
Abstract
To better understand the early virus-host interactions of the pandemic 2009 A(H1N1) viruses in humans, we examined early host responses following infection of human epithelial cell cultures with three 2009 A(H1N1) viruses (A/California/08/2009, A/Mexico/4108/2009, and A/Texas/15/2009), or a seasonal H1N1 vaccine strain (A/Solomon Islands/3/2006). We report here that infection with pandemic A/California/08/2009 and A/Mexico/4108/2009 viruses resulted in differences in virus infectivity compared to either pandemic A/Texas/15/2009 or the seasonal H1N1 vaccine strain. In addition, IFN-β levels were decreased in cell cultures infected with either the A/California/08/2009 or the A/Mexico/4108/2009 virus. Furthermore, infection with A/California/08/2009 and A/Mexico/4108/2009 viruses resulted in lower expression of four key proinflammatory markers (IL-6, RANTES, IP-10, and MIP-1β) compared with infection with either A/Texas/15/2009 or A/Solomon Islands/3/2006. Taken together, our results demonstrate that 2009 A(H1N1) viruses isolated during the Spring wave induced varying degrees of early host antiviral and inflammatory responses in human respiratory epithelial cells, highlighting the strain-specific nature of these responses, which play a role in clinical disease.
Introduction
The primary site for influenza A virus replication in humans is the respiratory epithelium (16). Infection of epithelial cells with influenza A virus leads to the production of inflammatory cytokines and chemokines that initiate an innate immune response by recruiting monocytes, macrophages, and other leukocytes to the site of infection (11,47). In response to influenza A virus infection, respiratory epithelial cells have been shown to produce antiviral cytokines such as interferon (IFN)-α/β (50), leukocyte chemoattractants such as RANTES/CCL5 (42), Eotaxin/CCL11 (37), interleukin (IL)-8 (12), migration inhibitory factor (MIF) (6), and MCP-1/CCL2 (5), and proinflammatory cytokines such as IL-6 (43). Importantly, in patients with culture-positive seasonal influenza illness, Kaiser et al. found a positive association between IL-6 levels and clinical symptoms (36). Furthermore, the more exuberant inflammatory cytokine response induced by highly pathogenic avian influenza (HPAI) H5N1 viruses in humans correlated positively with disease severity and fatal outcome (15). Findings from in vitro experimental models indicate that pandemic pH1N1 viruses induce relatively lower levels of cytokines and chemokines in human primary macrophages and dendritic cells compared to seasonal influenza viruses (45), or HPAI H5N1 virus (68). Given the fact that a substantial proportion of the population is serologically naïve to pH1N1 (27), it is important to better understand the inherent ability of pH1N1 viruses to cause severe disease in humans.
Using a well-established in-vitro model of influenza A virus infection (9,40,50,52), we investigated the impact of pH1N1 virus infection of human respiratory epithelial cells by comparing three pH1N1 viruses (2,4,14), evaluating virus infectivity and growth, the early host antiviral and inflammatory responses (31,33,61), and virus-induced cell death (13,39,40). Our results show that compared with a recent seasonal vaccine strain of H1N1 virus (A/Solomon Islands/3/2006, hereafter referred to as A/Sol), and more importantly among themselves, pH1N1 viruses demonstrate significant differences in early antiviral cytokine and inflammatory responses, as well as cell death.
Materials and Methods
Viruses, cell culture, and infection
In this study, we used three pH1N1 viruses isolated during the initial outbreak period from patients with distinct disease outcomes. Specifically, A/California/08/2009 (A/Cal) and A/Mexico/4108/2009 (A/Mex) were isolated from patients with non-fatal outcomes (2,4), and A/Texas/15/2009 (A/Tex) was isolated from a pediatric case with underlying medical conditions and a fatal outcome (14). A/Sol virus is a PR8 reassortant vaccine strain (lot # UC 405-8-2) provided by Sanofi Pasteur (Swiftwater, PA). A/Sol, A/Cal, and A/Mex were propagated by allantoic cavity inoculation of 10-day-old embryonated chicken eggs. A/Tex was propagated using Madin-Darby canine kidney (MDCK) cells (ATCC, Manassas, VA). Viral titers were determined by standard plaque assay, and are expressed in plaque-forming units (PFU)/mL (69). The human epithelial cell line A549, derived from a lung adenocarcinoma (ATCC), and MDCK cells were grown in Dulbecco modified Eagle medium (DMEM; Invitrogen, Carlsbad, CA), containing 10% fetal bovine serum, 100 U/mL penicillin, 100 μg/mL streptomycin solution, and 2 mM L-glutamine. Human lung mucoepidermoid (H292) cells (ATCC) were grown in RPMI 1640 (ATCC), containing 10% fetal bovine serum, 100 U/mL penicillin, 100 μg/mL streptomycin solution, and 2 mM L-glutamine. For A549 and H292 cell infections, virus was diluted in DMEM and RPMI-1640, respectively, both containing 0.3% bovine serum albumin (BSA). Cell monolayers were treated with 0.5% trypsin/EDTA (Invitrogen), washed, and 106 cells per well were seeded in complete medium in six-well plates and incubated at 37°C and 5% CO2 for 24 h. The cell monolayers were washed twice with DMEM or RPMI-1640, and virus was added in a volume of 200 μL, at 0.1 or 1 multiplicity of infection (MOI) in each well. After 1 h of incubation, the cell monolayer was washed, and 2 mL of DMEM or RPMI-1640 containing 0.3% BSA and 1 μg/mL of N-p-tosyl-l-phenylalanine chloromethyl ketone (TPCK)-treated trypsin (Sigma-Aldrich, St. Louis, MO) was added to the cells. All experiments and procedures involving pH1N1 virus isolates were performed in a biosafety (BSL)-2 facility using BSL-3 procedures.
Influenza NP-protein expression analysis by immunofluorescence assay
A549 cells (2 × 105 per well), plated on four-well chambered slides (Electron Microscopy Sciences, Hatfield, PA), were infected with 1 MOI of virus in 100 μL of 3% BSA in DMEM. At 4, 8, 16, and 24 h post-infection (hpi), the cells were fixed with 2% paraformadehyde for 20 min. The cells were permeabilized with 0.05% saponin (Sigma-Aldrich) in PBS for 30 min at room temperature, and blocked with Image-iT FX Signal Enhancer (Invitrogen), also for 30 min at room temperature. The slides were incubated with mouse anti-NP monoclonal A-3 antibody (62) at a 1:500 dilution in 0.05% saponin-PBS overnight at 4°C. This was followed by 2 h of incubation at room temperature with Alexa Fluor 594 goat anti-mouse secondary antibody (Invitrogen) at a 1:1000 dilution in 0.05% saponin-PBS. The cells were washed with 0.05% saponin-PBS, and incubated with 4,6-diamino-2-phenylindole (DAPI; Invitrogen) at a 1:1000 dilution for 5 min at room temperature. Slides were mounted with Slowfade Antifade reagent (Invitrogen), and viewed at 20 × magnification using an Axio Observer inverted fluorescence microscope (Zeiss, Thornwood, NY). Statistical analysis was performed for NP-positive counts per 100 cells counted in three representative fields.
Cell death analysis by MTT assay
As suggested by the manufacturer (Biotium, Hayward, CA), and subsequent to cell number optimization experiments, 10,000 cells in 100 μL of complete DMEM per well were added to 96-well plates. After 24 h, the cells were infected with 1 MOI of virus in DMEM with 0.3% BSA. At 16-, 24-, and 48-h time points, the infected cells, as well as an uninfected control, were treated with 10 μL of MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide], and incubated for 4 h at 37°C in 5% CO2. After incubation with MTT, the media was removed and the formazan crystals were solubilized in 200 μL of 2% paraformadehyde in dimethyl sulfoxide (DMSO). Absorbance was measured at 570 nm using a microplate reader (Spectramax Plus 384; Molecular Devices, Sunnyvale, CA), and cell viability (percentage) was determined by comparing the absorbance values of treated groups with untreated controls. Statistical analysis was performed on absorbance values from two independent experiments with four replicates per treatment group.
Quantification of IFN-β by qRT-PCR
The total RNA from cells was extracted using the RNeasy Mini Kit (Qiagen, Carlsbad, CA), and 2 μg of RNA was reverse-transcribed using the ThermoScript RT-PCR System (Invitrogen), in a volume of 20 μL after DNase I (Invitrogen) treatment. The resulting cDNA was diluted 1:10, and 2.5 μL was used in a SYBR green-based (SA Biosciences, Frederick, MD) real-time PCR reaction in a volume of 25 μL using an Mx3000 real-time PCR instrument (Stratagene, Cedar Creek, TX). PCR amplification of human IFN-β was carried out with the following primers: IFN-β forward, CTT ACA GGT TAC CTC CGA AAC TGA A, IFN-β reverse, GGT TGA AGA ATG CTT GAA GCA A. A housekeeping gene, β-actin with primers, forward ACC AAC TGG GAC GAC ATG GAG AAA, and reverse TAG CAC AGC CTG GAT AGC AAC GTA, was used to normalize the Ct values obtained in the real-time PCR reactions, which were then used to calculate fold changes compared to the uninfected sample using the ΔΔCt method.
Cytokine/chemokine analysis by Bio-Plex and ELISA assay
Following the manufacturer's protocol, a customized panel of 12 inflammatory cytokines and chemokines (IP-10, TNF-α, IL-6, IL-8, RANTES, VEGF, IL-1α, IL-12p70, MCP-1, MIP-1β, IL-4, and IFN-γ) was measured using a Bio-Plex suspension array system (Bio-Rad, Hercules, CA). Briefly, cell culture supernatants collected from uninfected and infected cells and assay standards were added to a 96-well filter plate, followed by anti-cytokine antibody-coupled beads, biotinylated bead detection antibodies, and finally phycoerythrin (PE)-conjugated streptavidin. The plate was read using a Bio-Plex suspension array system, and data were analyzed using Bio-Plex Manager 4.0 software (Bio-Rad) to calculate the observed concentrations (pg/mL) from standard curves. IFN-β concentrations in uninfected and infected cell culture supernatants were measured using a human IFN-β ELISA (PBL InteferonSource, Piscataway, NJ). Briefly, recombinant IFN-β or culture supernatants from infected or uninfected cell cultures were added to plates coated with anti-IFN-β antibodies, followed by biotinylated secondary antibody and streptavidin conjugated to horseradish peroxidase (HRP). Optical density was measured after the addition of tetramethyl-benzidine (TMB) substrate at 450 nm using a microtiter plate reader (BioTek, Winooski, VT). Observed concentrations of IFN-β (pg/mL) were determined using a standard curve.
NS1 protein analysis by Western blot assay
Cell lysates from infected A549 cells generated using Laemmli Sample Buffer (Bio-Rad) were subjected to SDS-PAGE on a 4–15% gradient gel (Bio-Rad). Proteins were transferred to a nitrocellulose membrane (Bio-Rad), and incubated in 5% BSA (Invitrogen) in 1 × Tween-PBS (Invitrogen) for 1 h at room temperature. They were then probed with mouse anti-NS1 monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA) at a 1:5000 dilution in blocking buffer overnight at 4°C, followed by incubation with HRP-conjugated anti-mouse goat antibody (Bio-Rad) at a 1:10,000 dilution for 1 h at room temperature. Anti-human β-actin mouse monoclonal antibody (Sigma-Aldrich) was used as a control. Bands were visualized and their intensities were estimated using a VersaDoc Imaging System (Bio-Rad).
Virus titer analysis by plaque assay
Confluent monolayers of MDCK cells in six-well plates were washed with DMEM and infected with serial 10-fold dilutions of supernatants from the infected cells for 1 h. After incubation with virus, the cells were washed with DMEM and overlaid with 1.6% SeaKem LE agarose (Lonza, Basel, Switzerland), mixed 1:1 with 2 × L15 medium (Lonza), containing 4 mM 4-(2-hydroxyethyl)-1-piperazine ethanesulfonic acid (HEPES), 2 mM L-glutamine, 5 μg/mL gentamicin, 1.5 mg/mL sodium bicarbonate, and TPCK-treated trypsin (Sigma-Aldrich). Virus plaques were stained with 0.3 % crystal violet solution (BD Biosciences, Sparks, MD), and counted after 48 h of incubation at 37°C in 5% CO2.
Statistical analysis
Statistical analyses were performed using an unpaired two-tailed t-test (GraphPad Prism 5.0; GraphPad Software, La Jolla, CA). The data are presented as mean ± SD. The differences were considered statistically significant when p < 0.05.
Results
pH1N1 virus-infected epithelial cells reveal virus isolate-specific differences in infectivity
To better understand the ability of the pH1N1 viruses to elicit early host responses, we used human lung epithelial cell lines susceptible to influenza infection as in-vitro models of influenza infection. We investigated the relative differences in host responses induced by three pH1N1 viruses (A/Cal, A/Mex, and A/Tex) in comparison with a seasonal vaccine strain of H1N1 virus (A/Sol) (9,50). First, using kinetic analysis of influenza NP expression by immunofluorescence assay as a measure of virus infectivity, we found that cell cultures infected with pH1N1 viruses, compared to A/Sol, demonstrated a significant delay in NP expression at 4 and 8 hpi (Fig. 1A and B). Among the pH1N1 isolates, NP expression was significantly lower in A/Mex- and A/Cal-infected cell cultures than in A/Tex-infected cell cultures at 8 hpi (A/Mex, p = 0.01; A/Cal, p = 0.003). At 16 hpi, when compared with A/Sol virus infection, the proportion of NP-positive cells in cultures infected with either A/Mex or A/Tex virus (but not A/Cal virus) was significantly lower (A/Mex, p = 0.01; A/Tex, p = 0.004; Fig. 1B). Next, we compared the kinetics of virus growth using an MDCK cell plaque assay to quantify virus in supernatants harvested from virus-infected A549 or MDCK cell cultures. As shown in Fig. 2A, we found that all four H1N1 viruses grew efficiently, and in fact, that by 48 hpi they had comparable peak virus titers. Similar virus-growth kinetics were observed in MDCK cells (Fig. 2B), suggesting that comparable growth characteristics of pH1N1 viruses are not restricted to a single epithelial cell type.
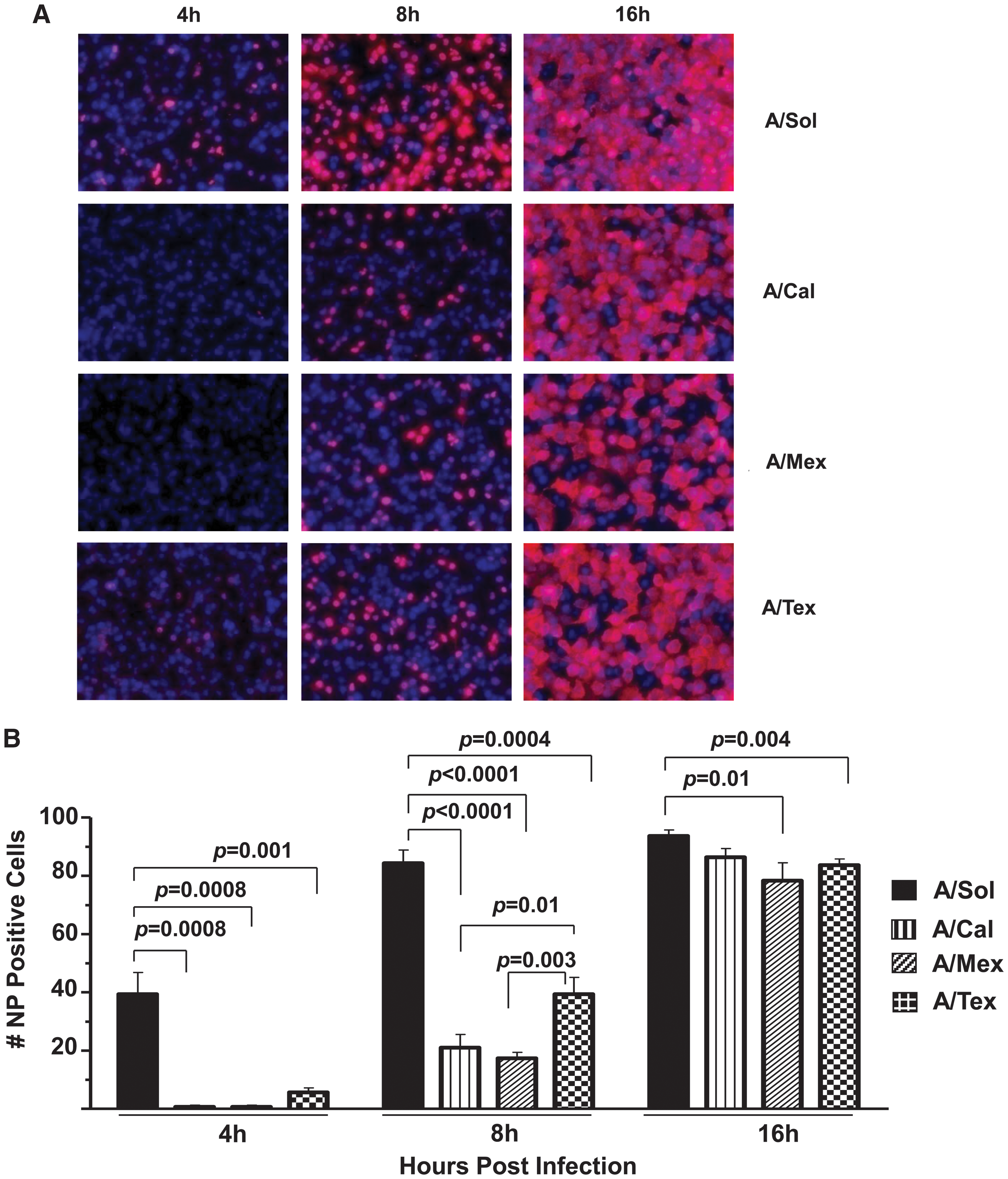
Infection with pH1N1 viruses reveals isolate-specific differences in influenza NP protein expression. (
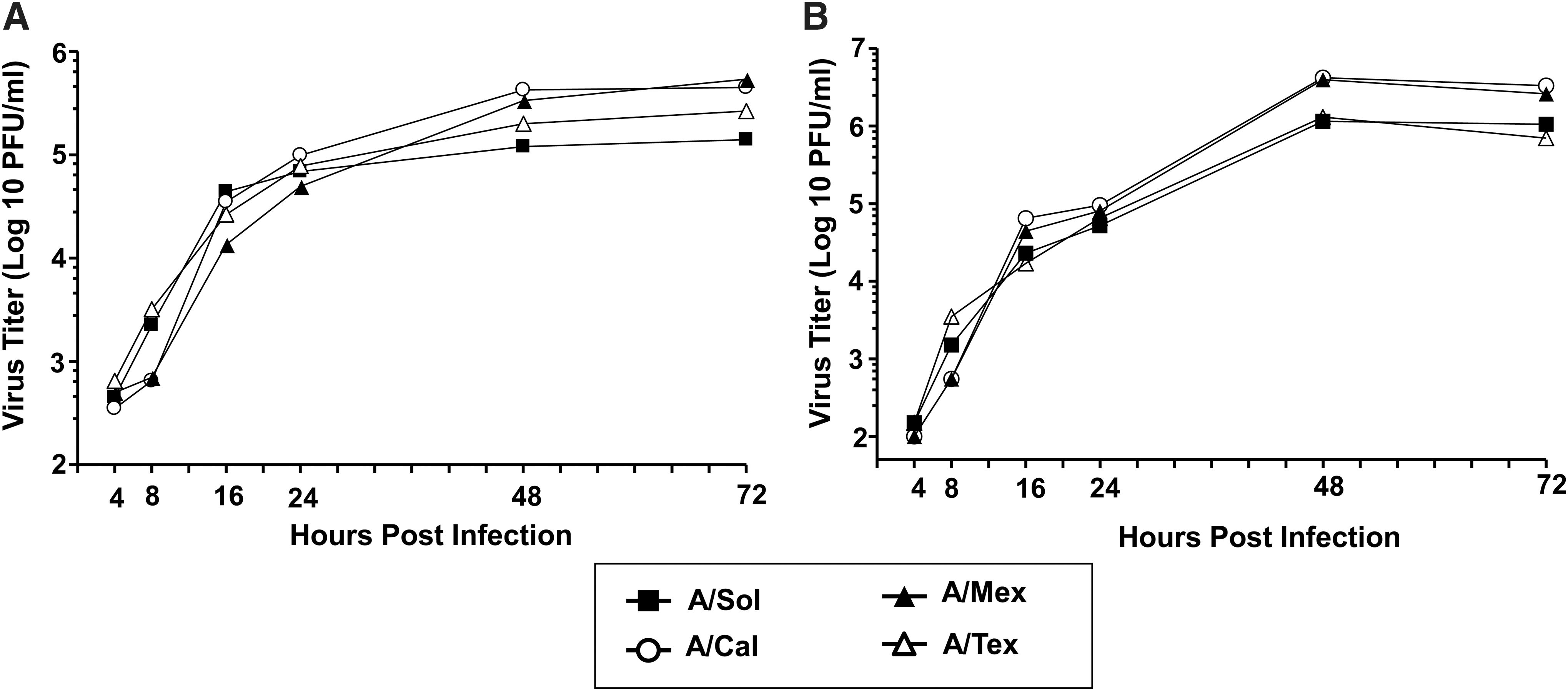
In-vitro growth characteristics of pH1N1 viruses. Cell culture supernatants from A549 (
pH1N1 virus-infected epithelial cells demonstrate virus isolate-specific changes in NS1 and IFN-β expression
Differences in infectivity among the three pH1N1 isolates (Fig. 1) prompted us to study potential differences in expression of the NS1 protein, an influenza protein with multiple functions, including regulation of viral RNA replication during infection (18,67). Analysis of NS1 protein expression at 16 hpi revealed that the A/Mex and A/Cal isolates, which yielded lower infectivity at early time points (Fig. 3), although not statistically significant, contained relatively higher NS1 protein levels compared with either A/Tex- or A/Sol-infected cell cultures (Fig. 3A).
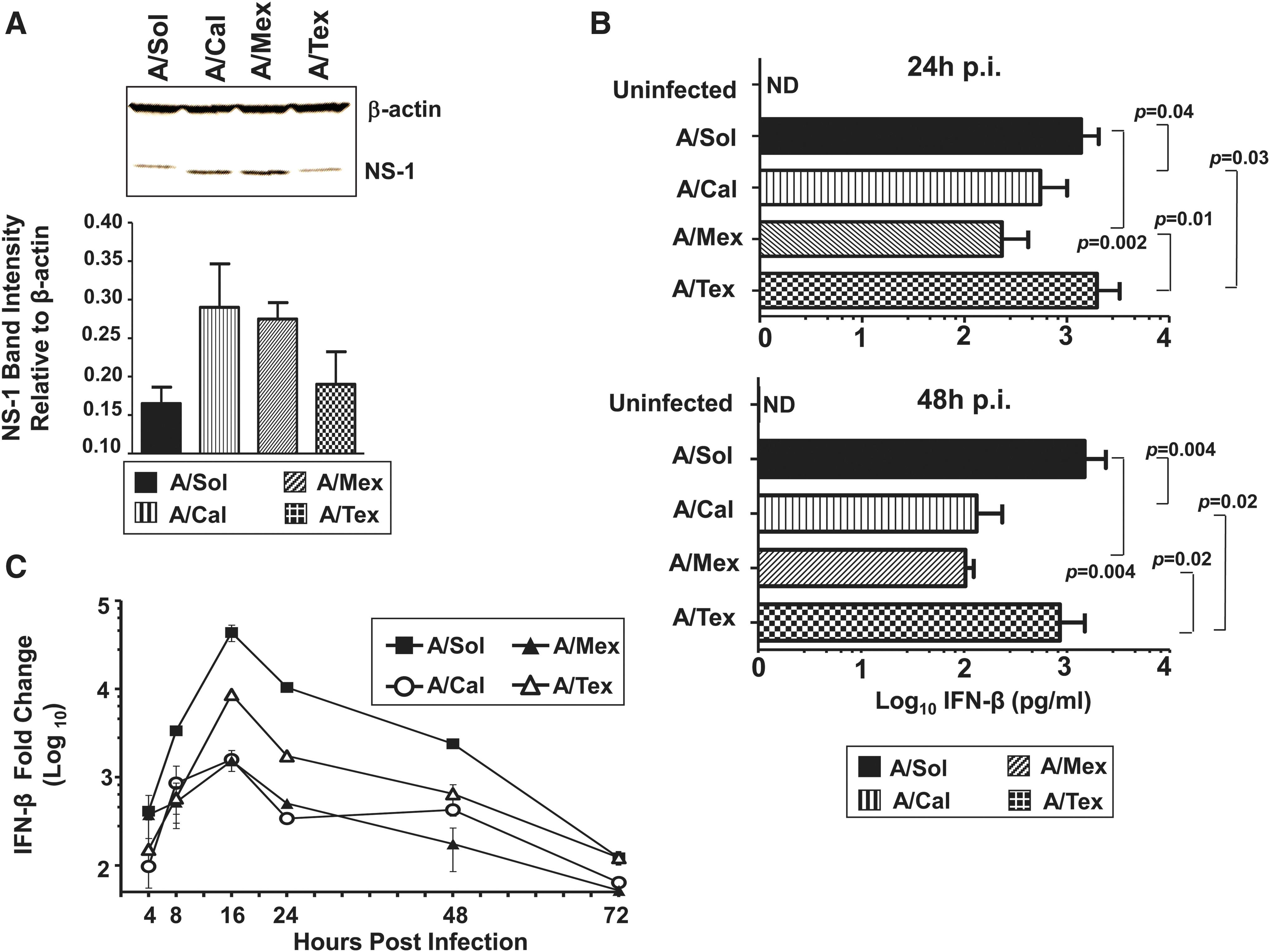
Infection with pH1N1 viruses reveals strain-specific differences in host IFN-β and influenza NS1 protein levels. (
Although strain-specific and multifunctional (26,30), the well-established antagonistic action of NS1 on the host innate immune response, and IFN-α/β-dependent responses in particular (17,22), prompted us to study pH1N1-induced IFN-β production. First, we examined IFN-β protein levels in virus-infected cell culture supernatants at 24 and 48 hpi. We found that among the pH1N1 viruses, cell cultures infected with A/Mex and A/Cal viruses expressed significantly lower IFN-β levels compared to A/Tex and A/Sol infections at 24 and 48 hpi (Fig. 3B). An analysis of IFN-β transcript levels in cell cultures infected with H1N1 isolates revealed that among the pH1N1 viruses, IFN-β mRNA expression was markedly higher in A/Tex virus-infected cells at 16 and 24 hpi, compared to cultures infected with either A/Cal or A/Mex virus, with levels dropping to the pre-infection range by 72 hpi (Fig. 3C). However, A/Sol virus infection, compared to the three pH1N1 viruses, yielded the highest levels of IFN-β transcripts by 8 hpi, and levels remained high through 48 hpi.
pH1N1 virus-infected epithelial cells demonstrate significant differences in expression of inflammatory mediators among different isolates
Influenza virus entry in susceptible cell types results in virus replication and expression of inflammatory mediators (29,34,43,54,55,60). The overall differences in the induction of the type I interferon response (Fig. 3B) observed among pH1N1 viruses, and between pH1N1 viruses and the seasonal H1N1 vaccine strain (A/Sol), led us to investigate possible differences in other virus-induced epithelial cell-specific inflammatory mediators. A549 cell cultures infected with the pH1N1 viruses or the A/Sol virus were harvested at 48 hpi and assayed for 12 different inflammatory markers by Bio-Plex assay. Interestingly, we found marked differences in the expression of four key inflammatory mediators, namely IL-6, IP-10, RANTES, and MIP-1β, among the pH1N1 viruses, and between the pH1N1 viruses and A/Sol virus (Fig. 4). Of note, the levels of IL-6 and IP-10 were significantly elevated in supernatants from A/Tex virus-infected cells compared with other pH1N1 viruses (IL-6, three- to sevenfold; IP-10, two- to threefold). Additionally, levels of RANTES and MIP-1β were higher in supernatants from A/Tex virus-infected cells compared with those from cultures infected with the other pH1N1 viruses, although A/Sol virus-infected cell cultures elicited the highest levels of all of the H1N1 viruses we tested. A/Mex virus-infected cells exhibited the lowest level of expression of all four inflammatory mediators (Fig. 4). Furthermore, infection of H292 cells, another epithelial cell line previously shown to be susceptible to influenza virus infection (9), with the four H1N1 viruses demonstrated similar changes in inflammatory mediators (data not shown). Overall, infection with the A/Tex or A/Sol viruses resulted in the highest levels of inflammatory mediators, possibly reflecting the early NP expression observed with these two viruses at early time points (Fig. 1). Moreover, of six other inflammatory mediators analyzed (TNF-α, IL-8, VEGF, IL-1α, IL-12p70, and MCP-1), IL-8 and MCP-1 were detectable above the control uninfected cultures, but the differences among the four H1N1 viruses were not statistically significant (data not shown). These results suggest that infection with different pH1N1 viruses leads to significant variation in the expression of key inflammatory mediators.
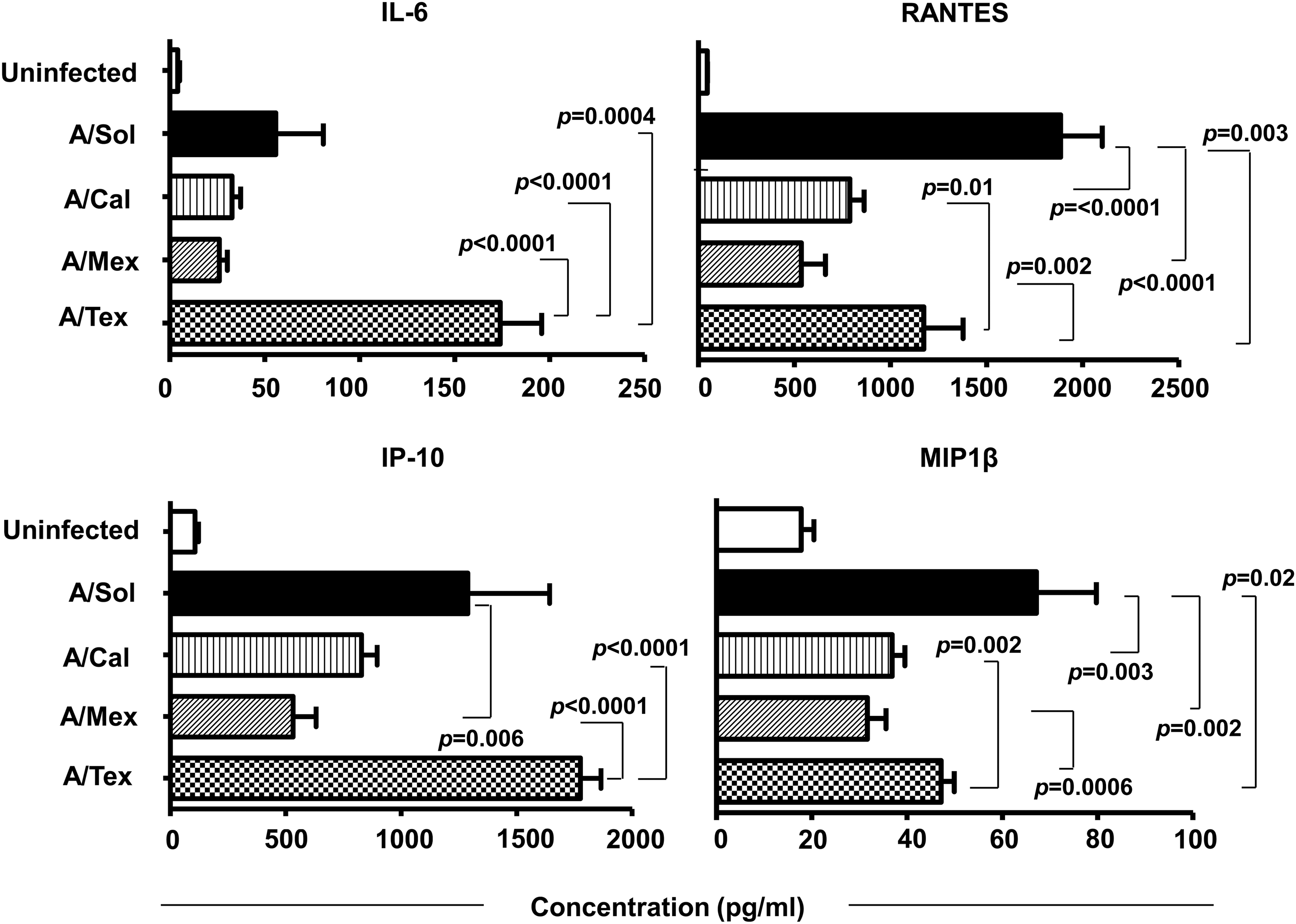
pH1N1 virus-infected epithelial cells express altered levels of proinflammatory mediators. Cell culture supernatants from A549 cells infected with 1 MOI of A/Sol or different pH1N1 viruses were collected at 48 hpi and analyzed for inflammatory cytokine/chemokine levels by Bio-Plex assay, as described in the Materials and Methods section. Data are the mean of two independent experiments. Error bars represent mean with SD.
pH1N1 virus-infected epithelial cells demonstrate virus isolate-specific changes in virus-induced cell death
The differences seen in virus infectivity (Fig. 1), early antiviral defense (Fig. 3B), and production of inflammatory mediators (Fig. 4), led us to study the fate of cells infected with pH1N1 viruses. Using an MTT assay (39), we determined the cell death kinetics in response to infection of A549 cells with the pH1N1 viruses or the A/Sol virus. As shown in Fig. 5, relative to infection with A/Sol, a significantly lower percentage of cell death was detected at 24 hpi in cultures infected with pH1N1 viruses. By 48 hpi, the difference between the viability of cells infected with the pH1N1 viruses and the A/Sol virus was even greater (p < 0.0001). Interestingly, among the three different pH1N1 viruses, at 48 hpi cell cultures infected with A/Tex virus exhibited the highest levels of cell death (Fig. 5). Taken together, these results demonstrate that among the three pH1N1 viruses tested, A/Mex and A/Cal isolate-infected cultures showed a comparative (1) delay in infectivity, yet overall comparable virus titers (Figs. 1 and 2), (2) an increase in NS1 protein expression (Fig. 3A), (3) a decrease in antiviral cytokine expression (Fig. 3B and C), (4) a reduction in expression of inflammatory mediators (Fig. 4), and (5) a significantly lower percentage of cell death (Fig. 5), relative to the seasonal H1N1 vaccine strain and the A/Tex pH1N1 virus.
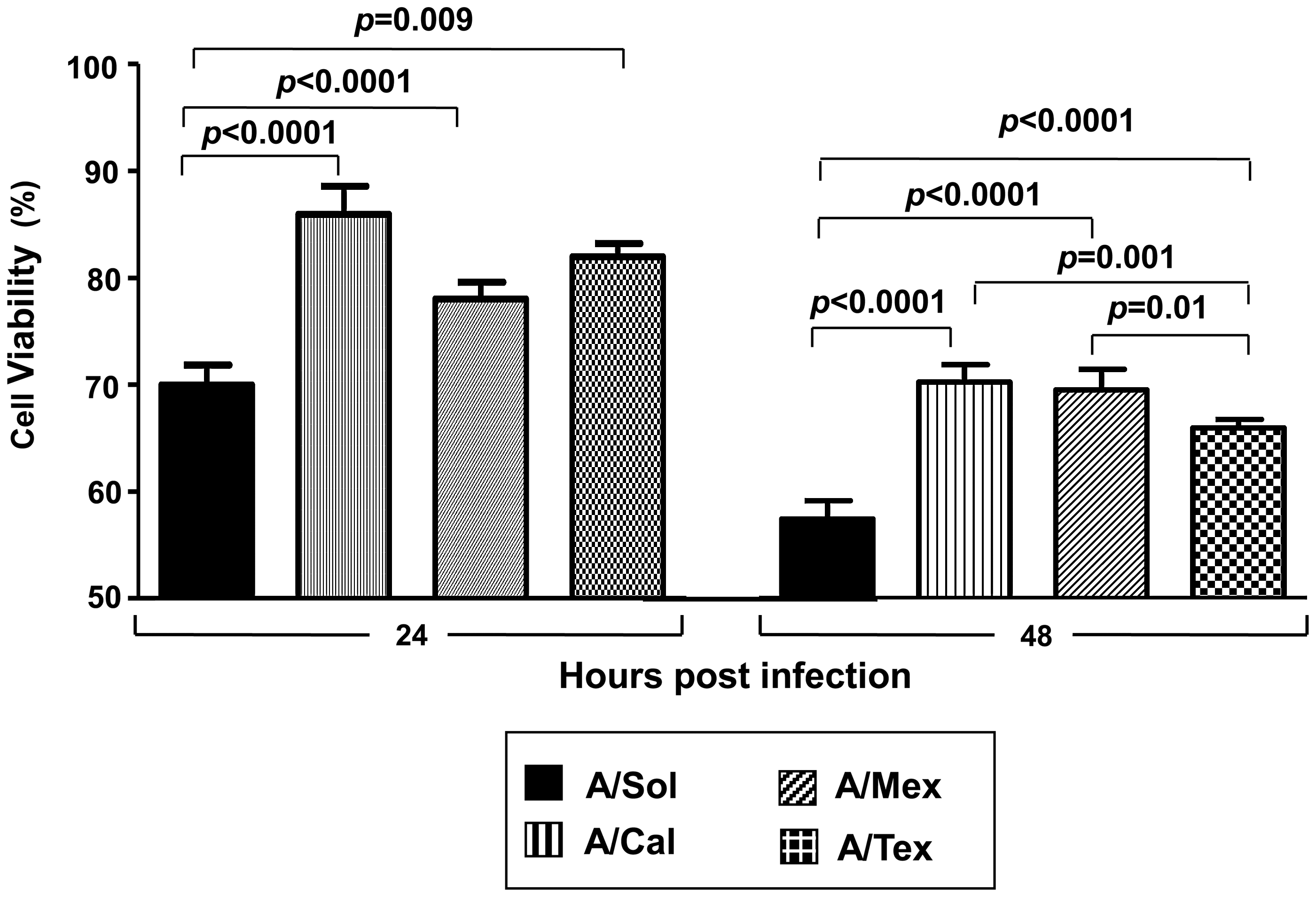
Infection with pH1N1 viruses reveals differences in cell viability. A549 cells infected with 1 MOI of A/Sol or the pH1N1 viruses were harvested at 24 (left panel) and 48 hpi (right panel), for cell death analysis by MTT assay, as described in the materials and methods section. Data represent results of two independent experiments, with each experiment consisting of four replicate values for each virus. Error bars represent mean with SD.
Molecular differences among pH1N1 viruses
Our results highlight the differences in infectivity and inflammatory response among pH1N1 viruses, A/Tex in particular, in an in-vitro model of influenza infection. The pH1N1 viruses used in this study were propagated in either embryonated chicken eggs or in a MDCK cell culture system. It is conceivable that the differences seen in the host response to pH1N1 infection, particularly A/Tex virus, could be either due to differences in the cell substrates used for propagation of virus (cell culture for A/Tex versus egg inoculation for the A/Cal and A/Mex isolates), or the inherent differences among the three viruses. To address the former possibility, we first compared the full genome sequences originally described by Garten et al. for A/Tex, A/Cal, and A/Mex viruses (23), with those of the actual virus stocks used in this study. No difference from the original deduced amino acid sequences were noted (data not shown). However, a comparison of amino acid sequences among the three pH1N1 isolates revealed differences in three of the eight gene segments (HA/125M and 227L; NP/D16; NS1/R45; and NS2/K67) of the A/Tex virus compared with the other two pH1N1 viruses used in the study (Table 1). Therefore, it is plausible that differences in the viral genomes may have contributed to the observed differences in antiviral and proinflammatory cytokine response detected for the A/Tex pH1N1 virus.
2009 A(H1N1) numbering (23).
The virus isolates used in this study were analyzed for the differences in amino acid sequences in different gene segments of influenza virus (NCBI Genbank and Fludb sequence search). All other sequences for PA, HA, NP, M2, NS1, and NS2 segments, apart from the differences shown (underlined) in this table, were identical among the three pH1N1 isolates.
NC, no change.
Discussion
Studies with pandemic influenza viruses in experimental models can help our understanding of the inherent differences among virus strains and their contribution to biological outcomes of infection. In this report, we examined the relative infectivity and early host response induction of newly emergent pH1N1 viruses using an in-vitro cell culture model of influenza infection. Infection of epithelial cells with three pH1N1 virus isolates resulted in early host responses distinct from a seasonal vaccine strain of H1N1 infection, and also revealed striking differences among the pH1N1 viruses. Among pH1N1 viruses, the A/Mex and A/Cal viruses demonstrated a marked delay in viral protein expression, compared to A/Tex virus; however, it grew efficiently by 16 hpi, and in fact achieved comparable virus titers at later time points. The delay in infectivity with either the A/Mex or A/Cal isolate was also reflected in comparatively lower levels of antiviral cytokines, as well as reduced virus-induced inflammatory markers and cell death. In addition, changes in the amino acid composition of gene segments were noted in A/Tex, the pH1N1 strain that triggered relatively higher antiviral and anti-inflammatory responses. Taken together, our findings reveal differences among closely-related pH1N1 viruses in their induction of host inflammatory responses.
The newly emergent pH1N1 viruses possess gene segments originating from influenza viruses of swine, human, and avian origin (23,51). Therefore, in our studies employing pH1N1 viruses, we first considered investigating differences in infectivity and growth characteristics between pandemic and seasonal H1N1 viruses. Even though delayed kinetics of NP expression in A/Mex and A/Cal virus-infected cell cultures, compared to A/Tex and A/Sol infection, correlated well with lower virus growth at an early time point (8 hpi), the virus titers were comparable at later time points (48 and 72 hpi). Of note, the viruses that demonstrated rapid NP expression (A/Tex and A/Sol) also triggered relatively higher levels of inflammatory markers and cell death.
Studies have shown that NS1 gene expression antagonizes the type 1 IFN response, and as a result, virus persists at higher titers in susceptible cell types (20,24,65). In the present study, pH1N1 viruses that failed to elicit a potent type 1 IFN response (A/Mex and A/Cal) expressed higher levels of NS1 (Fig. 3A), compared to either A/Tex- or A/Sol-infected epithelial cell cultures, but the differences were not statistically significant. Furthermore, the pH1N1 viruses used in this study possessed a single residue change in NS1 at residue 45 (glycine in A/Mex and A/Cal; arginine in A/Tex). Furthermore, a search of the NCBI Genbank and Fludb sequence databases revealed that among NS1 genes from a total of 4538 influenza A viruses, the G45R substitution in NS1 is rare, and occurs in less than 0.3% of human seasonal and pandemic H1N1 strains (data not shown). Residue 45 of NS1 is part of the RNA-binding domain, and is possibly involved in maintaining and stabilizing the NS1-RNA complex (26,48,64). Further studies with these pH1N1 viruses are needed to support a strong negative correlation of NS-1 with IFN-β production, and also to investigate a potential role of the G45R mutation in IFN-β expression and function.
Since influenza virus infection leads to production of inflammatory mediators in experimental models (8,41), we evaluated virus-infected cell culture supernatants for potential differences in virus-induced inflammatory mediators. Using two different influenza-A-virus-susceptible cell lines, we found that of the 12 different inflammatory mediators tested, levels of four key chemotactic markers (IL-6, IP-10, RANTES, and MIP-1β) were significantly different in virus-infected cell culture supernatants. Surprisingly, among all four H1N1 viruses tested in our model, levels of IL-6 and IP-10 were highest in cell culture supernatants harvested after A/Tex infection. IL-6, a cytokine known to be involved in the induction of fever and the acute phase response (25,53), and IP-10, a potent chemoattractant for the recruitment of monocytes, NK cells, T cells, and dendritic cells (19,57), could contribute to the differences seen in inflammation and disease severity caused by different pH1N1 viruses. Similarly, Svitek et al. reported that the relative severity of clinical signs in ferrets infected with seasonal H1N1 and H3N2 viruses correlated with stronger IL-6 upregulation (56). Furthermore, a previous clinical study highlighted the association between increased levels of IL-6 and influenza-induced clinical illness (36). Experimental evidence also suggests a positive correlation between cellular damage and proinflammatory cytokine induction. For example, Wang et al. found that excessive release of proinflammatory cytokines in avian influenza A virus (H5N1)-infected mouse microglia and astrocytes was associated with a concomitant increase in apoptosis (63). Consistent with these findings, among the four H1N1 viruses tested here, cultures infected with either A/Tex or A/Sol demonstrated significantly higher rates of cell death, compared to those infected with A/Mex or A/Cal. Further studies with these pH1N1 viruses in in-vivo models of influenza infection are needed to establish the biological relevance of our in-vitro findings.
In-vitro studies using human macrophages have documented relatively lower levels of pH1N1 virus-induced cytokines compared with either seasonal strains of influenza (45), or a highly pathogenic H5N1 virus (68). In contrast to other studies, which used pH1N1 viruses and demonstrated an attenuated inflammatory and antiviral phenotype, we used three pH1N1 viruses, and surprisingly, found a pH1N1 strain-specific response with A/Tex that differed from A/Mex and A/Cal in our experiments, and from other studies that generally demonstrated an attenuated inflammatory and antiviral phenotype. The attenuated host inflammatory response by A/Mex and A/Cal pH1N1 viruses seen in our experiments, compared with the seasonal H1N1 virus, has also been observed in polarized bronchial and pharyngeal epithelial cells that exhibit many features of human airway epithelium (Zeng and Tumpey et al., personal communication). Given the previous findings of influenza virus-induced expression of inflammatory mediators in chemotaxis (5,31,35), we speculate that the observed differences in indicators of inflammation induced by infection with pH1N1 viruses may recruit different cell types, leading to varying degrees of inflammation and disease severity.
In the studies presented here, we used pH1N1 viruses to study early host responses in a controlled experimental setting, and demonstrated strain-specific differences in the induction of host antiviral and inflammatory responses. In addition to inherent strain-specific differences in the induction of host responses, a number of other host factors such as age, nutritional status, host genetics, and most importantly, pre-existing immunity to influenza A viruses, are also likely to contribute to the clinical outcomes of pH1N1 viral infections. Notably, recent reports of hospitalized cases of pH1N1 suggest that more than half of the patients had underlying medical conditions, including pregnancy, obesity, asthma, diabetes, and other risk factors (1,28,32,44,46,59). While our findings highlight the inherent strain-specific differences in the induction of early host responses, due to the strong association of risk factors with outcomes of pH1N1 infections, it is difficult to establish a causal relationship between our in-vitro data and what has been observed in humans. Further studies addressing the link between the genetic composition of pH1N1 viruses and inherent variability in host responses are needed to better assess and modify the antiviral regimens used against pandemic influenza A viruses.
Footnotes
Acknowledgments
We thank various members of the Influenza Division, Centers for Disease Control and Prevention, including Kathy Hancock, Hong Sun, and Taronna R. Maines, for provision of reagents and constructive comments on procedures; Catherine Smith and Michael Shaw for providing genome sequences of the viruses used in this study; and J. Bradford Bowzard and Andrew K. Taylor for helpful comments on the manuscript.
Author Disclosure Statement
No competing financial interests exist.