
Abstract
Kaposi's sarcoma-associated herpesvirus (KSHV) is the etiological agent of Kaposi's sarcoma (KS), the most common cancer in individuals with untreated HIV/AIDS. Host control of KSHV infection and KS oncogenesis by CD8 T cells remains underexplored. Although KSHV CD8 epitopes have been identified, the responses they elicit are weak and little is known about their relative importance. We sought to make a direct comparison of the recognition of a selection of the best-described known epitopes by a cohort of KSHV-seropositive, HIV-co-infected individuals, in order to assess the relative dominance of these epitopes. We further sought to identify novel epitopes from within a candidate immunogenic protein encoded by KSHV ORF28. MHC binding and denaturation assays identified putative novel A*0201-restricted epitopes from within the late-lytic glycoprotein ORF28. Recognition of these candidate epitopes was tested in a cohort of KSHV-seropositive, HIV-1-seropositive, A*0201-positive individuals by ex vivo ELISPOT, and compared with recognition of nine previously described epitopes. One novel late-lytic epitope from ORF28 was recognized by 7.1% of individuals, and was used for further investigation of KSHV-specific T cells using multimer technology. One known late-lytic epitope from the glycoprotein-encoding K8.1 was recognized by 71.4% of individuals, and represented an immunodominant KSHV epitope, but was too hydrophobic for multimer synthesis. This study identifies two KSHV CD8 epitopes derived from late-lytic antigens that are recognized by KSHV-seropositive, HIV co-infected individuals, and will be useful in future immunological studies into the CD8 response against KSHV in similar patient cohorts.
Introduction
There are several lines of evidence to indicate that KS oncogenesis is associated with the loss of CD8 T-cell mediated control of KSHV-infected cells. In immunocompetent individuals, KSHV can establish lifelong asymptomatic infection. However, upon immunosuppression (acquired or iatrogenic) both KSHV seroprevalence and the incidence of KS in KSHV carriers dramatically increase (4). Moreover, immune reconstitution following HAART often results in clinical improvement of KS (6) that is associated with recovery of KSHV-specific CD8 T-cell responses (5,24). Longitudinal studies of a small number of individuals with relapsing-remitting KS found a correlation between loss of KSHV-specific T cells and recurrence of active disease (1,13). Both the frequency and the antigenic repertoire of KSHV-specific CD8 T cells are greater in asymptomatic carriers of KSHV compared to individuals with KS (9,13).
A few KSHV-specific CD8 epitopes have been described (5,7,9,10,13,15,17,22), but the responses elicited by these epitopes are generally weak compared to responses elicited by other viruses such as HIV-1 and Epstein-Barr virus (EBV; also known as human herpesvirus 4) (3,7). Furthermore, the relative importance of the known epitopes remains largely unexplored and no immunodominant epitopes have been identified. For example, Table 1 shows all published HLA-A*0201-restricted KSHV epitopes with an accumulated tally of the number of responses observed for each of these epitopes by all individuals (from all publications) in which the epitope has been tested. Strikingly, only 3 of these 21 epitopes have been confirmed by more than one study (K12-LLNGWRWRL, ORF8-LMWYELSKI, and ORF22-FLNWQNLLNV), and fewer than half (10 of 21) have been tested in more than 10 individuals. Of these 10 epitopes, only 6 elicited responses in more than 5% of individuals tested (K12-LLNGWRWRL, ORF6-VLGDEVLSL, ORF8-LMWYELSKI, ORF61-GLADVFAEL, ORF65-NMSQAEYLV, and ORF22-FLNWQNLLNV). For these six epitopes, the percentage of responders to each epitope ranged from 16–46% of all individuals (HIV-negative and HIV-positive) tested, and 13–32% of all HIV-positive individuals tested. It seems reasonable to conclude that there is no immunodominant HLA-A*0201 epitope evident from the compiled data from these studies.
Indicates according to expression in PEL cells (11,12); † indicates epitopes used in the present study; Nt indicates not tested.
KSHV, Kaposi's sarcoma-associated herpesvirus; ORF, open reading frame.
Identification of immunodominant epitopes would be a useful tool for further exploration of the KSHV-specific CD8 T-cell response, for example, in the creation of KSHV-specific multimers in order to perform characterization of the phenotypes and functions of KSHV-specific T cells, an area that remains largely unexplored. We therefore sought to make a direct comparison of the recognition of nine of the most well-described KSHV epitopes derived from a range of KSHV gene products expressed across the spectrum of the virus's life cycle (latent, immediate-early-lytic, early-lytic, and late-lytic; Table 1, marked †). We chose to focus on HLA-A*0201-restricted epitopes, since this HLA type is one of the most common in individuals of all ethnic origins. In this way we hoped to determine the relative importance of these epitopes, in order to identify those that would be the most useful in future immunological investigations.
Our recent studies indicated that the KSHV CD8 T-cell response preferentially targets antigens expressed in the early- and late-lytic phases of the KSHV viral life cycle (18). In particular, the late-lytic envelope glycoprotein-encoding gene ORF28 (25) was identified as a likely immunogenic candidate (18). Viral glycoproteins are often targets of anti-viral T-cell responses (20,21). Three other KSHV glycoproteins (glycoprotein B, encoded by ORF8; glycoprotein H, encoded by ORF22; and glycoprotein 35/37, encoded by K8.1) are documented targets of the KSHV-specific CD8 T-cell response (15,22,24). We thus additionally sought to identify novel CD8 epitopes from within KSHV ORF28. Overlapping nine-mer peptides were synthesized spanning the entire ORF28 amino-acid sequence, and peptide-A*0201 binding assays were used to identify candidate epitopes. We then compared recognition of these epitopes with recognition of the nine selected previously identified epitopes by the same cohort of KSHV-seropositive individuals.
Materials and Methods
Study participants
All study participants provided written informed consent. The study was approved by Riverside Research Ethics Committee. Seventeen HIV-1-seropositive individuals with a history of KSHV-related neoplasia (KS or multicentric Castleman's disease [MCD]) were recruited from Chelsea and Westminster Hospital, London, U.K. Fourteen were HLA-A*0201-positive, and three were A*0201-negative. All participants were male, with a median age of 45 y (range 29–60 y). The median CD4 cell count was 468 cells/mm3 (range 48–1467 cells/mm3), plasma HIV-1 viral load was undetectable (<50 copies per mL) in 71% of individuals, and <400 copies per mL were found in 94%. One A*0201-positive individual who had a plasma HIV-1 viral load of 11,164 copies per mL was not receiving HAART, but had done so previously. All other HIV-1-positive participants were receiving HAART. One A*0201-positive individual had active KS, while all of the rest were in remission of KSHV-related neoplasia (KS or MCD). Clinical characteristics of KSHV-seropositive individuals are summarized in Supplementary Table 1 (see online supplementary material at
HLA typing
For high-resolution HLA subtyping of KSHV-seropositive individuals, genomic DNA was extracted from 200 μL peripheral blood using a QIAamp DNA Blood Mini Kit (Qiagen, Crawley, U.K.) according to the manufacturer's instructions. HLA typing was performed by Anthony Nolan Research Institute, Royal Free Campus, London, U.K. High- and low-resolution typing was conducted using PCR-sequence specific oligonucleotides (PCR-SSO) probes, and detected with a Luminex-based system (One Lambda LABtype SSO VH Bio, Gateshead, U.K.). The presence of the HLA*A02 allele in three healthy controls was assessed by immunostaining of PBMCs with an antibody against HLA-A2 PE, clone BB7.2 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA).
Peptide-MHC binding and dissociation assays
Ninety-four overlapping nine-mer peptides (offset by one amino acid) were generated spanning the entire KSHV ORF28 protein sequence using PEPscreen® custom peptide library synthesis. Peptide sequences are shown in Supplementary Table 2 (see online supplementary material at
IFN-γ release ELISPOT assays
Detection of IFN-γ release by ELISPOT assays was performed as previously described (5). Briefly, 96-well polyvinylidene difluoride-backed plates (Millipore, Watford, U.K.) were coated with a monoclonal antibody against IFN-γ (Mabtech, Stockholm, Sweden), and then seeded with 2 × 105 peripheral blood mononuclear cells (PBMCs) per well in RPMI-1640 media supplemented with 10% heat-inactivated human AB serum, 2 mM L-glutamine, and 1% penicillin-streptomycin (all from Sigma). Freshly isolated PBMCs were stimulated in triplicate with KSHV peptides (20 μg/mL; ProImmune Ltd.), or the following controls (all at 5 μg/mL): phytohemagglutinin (PHA; Sigma); a pool of influenza, Epstein-Barr virus (EBV), and cytomegalovirus (CMV) peptides (FEC; National Institute for Biological Standards and Control [NIBSC], Hertfordshire, U.K.); the immunodominant HLA-A*0201-restricted epitopes from HIV-1 GAG (SLYNTVATL; NIBSC); CMV pp65 (NLVPMVATV; ProImmune Ltd.); and tissue culture medium alone (media only). Spot-forming cells (SFC) were detected following the manufacturer's instructions (Mabtech), using an AP Conjugate Substrate Kit according to the manufacturer's instructions (Bio-Rad, Hemel, Hempstead, U.K.). Positive responses were designated as responses >20 SFC per million PBMCs above the background response to Tcm, and >5 times the background response. Background responses ranged from 0.0–5.0 SFC per million PBMCs.
IFNγ ELISPOTS using peptide-pulsed moDCs as stimulants
CD14-positive monocytes were isolated from whole PBMCs using labeled magnetic beads (Miltenyi Biotech, Bergisch, Gladbach, Germany), and cultured for 7 d in RPMI 1640 with HEPES modification (Sigma), supplemented with L-glutamine (Sigma), penicillin-streptomycin (Sigma), and 5% human AB serum (lot number 027K0432; Sigma). The cells were further supplemented with 75 ng/mL interleukin 4 (IL4; R&D Systems, Oxford, U.K.), and 75 ng/mL granulocyte/macrophage colony-stimulating factor (GMCSF; R&D Systems) in order to drive differentiation into immature moDCs. Cells were fed with fresh media and cytokines every second day. Immature moDCs were matured by stimulation with a “cytokine cocktail” consisting of 1 μg/mL prostaglandin E2 (PGE2; Sigma), 5 ng/mL TNF-α (R&D Systems), 5 ng/mL IL-1ß (R&D Systems), and 150 ng/mL IL-6 (R&D Systems) for 48 h. After maturation, moDCs were pulsed with 50 μg/mL peptide for 2 h at 37°C. Pulsed moDCs were then washed three times, and used as stimuli in the place of peptide solutions in an ELISPOT assay with autologous PBMCs. ELISPOTS were performed as described above.
Pentamer staining for further analysis of KSHV-specific CD8 T cells
The pentamer-specific PE fluorotag was spun in a chilled centrifuge (14,000 g) for 5 min, stored on ice, and protected from light until use. Two to three million PBMCs per staining condition were aliquotted into FACS tubes (Greiner Bio-One, Stonehouse, U.K.), washed with 2 mL BSA stain buffer (BD Biosciences, Franklin Lakes, NJ), and spun at 1000 g and 4°C for 5 min. The supernatant was discarded and the cells were resuspended in the residual liquid (∼50 μL). One test of unlabeled pentamer (1 μg for CMV-NLVPMVATV-specific pentamers, and 0.5 μg for the KSHV-specific pentamer) was added per tube, and mixed by pipetting. The cells were incubated on ice for 40 min, washed once as before, and the supernatant was discarded and the cells were resuspended in the residual liquid. Optimal amounts of monoclonal antibodies for T-cell-surface markers or isotype controls were added to the cells, along with 8 μL of the chilled fluorotag and mixed by pipetting. Antibodies used were CD3-PerCP clone SK7, and CD45RA-PECy7 clone L48 (BD Biosciences); CD8-FITC clone LT8 (ProImmune Ltd.); and CCR7-APC clone 150503 (R&D Systems). The cells were incubated on ice, protected from light for 20 min, washed twice as above, and resuspended in 300 μL in stabilizing fixative (BD Biosciences) before analysis on an LSRII flow cytometer (BD Biosciences). A minimum of 100,000 CD8 events were acquired for each sample, and data were analyzed using FlowJo software (Tree Star Inc., San Francisco, CA). The live lymphocyte gate was set according to expected forward scatter and side scatter. Isotype controls were used to set gates for positive staining for each of the antibodies. A sample labeled with the fluorotag alone without a pentamer was used as a negative control.
Results
Identification of candidate epitopes from within KSHV ORF28
Ninety-four overlapping nine-mer peptides (offset by one amino-acid residue) were synthesized spanning the entire amino acid sequence of ORF28, and numbered P1 to P94 sequentially. The ability of each peptide to bind to HLA-A*0201 was measured and compared to the binding affinity of a known high-affinity A*0201-restricted T-cell epitope (the human influenza matrix protein epitope GILGFVFTL [HI-GIL]), in order to identify putative immunogenic peptides. One of the best documented KSHV HLA-A*0201-restricted epitopes, LMWYELSKI (aa492-500 from glycoprotein B/ORF8 [ORF8-LMW]; indicated in Table 1), was included in the assay for comparison, and was designated P95. A known T-cell epitope of marginal binding was also included as an intermediate-affinity control. Unfortunately, the sequence information for the intermediate control could not be provided to us by the manufacturers (ProImmune Ltd.). Results are shown in Fig. 1 and Supplementary Table 3 (see online supplementary material at
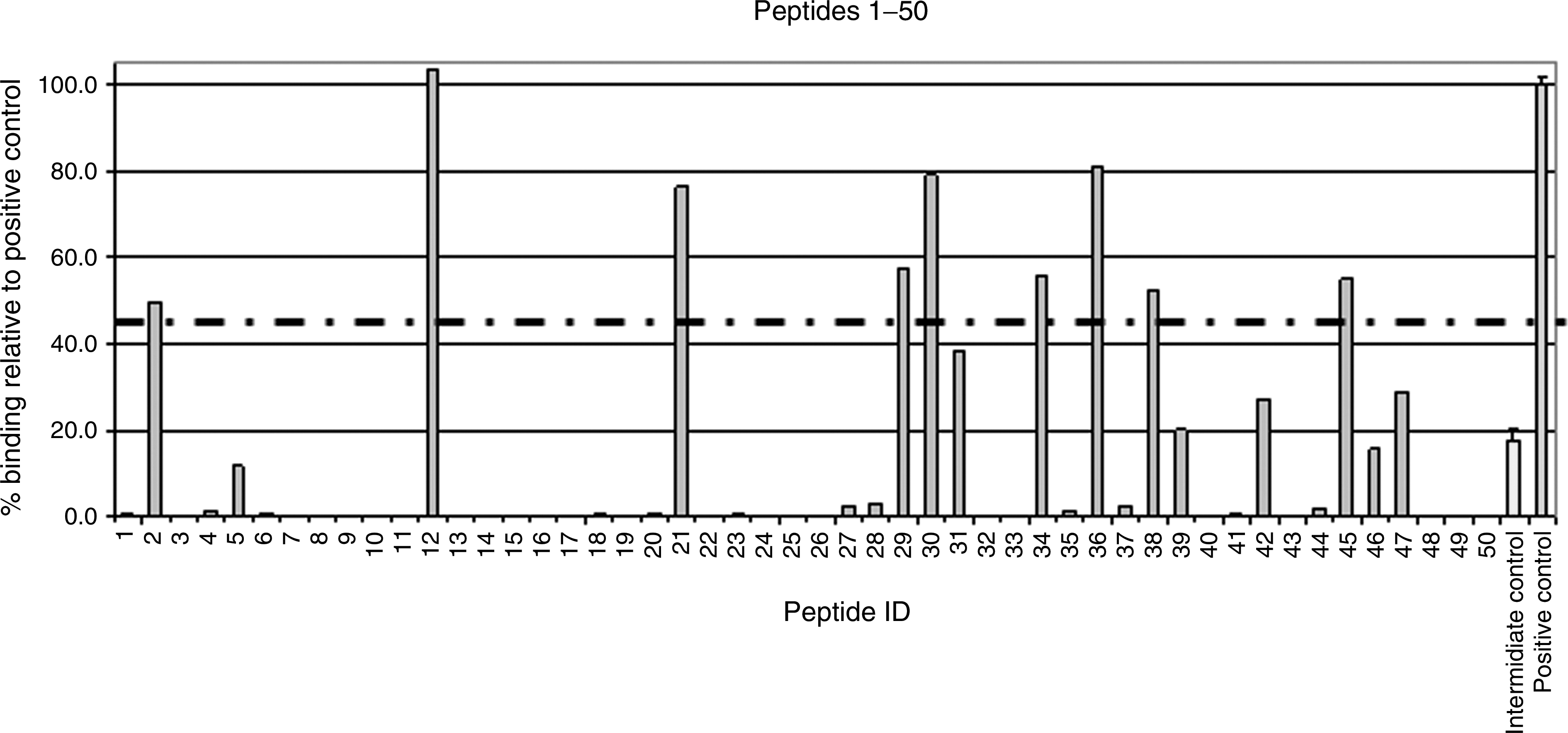
Binding affinity of ORF28-derived peptides to HLA-A*0201. Binding is shown as a percentage relative to the binding of the known high-affinity HLA-A*0201-restricted epitope, GILGFVFTL (from the influenza matrix protein; “positive control”; shown on the far right on both graphs). Dashed line indicates the cutoff score of 45%, designated to identify the best binders for use in ex vivo assays. The intermediate-affinity control epitope is shown second from right on both graphs and the KSHV control epitope (ORF8-LMW, P95) is third from the right on the lower graph.
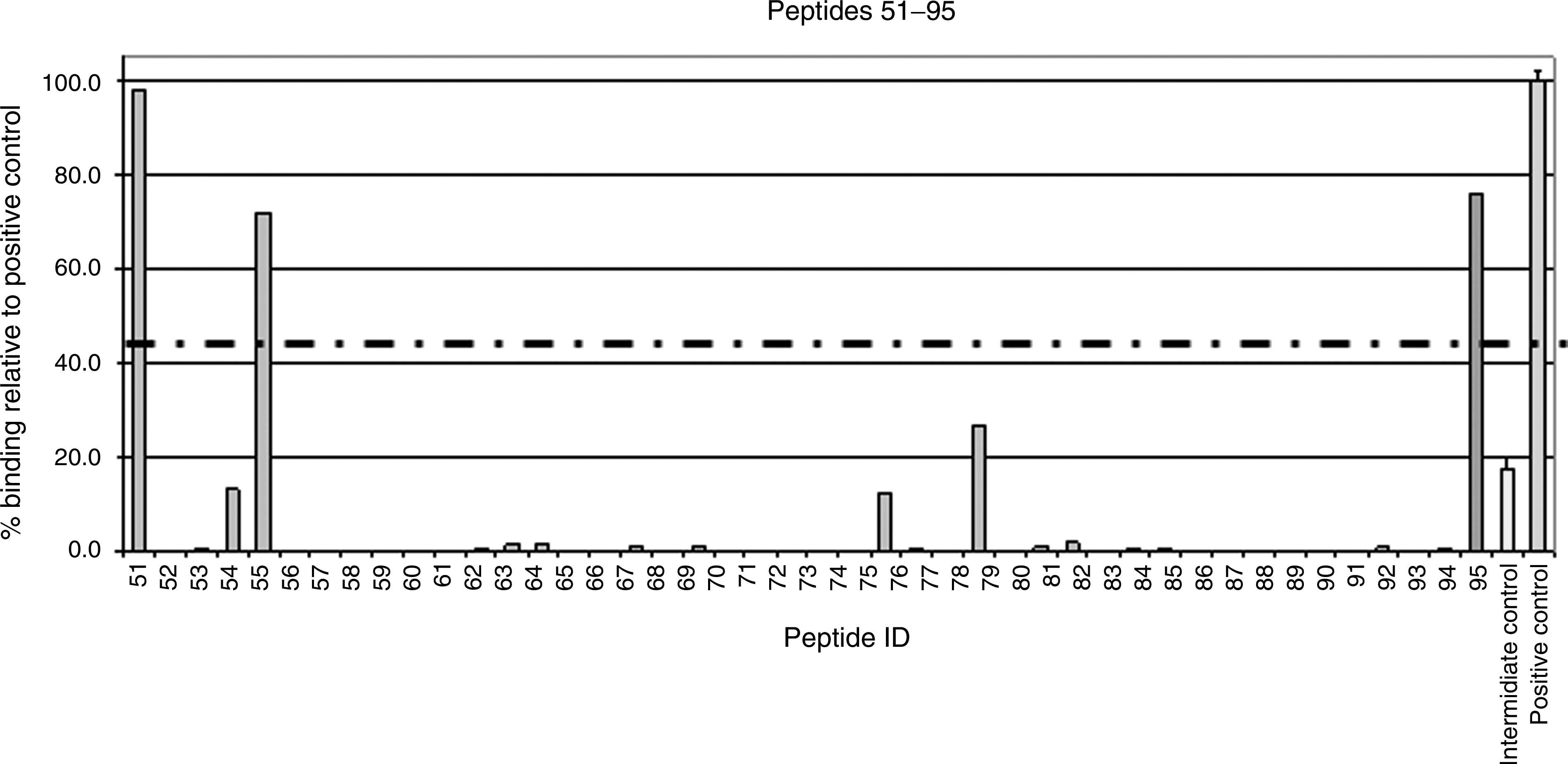
Each peptide was given a binding score as a percentage relative to the binding of the high-affinity positive control, HI-GIL. A cutoff score of 45% was selected in order to identify the best peptide binders to prioritize in ex vivo experiments. Twelve KSHV peptides (P12, P51, P36, P30, P21, P95, P55, P29, P34, P45, P38, and P2) had binding scores above this cutoff rate (103.49%, 97.95%, 81.2%, 79.09%, 76.27%, 76.25%, 71.89%, 57.52%, 55.69%, 55.20%, 52.35%, and 49.80%, respectively). These included the KSHV control epitope, P95 (ORF8-LMW; binding score 76.25%), indicating that the results of the assay concur with the results from previous ex vivo studies used to identify KSHV epitopes.
For the 12 best peptide binders, the denaturation rates for each peptide-A*0201 complex was also measured to assess how long each individual peptide could be presented to T cells, to help identify putative epitopes. The rate of dissociation of A*0201-peptide complexes was measured at 0 h, 2 h, and 24 h at 37°C (Supplementary Figure 1; see online supplementary material at
Ex vivo comparison of T-cell responses to ORF28-derived and known KSHV CD8 peptides by IFN-γ ELISPOT
The 11 ORF28-derived peptides identified as potential HLA-A*0201-restricted epitopes above, and nine previously-described HLA-A*0201-restricted KSHV CD8 epitopes (including the KSHV control peptide used in the binding assays, ORF8-LMW; indicated in Table 1), were tested for recognition by T cells in ex vivo IFN-γ ELISPOT assays (Fig. 2).
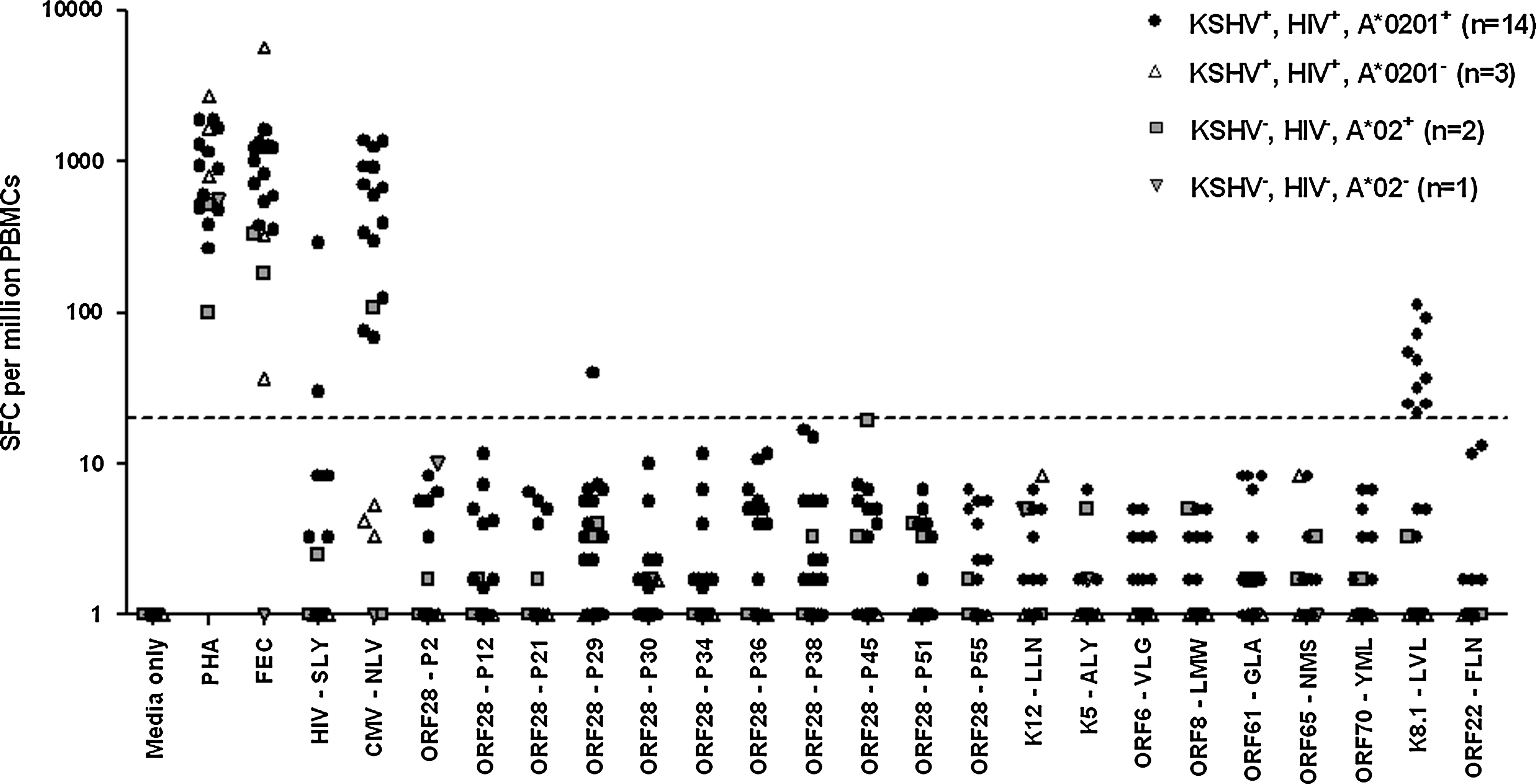
CD8 T-cell responses to A*0201-restricted peptides by HIV-1-seropositive, KSHV-seropositive individuals. Filled circles indicate 14 HLA*0201-positive, HIV-1-seropositive, KSHV-seropositive individuals. Open triangles indicate three HLA*0201-negative, HIV-1-seropositive, KSHV-seropositive individuals. Gray squares indicate two HIV-1 seronegative KSHV-seronegative HLA*2-positive individuals and gray triangles indicate one HIV-1 seronegative KSHV seronegative HLA*2-negative individual. Responses shown as SFC per million PBMCs with background response to T-cell media alone (media only) subtracted. Responses below 1.0 were set to 1.0 to enable plotting of data on a logarithmic scale. The dashed line indicates the positive response cut-off value of 20 SFC per million PBMCs. One KSHV-seropositive, HIV-1-seropositive, HLA*0201-negative individual was not tested for recognition of any of the new KSHV peptides (ORF28 P2 to P55) due to insufficient numbers of PBMCs.
All 14 HIV-1-seropositive, KSHV-seropositive, A*0201-positive individuals responded to the controls PHA, FEC, and CMV (also known as human herpesvirus 5)-NLVPMVATV (CMV-NLV). Two out of 13 tested responded to HIV-1-SLYNTVATL (HIV-SLY; 30.0 and 289.2 SFC per million PBMCs), a frequency of recognition similar to that observed by others in cohorts of HIV-1-seropositive individuals receiving HAART (13). Ten of the 14 individuals responded to one of the KSHV peptides: the previously identified peptide LVLILYLCV, derived from the late-lytic ORF K8.1 that encodes glycoprotein 35/37 (K8.1-LVL). Responses to K8.1-LVL were weak (21.7–113.3 SFC per million; mean = 52.7; median = 42.8), especially compared to responses to CMV-NLV by these 14 individuals, which ranged from 68.3–1383.3 SFC per million (mean = 653.4; median = 635.3). Nonetheless, these data indicate that K8.1-LVL represents an immunodominant HLA-A*0201-restricted KSHV CD8 epitope, as it was recognized here by T cells from 71.4% of HIV-1-seropositive, KSHV-seropositive, HLA-A*0201-positive individuals. The one individual who had active KS at venisection did not respond to K8.1-LVL, which may further indicate a protective effect for CD8 T cells specific for this peptide in the control of KSHV infection.
One out of the 14 individuals (T107) responded to one of the novel late-lytic KSHV epitopes, ORF28-P29 (40.0 SFC per million). Thus the frequency of recognition of peptide ORF28-P29 was 7.1% in this cohort of HIV-1-seropositive, KSHV-seropositive, HLA*0201-positive individuals. This was the only other KSHV epitope to be recognized by any KSHV-seropositive individuals, including eight out of the nine previously-identified KSHV epitopes.
Three HIV-1-seropositive, KSHV-seropositive, A*0201-negative participants tested responded to PHA and FEC, but not to the A*0201-restricted peptide epitopes from HIV-1 and CMV, as would be expected. None of these three individuals responded to K8.1-LVL, indicating that this epitope may indeed be A*0201-restricted.
Three HIV-1-seronegative, KSHV-seronegative Caucasian individuals were tested for responses to all peptides to ensure that there was no cross-reactivity of peptides with other antigens. The exact HLA-subtypes of these individuals were unknown, although two were A*02-positive as assessed by immunostaining with HLA-A2 PE clone BB7.2 antibody. All three of these individuals responded to PHA, and two responded to FEC. One A*02-positive individual responded to CMV-NLV (108.3 SFC per million PBMCs), confirming that they were of the A*0201-subtype. No responses were seen to any of the KSHV peptides by T cells from any of these three individuals.
Enhancing responses to KSHV peptides by using peptide-pulsed autologous monocyte-derived dendritic cells (moDCs) as stimulants in ELISPOTS
The results presented above highlight the difficulties associated with investigating weak KSHV-specific T-cell responses to peptides using such IFN-γ ELISPOT assays, particularly in cohorts of HIV-1-seropositive individuals. Others have used autologous moDCs pulsed with KSHV peptides as stimulants in place of peptide solutions to facilitate detection of elusive weak responses (19). We therefore used this technique with a selection of peptides (ORF28-P36, ORF28-P51, ORF8-LMW [P95 in the binding assays], and K8.1-LVL) to see if any additional responses could be detected. ORF28-P36 and ORF28-P51 were selected, as these were two of the highest-scoring ORF28 peptides in the MHC binding and off-rate assays. ORF8-LMW was included as the best-documented A*0201-restricted KSHV epitope, and also as others have detected responses to this peptide using peptide-pulsed moDCs in a similar protocol (19). K8.1-LVL was selected to examine whether weak responses observed to this peptide in solution could be replicated and enhanced using this protocol. Fig. 3 shows results from six individuals for whom sufficient PBMCs were available for the generation of moDCs (D850, J228, M907, S314, T514, and healthy control [HC]-A).
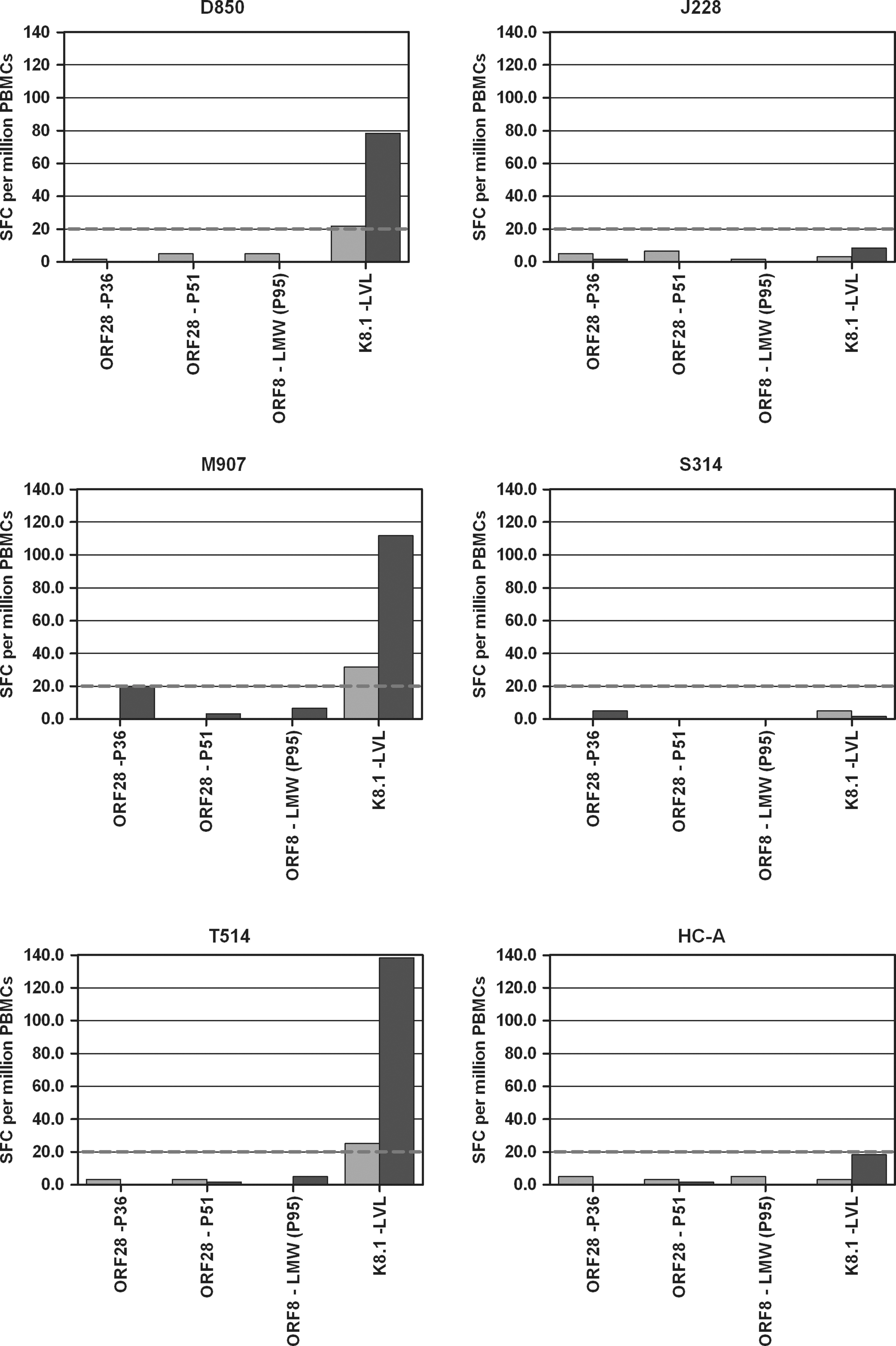
CD8 T-cell responses to peptide-pulsed autologous moDCs. Bars indicate T-cell responses to HLA-A*0201-restricted peptides in IFN-γ ELISPOT assays using peptide solutions (pale gray bars), or peptide-pulsed autologous moDCs (dark gray bars) as stimulants. Responses shown are SFC per million PBMCs after the background response to T-cell media alone (peptide solution assays), or moDCs alone (peptide-pulsed moDC assays) had been subtracted. The dashed line indicates the positive cut-off value of 20 SFC per million PBMCs.
Three of the six individuals (D850, M907, and T514) gave weak positive responses to K8.1-LVL in peptide-stimulated ELISPOTS (21.7, 31.7, and 25.0 SFC per million PBMCs, respectively). Using peptide-pulsed autologous moDCs as stimulants enhanced these responses to 78.3, 111.7, and 138.3 SFC per million PBMCs, respectively. The other three individuals (J228, S314, and HC-A) did not respond to K8.1-LVL in the peptide-stimulated assay, and did not respond to autologous moDCs pulsed with K8.1-LVL. Likewise, none of the six individuals responded to any of the other peptides (ORF28-P36, ORF28-P51, and ORF8-LMW) in the peptide-stimulated assay, and did not respond to moDCs pulsed with these peptides. This gives confidence that our techniques did not generate false-positive responses, and confirms the validity of the low-positive responses (for example, those below 50 SFC per million PBMCs) observed in peptide-stimulated assays. A limitation of this technique is, however, that far fewer peptides can be investigated in this manner, due to the number of PBMCs needed to generate sufficient numbers of moDCs for use with each different peptide.
Memory phenotypes of KSHV-specific CD8 T cells
A goal of these studies was to identify immunodominant epitopes (either one of several previously described epitopes or a novel ORF28-derived epitope) that would be a suitable target for investigations utilizing multimers to enumerate KSHV-specific T cells, and to explore their memory phenotypes and functional properties. The study identified one clear immunodominant epitope, LVLILYLCV, from the late-lytic ORF K8.1. Disappointingly, this peptide was too hydrophobic for multimer synthesis. One other KSHV-derived peptide (ORF28-P29) elicited a response in a single individual (T107). Synthesis of the corresponding multimer was successful and the multimer was used to enumerate KSHV-specific cells in the responsive participant, and to determine the memory phenotype of KSHV-specific CD8 T cells in conjunction with antibodies against the T-cell differentiation markers CCR7 and CD45RA (23). A pentamer specific for the immunodominant HLA-A*0201-restricted CMV peptide CMV-NLV was used as a control (Fig. 4).
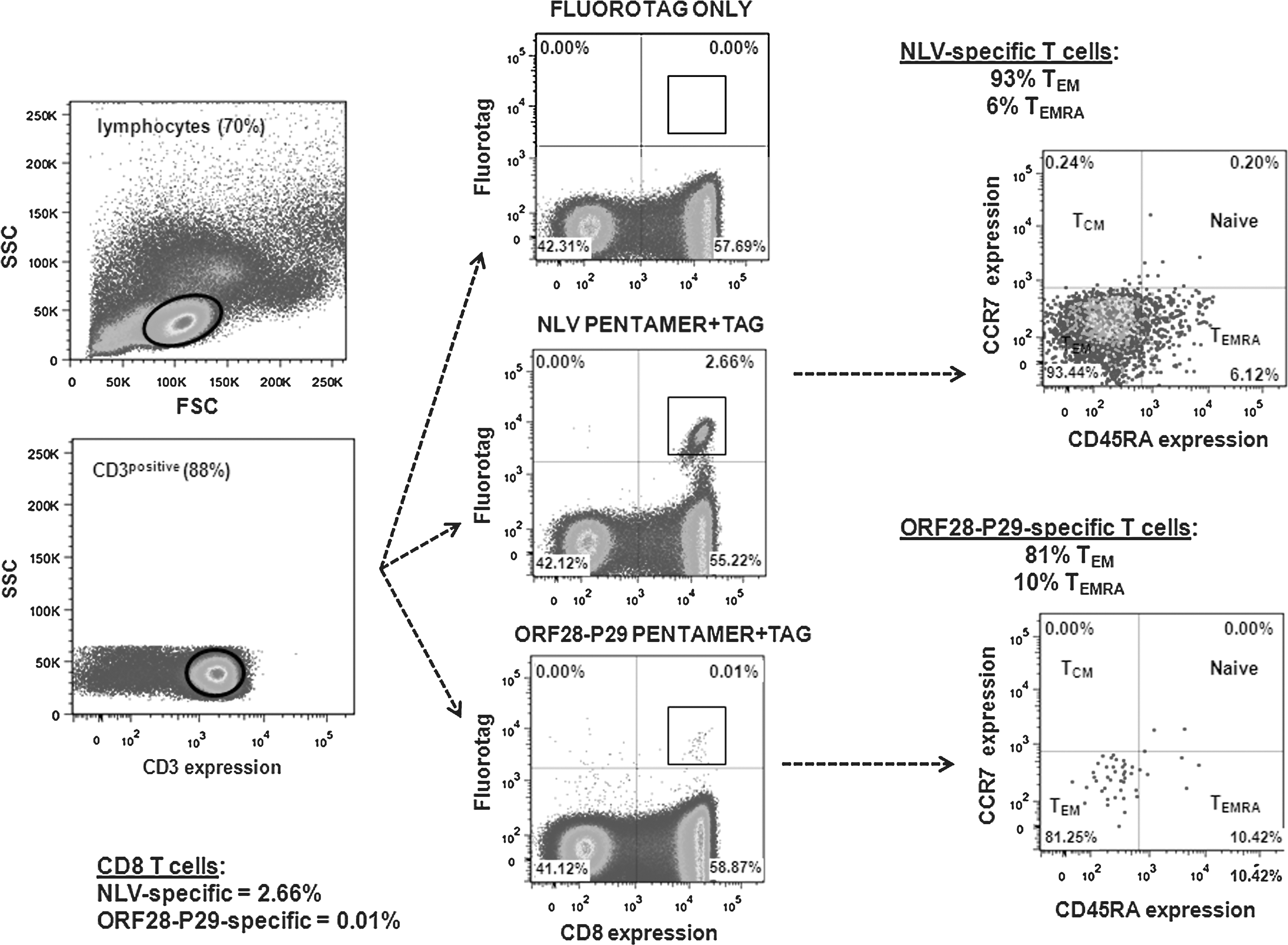
Memory phenotypes of CD8 T cells specific for the novel KSHV epitope ORF28-P29. Results are shown from one individual, T107, with comparison to memory phenotype of CD8 T cells specific for the immunodominant HLA-A*0201-restricted CMV epitope, CMV-NLVPMVATV, from the same individual. CD8 T-cell memory subpopulations were identified by immunostaining with antibodies against the lymphoid homing marker CCR7 and the “RA” isoform of CD45.
In this individual, the proportion of CMV-NLV-specific T cells was unusually high (2.66% of total CD8 T cells), and these cells were highly skewed towards a CCR7-CD45RA-effector memory (TEM) phenotype (93.44% TEM cells and 6.12% CCR7-CD45RA+ terminally-differentiated effector memory [TEMRA] cells). KSHV-specific T cells against ORF28-P29 were much lower in frequency (0.01%), in accord with the data from the ELISPOTS (Fig. 2). The KSHV-specific T cells were also skewed towards a TEM phenotype (81.25% TEM cells and 10.42% TEMRA cells).
To confirm specificity of pentamer staining, the ORF28-P29 pentamer was also tested in an A*0201-positive, KSHV-seronegative individual, as well as an A*0201-positive, KSHV-seropositive individual who did not respond to the ORF28-P29 peptide in an ELISPOT assay. No pentamer staining was observed in either individual (Supplementary Figure 3; see online supplementary material at
Discussion
MHC binding assays were used to identify potential new HLA-A*0201-restricted epitopes from within the late-lytic KSHV glycoprotein encoded by ORF28. These were then tested for recognition by T cells from 14 HIV-1-seropositive, KSHV-seropositive, HLA-A*0201-positive individuals, alongside nine previously identified A*0201-restricted KSHV epitopes from ORFs expressed across the spectrum of the KSHV replication cycle.
Only one of the candidate A*0201-restricted epitopes identified from within ORF28 was recognized by T cells from any of the 14 individuals tested (epitope ORF28-P29, recognized by one participant). Frequency of recognition of this epitope was low (7.1% of individuals). Nonetheless, this epitope represents a novel KSHV epitope derived from the late-lytic glycoprotein ORF28, and comprised of aa29-37 (IGLITVLFL). Furthermore eight of the nine previously identified KSHV epitopes were not recognized by any of the 14 KSHV-seropositive individuals, and so the new ORF28-IGLITVLFL epitope may represent an epitope of equal, if not greater, biological importance than many of the currently identified epitopes.
Unfortunately, a limitation of the present study was that we were only able, in this instance, to focus on nine-mer peptides and their binding affinity to A*0201. A wider study of peptides of different lengths and their binding affinity to alternative MHC-I complexes may yet reveal more frequently recognized epitopes from within ORF28.
The lack of responses to eight of the nine previously identified epitopes was surprising, as reports in the literature indicated that these eight epitopes were recognized by 16–100% of all individuals tested (total of HIV-positive and HIV-negative), or 13–32% of HIV-positive individuals tested in previous studies (Table 1). This disparity may arise from differences between previous studies and the present study in both the experimental approaches used to detect T-cell responses, and in the patient cohorts in which responses were investigated. In particular, many of the previous studies used long culture techniques to expand memory T cells before detection (7,15,17,22). The present study aimed to make the first direct comparison of ex vivo responses to a wide selection of epitopes in a well-defined cohort of KSHV-seropositive individuals who were also HIV-1-coinfected and had a history of KSHV-related neoplasia. This cohort may have disadvantages with respect to eliciting suboptimal KSHV-specific T-cell responses. Nonetheless, it could be argued that with a long-term goal of developing better treatments for, or a prophylaxis against, symptomatic KSHV infection, investigations in such a cohort of individuals remain the most relevant.
Strikingly, in this cohort, one peptide, K8.1-LVLILYLCV, from the late-lytic, glycoprotein-encoding ORF K8.1, did elicit a response by T cells from 10 of the 14 individuals tested (71.4%) in ex vivo assays. Although the responses we observed were weak, we found that they could be enhanced by using peptide-pulsed autologous moDCs in place of peptide solutions as stimulants in ELISPOT assays. These findings concur with the original data collected by our group (5), which indicated that K8.1-LVL could be the most frequently recognized of all known A*0201-restricted KSHV epitopes, with responses seen in 50% of individuals tested by ex vivo ELISPOT (5) (Table 1). The limitations of this original study were a small sample number (eight individuals), and a lack of comparison to responses to other known KSHV CD8 epitopes by the same individuals. The new data described in the present study thus provide further evidence for K8.1-LVL representing an immunodominant A*0201-restricted KSHV epitope, arguably the first such epitope to be described.
The identification of K8.1-LVL as an immunodominant peptide, and in particular, the finding that out of nine of the best described known KSHV CD8 epitopes, K8.1-LVL was the only one that could be readily detected ex vivo in our cohort of KSHV-seropositive, HIV-seropositive individuals, may have implications for the design of future immunological studies of KSHV infection. It would, for example, be a useful target in experiments tracking changes in the KSHV CD8 T-cell response during relapsing-remitting KS. This information may also be useful in the future in the potential development of immunotherapies for KSHV-related malignancies, or even a vaccine against KSHV. It may also be useful to researchers in the field to note the difficulties we experienced in detecting responses to any of the other previously identified KSHV CD8 epitopes.
Unfortunately, K8.1-LVLILYLCV proved to be too hydrophobic for synthesis of a multimer that would enable fluorescence labeling of KSHV-specific T cells. Specifically, the peptide was too hydrophobic to remain in solution under the conditions used for the artificial assembly of MHC chains around the peptide during pentamer synthesis. Nonetheless, there is evidence that similarly hydrophobic peptides are presented in vivo during the human immune response to other viruses. Highly hydrophobic HLA-A2-restricted peptides have been identified in the EBV proteins BNLF2a (VLFGLLCLL), and LMP2 (LLWTLVVLL and CLGGLLTMV) (2,14). These peptides are efficiently presented in the context of HLA-A2 (2), and they appear to be processed for MHC-I presentation via a TAP-independent pathway, although the precise mechanism of presentation has not been determined (14).
However, the novel epitope (ORF28-P29) identified by this study was suitable for multimer synthesis. The phenotype of T cells specific for this peptide was investigated in the one responder identified in this study. ORF28-P29-specific T cells were predominantly of a TEM phenotype (81.25%), with few cells showing a TEMRA phenotype. It would not be rational to attempt to draw conclusions from this single experiment, and it is especially worth noting that in this individual, CMV-NLV-specific T cells were unusually highly skewed towards a TEM phenotype (93.44%). Nonetheless, the observed phenotype of KSHV-specific T cells from this one individual is in keeping with findings by others (3). Further experiments to identify additional ex vivo responders to this epitope, and to use multimers to examine the phenotype of these cells are warranted.
Footnotes
Acknowledgments
This work was supported by funding from the Medical Research Council UK, the Westminster Medical School Research Trust, the Westminster Healthcare Charities, and the Skin Treatment and Research Trust (START). The authors wish to thank study participants and staff at the Chelsea and Westminster Hospital who contributed to this study, and Professor Steven Marsh and Finnuala Fowles from the Anthony Nolan Research Institute for their help with the HLA typing of study participants.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.