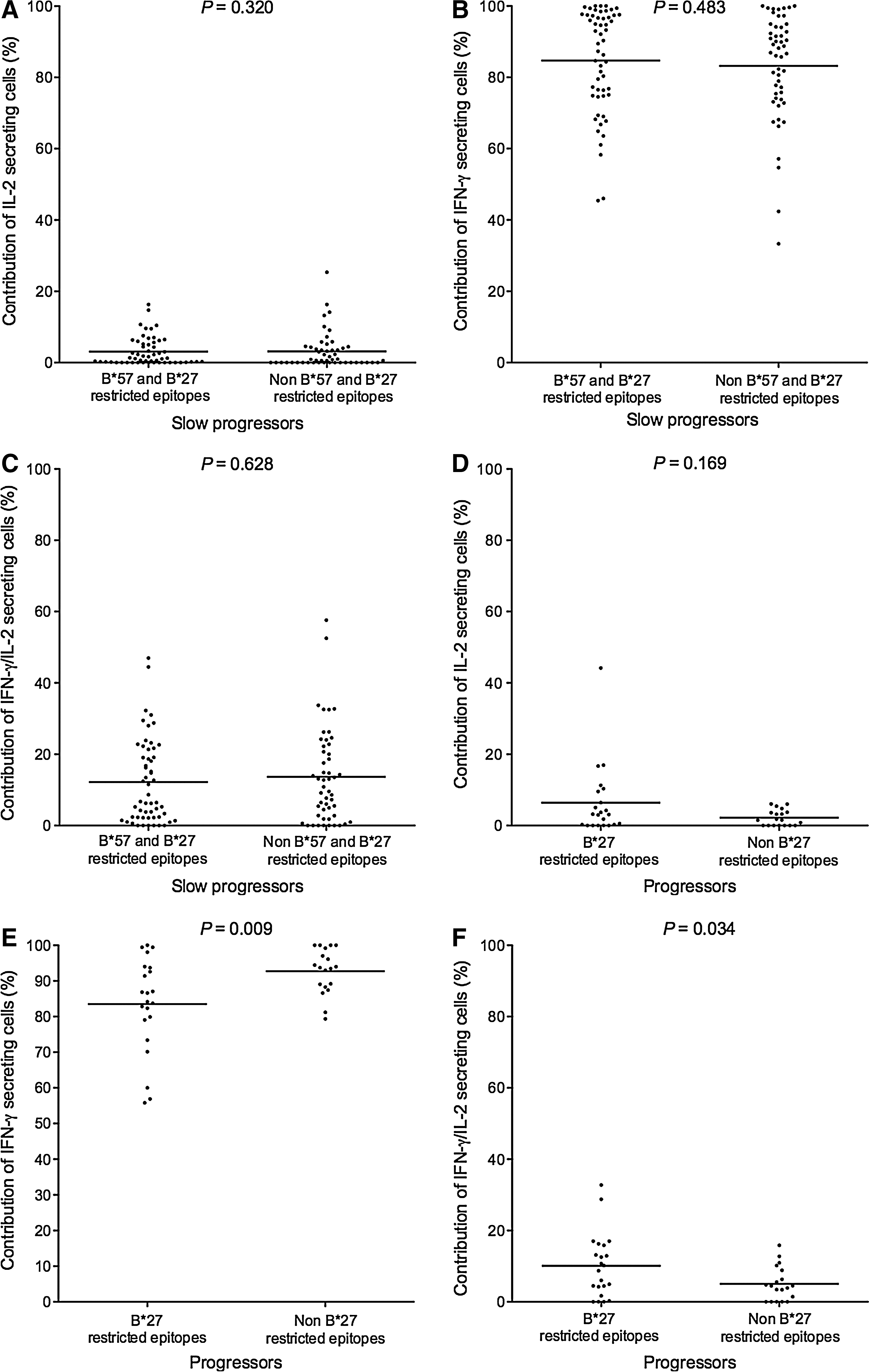
HIV-infected subjects expressing alleles associated with slow progression also express other MHC class I alleles associated with typical or rapid disease progression. We therefore assessed within SP and progressor subjects the contribution of functional subsets restricted by these two subsets of MHC class I alleles. Within the 14 SPs expressing HLA-B*57 and/or B*27, the contribution of single IL-2-, single IFN-γ-, and dual IFN-γ/IL-2-secreting lymphocytes was similar, whether stimulatory peptides were restricted by protective or other MHC class I alleles (Fig. 2A, B, and C).
There were only 8 subjects expressing B*27 among the progressor group. Despite the fact that the panel of stimulatory optimal peptides used to screen these individuals was of similar size to that used to screen SP (a median of 14 and 16 peptides, respectively; p = n.s. by Mann-Whitney U test), few responses to epitopes restricted by alleles other than B*27 were seen in progressors, a phenomenon also described by others (3). Therefore we were unable to compare within this group of 8 progressors the functional subset contribution to responses restricted by alleles associated with slow versus typical/rapid disease progression. To address this issue in another manner, we compared the contribution of functional subsets between the 8 progressors who expressed B*27, and 5 additional progressors who expressed neither of these MHC class I alleles (Table 1). These two groups of progressors had similar times from infection, CD4 counts, and VL at the time point tested (p = n.s. for all comparisons by unpaired t-test). The contribution of single IFN-γ secretion was lower and dual IFN-γ/IL-2 secretion greater to responses restricted by B*27 alleles, compared to responses restricted by other MHC class I alleles (p < 0.05 for both comparisons by unpaired t-test). Although statistical significance was not reached, we found a trend towards a greater contribution of single IL-2-secreting lymphocytes to epitopes restricted by protective alleles compared to other expressed MHC class I alleles (Mann-Whitney U and unpaired t-test; p = 0.169; Fig. 2D, E, and F). These results suggest that within SP, responses restricted by alleles associated or not with protection are similarly poly-functional, while in progressors, poly-functional responses are more likely to be a feature of responses restricted by MHC class I alleles associated with protection than responses restricted by other MHC class I alleles.
Discussion
Several studies have reported that the expression of individual HLA class I alleles have an impact on the natural history of HIV. Although the effect of certain alleles shows considerable variation due to race/ethnicity and HIV-1 subtype infection, HLA-B*27 and B*57 consistently associate with slower rates of disease progression (19,40). Another feature of HIV-specific immune responses in SP is their functional signature, characterized by proliferation and maintenance of IL-2, TNF-α, and IFN-γ secretion and cytolytic activity (5,20,23,32,36,37,53). Of these cytokines, secretion of IL-2 by HIV-specific cells is the most sensitive to extinction, whereas secretion of IFN-γ persists in the setting of high viral load (4,5,54). Using a dual-color ELISPOT assay detecting secretion of IL-2 and IFN-γ, we addressed whether functional heterogeneity of responses to individual epitopes was influenced by the restricting HLA allele, and whether alleles associated with protection preferentially stimulated poly-functional immune responses. Our findings show that epitopes restricted by protective alleles stimulate poly-functional responses in both SP and progressors. However, epitopes restricted by MHC class I alleles other than B*57 and B*27 contribute significantly more IFN-γ/IL-2-secreting lymphocytes in SP than in progressors. A within-group analysis of SP showed that the proportional contribution of single IL-2-, single IFN-γ-, and dual IFN-γ/IL-2-secreting cells was similar for responses restricted by B*57 and B*27 versus other MHC class I alleles. In contrast, in progressors the contribution of the functional subset secreting both IL-2 and IFN-γ was greater, while the contribution of the subset secreting IFN-γ alone was smaller for responses restricted by HLA B*27 alleles than the other MHC class I alleles.
Several groups have provided a rationale for the protective effect exerted by specific HLA alleles. Carrying B*57 alleles contributes to early defects in HIV fitness due to the selection of CTL escape mutations that reduce replicative fitness (9). This may in turn play a role in controlling VL and preserving poly-functional HIV-specific responses. The finding that the contribution of poly-functional lymphocytes is stronger when restricted by protective alleles is consistent with a study showing that B*57- and B*27-restricted CD8+ T cells isolated from chronically-infected individuals have a stronger proliferative index compared to epitopes restricted by other alleles (26). Poly-functional cells have higher levels of each of the functions they exhibit than lymphocytes with corresponding single functions (13). Therefore poly-functional cells may be superior to mono-functional cells at mediating antiviral responses. A previous study assessing poly-functional immune responses during chronic HIV infection found that the functional profile of HIV-specific cells was independent of HLA type and memory phenotype. However, T-cell functionality was measured using peptide pools corresponding to HIV gene products, and therefore could not address functional heterogeneity at the epitope level (5).
CTLs targeting identical epitopes may differ substantially in their ability to select for escape mutations when restricted by distinct HLA alleles. Unique peptide/MHC complexes were found to stimulate qualitatively different CTLs with distinct TCR clonotypes and functional avidities, thus reinforcing the notion that HLA alleles influence the function of the responding repertoire (2,31). Although we did not address the functional differences between identical epitopes presented by distinct MHC class I alleles, we found that the relative distribution of functional subsets differed based on the restricting MHC class I allele.
HIV-infected subjects expressing HLA alleles associated with rapid HIV disease progression recognize a smaller fraction of their potential viral epitope repertoire compared to those expressing alleles associated with slow disease progression (46). This finding was confirmed in a large cohort of individuals in primary infection, in whom responses restricted to HLA-B*57 and B*27 made a larger contribution to the total HIV-specific IFN-γ-secreting immune response compared to responses restricted by other co-expressed alleles (3,27). Furthermore, expression of either B*57 or B*27 also reduced the magnitude of responses restricted by other alleles co-expressed in the same individuals (3). Comparison of the functional makeup of responses in individuals that can be classified as SP and progressors revealed that the contribution of single IFN-γ- and dual IFN-γ/IL-2-secreting lymphocytes differed significantly only to responses restricted by MHC class I alleles other than HLA B*57 or B*27. This suggests that protective alleles maintain the potential to stimulate poly-functional immune responses in progressing individuals, while lymphocytes recognizing epitopes restricted by non-protective alleles are more susceptible to functional impairment. Recent studies reported that CTLs restricted to protective alleles imposed stronger selection pressure compared to responses restricted by other alleles (9,17). Although we did not analyze the relationship between functionality and viral sequence variation, it is tempting to speculate that poly-functional CTLs restricted by protective alleles impose stronger selection pressure compared to functionally-impaired CTLs. The presence of multi-functional responses in HIV-infected progressors may bring into question their role in controlling viral replication and disease progression. Although the progressor subjects were in the chronic phase of infection, their median time infected was 20.4 (8.3–55.9) mo. Therefore, at least half were infected for less than 2 y. It is possible that responses characterized by IFN-γ/IL-2 secretion restricted by protective HLA alleles take longer to be extinguished than those restricted by non-protective alleles.
Among progressing individuals, poly-functional immune responses contribute significantly less to the total immune response when restricted by non-protective alleles. Interestingly, a recent study has shown that epitope-specific responses restricted to protective alleles exhibit the most efficient cross-recognition of variants compared to epitope variants restricted to alleles associated with normal or accelerated disease progression (50). Antigenic variation in the context of non-protective restricting alleles potentially reduces the effectiveness of the responding CTL generating variant-specific lymphocytes with proliferative defects (51) and possibly impairment of IL-2 secretion. Of note, our results demonstrate that HIV-derived optimal epitopes, which according to size and sequence are limited to forming complexes with MHC class I alleles to stimulate CD8+ T cells, can stimulate cells secreting only IL-2 (Fig. 1A and D and 2A and D). Although the contribution of single IL-2 secretion to peptide-specific responses is clearly of low magnitude compared with IFN-γ/IL-2 and single IFN-γ secretion, certain epitopes stimulated CD8 responses that were above the threshold identifying positive responses.
In conclusion, our findings suggest that functional heterogeneity within an individual is influenced by the restricting HLA allele, and that associations observed between slow disease progression and the expression of protective alleles is partly explained by the ability of these molecules to present relevant determinants and stimulate poly-functional lymphocytes.
Footnotes
Acknowledgments
The authors wish to thank the study participants of the Montreal Slow Progressor (SP) and Primary Infection (PI) cohorts, and the study participants followed at the McGill University Heath Center Immunodeficiency Treatment Center. The work of the treating physicians enrolling and following subjects in the SP and PI cohorts is very much appreciated, in particular Drs. Roger LeBlanc, Julie Bruneau, and Jean-Guy Baril. We wish to acknowledge Ms. Nancy Simic and Marie-Pierre Boisvert for expert technical assistance, Ms. Louise Gilbert for clinical data management, Ms. Mélanie Lemay and Ms. Chantal Grignon for nursing support for the leukapheresis infrastructure, and Mr. Mario Legault and Ms. Linda Racicot, coordinators of the PI and SP cohorts, respectively. This work was funded by the Réseau du SIDA et Maladies Infectieuses du Fonds de Recherche en Santé du Québec (FRSQ), and by a grant from the Canadian Institutes for Health Research #HOP-86862. Y.P. was supported by a Ph.D. scholarship from the FRSQ, and J.-P. Routy is a scientific scholar receiving support from FRSQ.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.