
Abstract
The pandemic H1N1 2009 influenza A emerged in April 2009 and spread rapidly all over the world. In Greece, the first case of the pandemic H1N1 was reported on May 18, 2009, while a considerable increase in the number of cases was noticed at the beginning of July 2009. The need for surveillance of the immune status of the Greek population led us to develop a virus-free ELISA that specifically recognizes pandemic H1N1 2009 influenza virus antibodies in human sera. The method is based on the use of synthetic peptides (H1-pep and N1-pep) that are derived from the hemagglutinin and neuraminidase of the 2009 pandemic strain, respectively, and differentiate the swine-origin influenza A/California/14/2009 (H1N1) from the seasonal influenza A viruses. Serum samples were obtained from 271 healthy blood donors during May, November, and December 2009. Among sera collected during May, November, and December, IgG antibodies against the peptide H1-pep were detected in 7.4, 13.8, and 19.3% of the donors, respectively, while IgG antibodies against the peptide N1-pep were detected in 5.3, 9.6, and 16.9% of the donors, respectively. The application of the immunoassay indicated a time-dependent increase of the prevalence of anti-H1-pep and anti-N1-pep IgG antibodies during the pandemic H1N1 outbreak in Greece. The method could be also indicative for the discrimination of immune persons from those susceptible to infection with the pandemic H1N1 strain, as well as for the establishment of effective vaccination programs.
Introduction
The novel H1N1 influenza strain, which caused the new pandemic, was first recognized at the border between Mexico and the United States on April 9, 2009; it has undergone triple reassortment since it contains genes from avian, swine, and human influenza viruses (19). Through rapid and frequent international travel, it has spread to over 74 countries around the world and has been implicated in 29,000 cases up to June 12, 2009, when it was declared as the first pandemic of the 21st century (6).
In Greece the first laboratory-confirmed case of the pandemic H1N1 2009 influenza was reported on May 18, 2009 and was detected in a 19-year-old male who had returned from New York City 2 d earlier. On 26 and 27 May, the second and the third cases were detected in two students returning from the United Kingdom (20). During July and August, Greece experienced a moderate wave of transmission of 2009 H1N1, which may have reflected the increased frequency of tourist visits to Greece during this period (15).
The total number of people infected around the world is difficult to estimate. By the end of the year 2009, the WHO had declared that 120,000 persons had died due to influenza A/H1N1, half of them in North America (Mexico, Canada, and the U.S.) (2). On the other hand, a significant fraction of the world population may already have been exposed to the virus and although asymptomatic, could be at least partially immune to the disease (1,17,22).
The diagnosis of infection with 2009 influenza virus can be carried out by direct detection of the virus, which is accomplished by isolation of virion particles, viral antigen detection through immunospecific assays and targeting of viral-specific nucleic acids in respiratory specimens, or by detection of antibodies specific for the virus in serum samples (8,12,14,18,24,25). The hemagglutination inhibition (HI) test is considered the gold standard method for the detection of antibodies against influenza virus, while the microneutralization method, which measures the reduction in virus growth due to neutralization by antibodies, has also been developed (5,11). Recently, an ELISA method was developed based on the use of a recombinant fragment of HA of influenza A H1N1/2009 virus expressed in Escherichia coli that evaluates the presence and relative concentration of anti-influenza A antibodies in human serum samples (2).
In the present study, we documented the development and use of an ELISA method that specifically recognizes 2009 H1N1 influenza virus antibodies in human serum samples, based on the use of synthetic peptides derived from specific amino acid sequences on HA and NA of the novel pandemic H1N1 influenza virus.
Materials and Methods
Collection of sera
Serum samples were obtained from 271 healthy blood donors. Ninety-four of these sera were collected during May 2009, 94 during November 2009, and 83 sera during December 2009, from blood transfusion centers in Greece. Since vaccination against 2009 pandemic influenza A (H1N1) in Greece was initiated at the end of November 2009, blood donors who had already received the 2009 H1N1 vaccine were excluded from the study. All sera were tested for the presence of IgG antibodies by ELISA that uses two specific antigens: (1) a synthetic peptide derived from HA-H1N1 swine-origin influenza A/California/14/2009 (H1-pep), and (2) a synthetic peptide derived from NA-H1N1 swine-origin influenza A/California/14/2009 (N1-pep).
Synthetic peptides
The synthetic peptides (R01419 and R01421), purchased from Meridian Life Science, Inc. (Memphis, TN), include sequences from H1 (H1-pep) and N1 (N1-pep) antigens, that differentiate the swine-origin influenza A/California/14/2009 (H1N1) from the seasonal A H1N1 viruses. Synthetic H1 and N1 peptides derived from the seasonal A/Brisbane/59/2007 (H1N1) and the seasonal influenza A/Georgia/20/2006 (H1N1), respectively, were also purchased. These peptides were also used for the immunization of rabbits and the production of antibodies capable of recognizing these different amino acid sequences. The rabbit antisera were used for estimation of the efficiency of the ELISA method and also as positive controls.
ELISA for the detection of antibodies specific to hemagglutinin and neuraminidase of H1N1 swine-origin influenza A/California/14/2009
Preliminary ELISA experiments were performed in order to define the optimal conditions. The optimal peptide concentration for coating was determined to be 1 μg/mL for each peptide. ELISA plates (Costar; Corning Life Sciences, Acton, MA) were coated for 3 h at room temperature with the peptides diluted in carbonate-bicarbonate buffer (pH 9.6) at their optimal concentrations. Afterward, the plates were blocked with 200 μL of bovine serum albumin (BSA) in 2% in PBS (pH 7.2) for 2 h at room temperature. Then the plates were incubated for 2 h at room temperature with human serum diluted 1:100 in blocking buffer. After three washes with PBS, alkaline phosphatase-conjugated anti-human IgG (Jackson ImmunoResearch, West Grove, PA; diluted 1:1100 in blocking buffer) was added in order to detect antibodies bound onto the solid phase. After an incubation period of 1 h at room temperature, the plates were washed three times with PBS, and the enzyme reaction was developed with p-nitrophenyl phosphate substrate (Sigma-Aldrich, St. Louis, MO). The optical densities (ODs) were quantified at 405 nm with an ELISA reader ChroMate™ 4300 (Awareness Technology Inc., Palm City, FL). Specific rabbit antisera that recognize H1-pep and N1-pep peptides (Meridian Life Science) were used as positive controls and for normalization of readings among different ELISA plates. All ODs were transformed and expressed as binding units according to the following formula: binding units (BU) = (ODSample/ODPosCtlr) × 100, where ODSample is the OD reading of the current sample, and ODPosCtlr is the mean OD of the three positive controls in the current ELISA plate. The cutoff value for anti-peptide ELISA was calculated as mean normal serum binding units plus 3 standard deviations. Samples were considered positive when the corrected OD was above the cutoff.
Statistical analysis
Using the χ2 test, statistical comparisons between seroprevalence rates were performed. Correlation between the reactivity against H1-pep and N1-pep was achieved using Pearson r.
Results
Evaluation of the potential of synthetic peptides to discriminate between antibodies directed against seasonal influenza A H1N1 and swine-origin 2009 influenza A H1N1
The efficiency of the ELISA assay developed in our study as well as its ability to discriminate between antibodies recognizing seasonal influenza H1N1 and pandemic 2009 influenza H1N1 virus were evaluated using polyclonal anti-peptide antibodies that were produced in rabbits immunized with either H1-pep or N1-pep from seasonal and pandemic 2009 H1N1 viruses. The polyclonal antibodies recognized specifically their immunizing peptide, producing a minimal cross-reaction with the homologous peptide. In this regard, antibodies produced after immunization with seasonal influenza A virus H1-pep or N1-pep recognized specifically the corresponding peptides derived from seasonal H1N1, but not the homologous peptides derived from swine-origin 2009 H1N1 and vice versa (Fig. 1).

Antibodies against either H1-pep or N1-pep from seasonal H1N1 and swine-origin 2009 H1N1 recognize specifically their target peptide without producing cross-reactions with the other peptides.
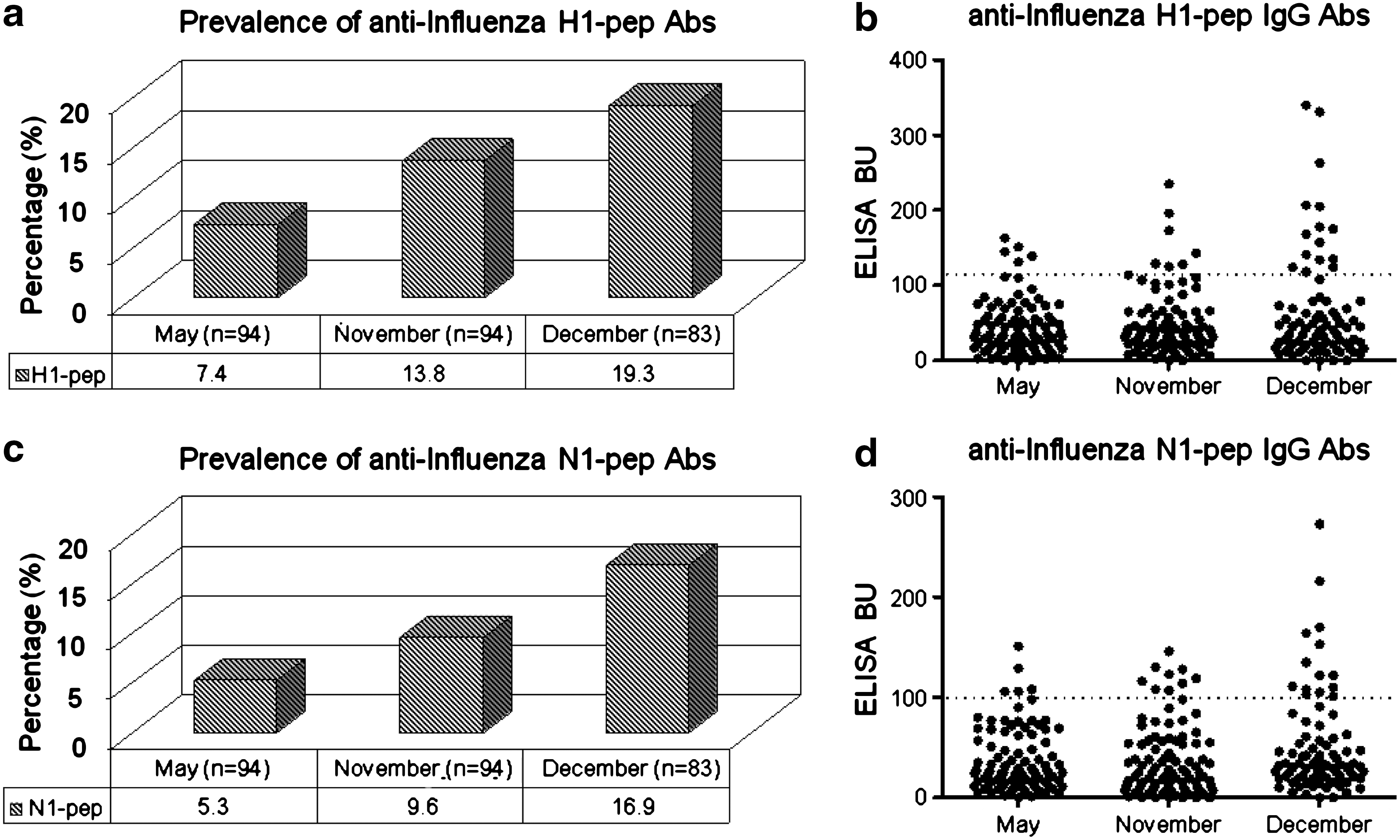
Prevalence of IgG antibodies against H1N1 swine-origin influenza A/California/14/2009 in sera collected during the H1N1 outbreak
IgG antibodies against the synthetic peptide H1-pep, derived from HA-H1N1 swine-origin influenza A/California/14/2009, were detected in 7.4, 13.8, and 19.3% of the serum samples collected during May, November, and December 2009, respectively (Fig. 2a and 2b). Similarly, IgG antibodies against the synthetic peptide N1-pep derived from NA-H1N1 swine-origin influenza A/California/14/2009 were found in 5.3, 9.6, and 16.9% of serum samples collected during May, November, and December 2009, respectively (Fig. 2c and 2d). The highest prevalence of IgG antibodies against H1-pep was detected in blood donors from December (χ2 = 4.460, p < 0.05). Similarly, the prevalence of IgG antibodies against N1-pep was found to be significantly higher in blood donors in December (χ2 = 5.122, p < 0.05). These data suggest an increase in the prevalence of IgG antibodies against HA and NA during the 2009 H1N1 influenza outbreak.
(
Correlation of IgG reactivity against H1-pep and N1-pep peptides during the course of the outbreak
The quantitative analysis of the sera binding revealed that their reactivities against H1-pep were statistically correlated with those against N1-pep for every month the serum samples were collected (May 2009, Pearson r = 0.8501, p < 0.0001; November 2009, Pearson r = 0.7663, p < 0.0001; December 2009, Pearson r = 0.7287, p < 0.0001; Fig. 3). The blood donors bearing antibodies against H1-pep also exhibited anti-N1-pep reactivity, and the application of Pearson r confirmed the positive correlation.
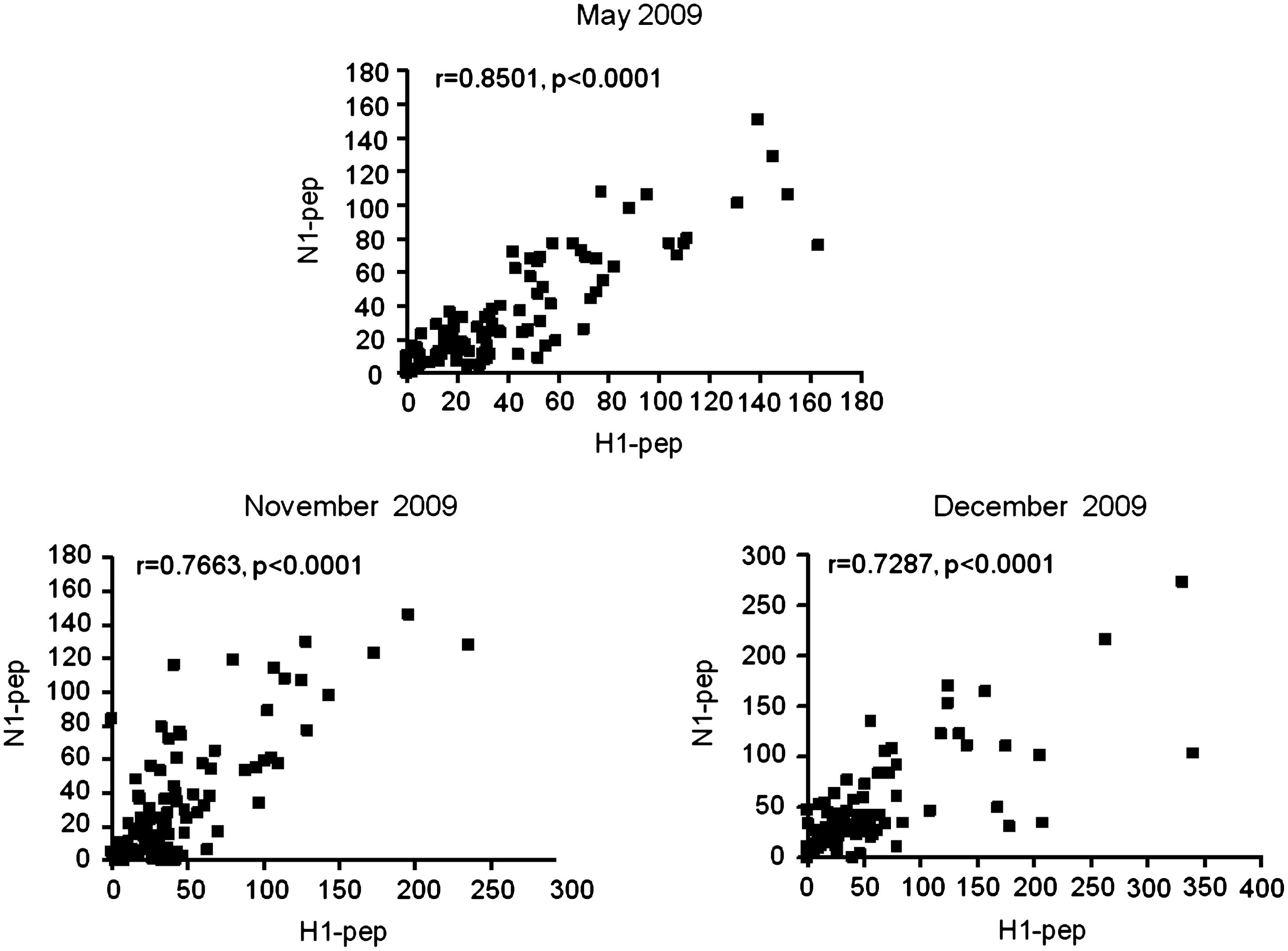
Correlation between anti-H1-pep and anti-N1-pep reactivities, using Pearson r, during the three time periods of the study.
Discussion
In the face of a pandemic influenza, the application of the typical HI assay for the detection of immune status against the pandemic influenza virus is limited due to some disadvantages. Typically, the HI assay comprises the universal protocol for estimating antibody titers against a particular influenza strain based on the ability of a serum sample to inhibit hemagglutination. However, HI titers only indicate interference of a proportion of anti-HA antibodies with sialicated receptors at the erythrocyte membrane (2). Moreover, HI assays require the use of viruses. As far as epidemiological studies are concerned, the analysis of a great number of samples is typical (1,11,17,22). The completion of these studies using an HI assay requires an abundant supply of virus and fresh chicken or turkey erythrocytes, while in case of new influenza strains, level 2 or 3 bio-safety laboratory spaces and specially-trained personnel are needed.
In the present work, we present an ELISA method that allows the determination of titers of antibodies specifically directed against the swine-origin influenza A/California/14/2009 (H1N1). The immunoassay is based on the use of synthetic peptides corresponding to amino acid sequences, which are identified on HA and NA of H1N1 swine-origin influenza A/California/14/2009.
In comparison with conventional HI tests, the immunoassay proposed here has significant advantages. The present assay does not require the use of viral particles and does not depend on the use of erythrocytes, therefore it could be easily reproduced in clinical laboratory settings by personnel with typical laboratory skills. Additionally, the interpretation of results relies on absorbance readings instead of subjective visual estimations of agglutination. Consequently, the protocol presented here would facilitate the generation of reliable results during epidemiological surveys of the immune status against the pandemic influenza strain.
Our data suggest a time-dependent increase of the prevalence of subjects positive for H1-pep and N1-pep peptides during the course of the H1N1 influenza outbreak in Greece. The positivity of sera against N1-pep was confirmed by the concurrent positivity of the same sera against H1-pep. We should also note that the prevalence of anti-N1-pep antibodies was found to be slightly lower than the prevalence of anti-H1-pep antibodies for each time period tested. These findings most likely could be explained by differences in the immunodominance between the two epitopes rather than the accumulation of mutations in the HA antigen of the H1N1 2009 influenza virus (16). The first case of 2009 H1N1 in Greece was reported on May 2009, approximately 4 wk after the initial reports of novel influenza cases in Mexico and the United States (9,10). In mid-October, influenza incidence accelerated rapidly, peaked at the end of November, and then declined steadily (7).
It is well-established that cross-reactive antibodies to 2009 H1N1 virus were present in a large proportion of older adults prior to the emergence of the novel virus (13). In our study, sera positive for IgG antibodies against H1-pep and N1-pep were probably derived from two groups of individuals. The first group constitutes blood donors who were previously infected with seasonal influenza A H1N1, and in this case the reactivities against H1-pep and N1-pep peptides are the result of cross-recognition of H1-pep and N1-pep antigens from seasonal and 2009 influenza A H1N1. The second group is comprised of blood donors who actually got infected with the pandemic H1N1 influenza A virus. Many people had a subclinical infection with 2009 H1N1 flu and never sought treatment, while others may have sought and received treatment, but were never officially tested or diagnosed, and as the outbreak intensified, in some cases testing was limited to only hospitalized patients. Epidemiological data in Greece indicate 18,075 laboratory-confirmed 2009 H1N1 cases, 294 ICU admissions, and 140 deaths related to 2009 H1N1 during May 18, 2009–February 28, 2010 (7).
Besides its obvious epidemiological significance, the estimation of the number of persons that have specific antibodies would only be indicative of the discrimination of immune persons from those more susceptible to infection, since the existence of antibodies against H1-pep and N1-pep does not assure that any particular person is entirely protected against H1N1 2009 viral infection. Despite the presence of pre-existing immunity in a large part of the older population (13), the majority remains susceptible to infection with the pandemic influenza strain (23). In Greece, vaccination against 2009 pandemic influenza A H1N1 was initiated at the end of November 2009, and although the goal was widespread coverage, as of February 28, 2010, only 3.2% of the Greek population had been vaccinated against 2009 H1N1 (23). The low acceptance of vaccination was primarily due to fear about the vaccine's safety and the probable side effects (21). This immunosorbent assay could serve as a useful tool for the continued surveillance of the immune status of different risk groups, and could help to establish an effective vaccination policy and discern which groups should be vaccinated first.
Footnotes
Author Disclosure Statement
No competing financial interests exist.