
Abstract
The pandemic 2009 H1N1 influenza virus broke out in North America and spread rapidly throughout the world. The type I interferon (IFN) response represents one of the first lines of defense against influenza virus infections. In this study, the protective potential of human exogenous IFN-ω against pandemic 2009 A (H1N1) influenza virus was assessed both in vitro and in guinea pigs. The viral loads of pandemic 2009 A (H1N1) influenza virus strains A/California/04/2009 and A/Beijing/501/2009 were reduced by up to 5000-fold in Caco-2 cells by the addition of human IFN-ω. With daily intranasal treatment with human IFN-ω the viral load of pandemic 2009 A (H1N1) influenza virus strain A/California/04/2009 decreased by 1000-fold in lung tissues of guinea pigs. These results provide strong support for the application of human IFN-ω pretreatment to human influenza control.
Introduction
Human omega interferon (IFN-ω), like IFN-α, is secreted from leukocytes in response to viral infection, and has antiviral, antiproliferative, and immunomodulatory activities (7,8). Human IFN-ω has 62% amino acid identity with human IFN-α, and 33% amino acid identity with human beta interferon (IFN-β). The human IFN-ω produced by CHO cells (glycosylated IFN-ω) has been shown to be a potent antiviral agent (9). As a distinct IFN, IFN-ω might be beneficial either for the treatment of patients who fail to respond to IFN-α, or as an additional treatment option.
The 2009 influenza pandemic was a global outbreak of a new strain of H1N1 influenza virus, which caused increased morbidity and replicated to higher titers in lung tissue of animal models than seasonal influenza virus (10,11). Steel et al. found that the use of IFN-α as antiviral prophylaxis could be an effective way to limit the spread of this virus in a guinea pig model (12). In this report, we therefore investigated the efficacy of nasal administration of human IFN-ω as a potential prophylactic agent against 2009 A (H1N1) influenza virus infection in a guinea pig model. We utilized recombinant human IFN-ω produced in the yeast Pichia pastoris, which is now in phase I clinical trials for the treatment of hepatitis C virus (HCV) infection in China (13). In this study, we demonstrated that recombinant human IFN-ω was effective in reducing the replication of a pandemic influenza virus strain in guinea pigs.
Materials And Methods
Cell culture and virus
Madin-Darby canine kidney (MDCK) cells and human colonic carcinoma (Caco-2) cells were maintained in Dulbecco's modified Eagle's medium (DMEM; Thermo Scientific HyClone, Logan, UT) supplemented with 10% fetal bovine serum (FBS; Thermo Scientific HyClone) and 2 mM glutamine. Both cell lines were from the cell library of the Institute of Biochemistry and Cell Biology, CAS (Shanghai, China). The influenza virus strains H1N1 A/California/04/2009 (Ca/04) and H1N1 A/Beijing/501/2009 (BJ/501) were grown in MDCK cells in DMEM with 2 μg/mL tolylsulfonyl phenylalanyl chloromethyl ketone (TPCK)-trypsin (Sigma-Aldrich, St. Louis, MO) at 35°C in a 5% CO2 incubator. BJ/501 influenza virus was isolated from a female patient with severe respiratory disease in PLA 302 Hospital in Beijing, China. The entire DNA sequence of BJ/501 influenza virus shares 99% similarity with the sequence of Ca/04 influenza virus. Viral titers were determined by limiting dilution analysis, and are expressed as 50% tissue culture infectious doses (TCID50) in MDCK cells according to the Spearman-Karber formula (14).
Immunostaining of pandemic 2009 A (H1N1) influenza virus
Caco-2 cells or MDCK cells were maintained in DMEM containing 10% FBS at 37°C in a 5% CO2 incubator. Both cells lines were grown on 6-well slide glasses and infected with Ca/04 or BJ/501 influenza virus at a multiplicity of infection (MOI) of 0.1. The slides were air dried at 48 h post-infection (hpi), then fixed with 4% paraformaldehyde for 10 min at room temperature. The slides were permeabilized in 0.5% Triton-X 100 solution for 15 min, washed twice with phosphate-buffered saline (PBS), and blocked in 1% bovine serum albumin (BSA) solution for 30 min. Then 1 μg/mL of polyclonal antibody against Ca/04 HA (Sino Biological Inc., Beijing, China) was placed on the slides under humidified conditions, and incubated at 37°C for 1 h. After washing with PBS, HRP-labeled secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA) was added to the slides and incubated for 1 h at 37°C. The 3,3′-diaminobenzidine (DAB) liquid chromogen substrate kit (Sigma-Aldrich) was used for staining procedures. After washing with PBS, the slides were examined for the staining pattern under a microscope.
In vitro protection assay by IFN-α, IFN-β, and IFN-ω
Caco-2 cells (2×104 cells per well) were plated in 96-well plates, grown overnight to obtain>80% confluence, and the medium was replaced with DMEM containing 2 μg/mL TPCK-trypsin. The cells were then primed with increasing doses of human IFN-α (0–5.0×105 IU/mL IFN-α2b; Merck, Kenilworth, NJ), human IFN-β (0–1.0×105 IU/mL Rebif®; EMD Serono, Rockland, MA), and human IFN-ω (0–1.0×105 IU/mL; Beijing Institute of Microbiology and Epidemiology, Beijing, China), at fivefold dilutions for 12 h. Human IFN-α2b is produced in Escherichia coli, human IFN-β is a recombinant glycoprotein produced in mammalian cells, and human IFN-ω is a recombinant glycoprotein produced in the yeast Pichia pastoris with a specific activity of 7×107 IU/mg. After washing, the medium was replaced with DMEM containing 2 μg/mL of TPCK-trypsin, and the monolayers were infected with Ca/04 or BJ/501 influenza virus at a MOI of 0.1. Infected cells without IFN pretreatments were used as a positive control. For the viral copy number assay, the plates were frozen at −20°C at 72 hpi, and RNA was extracted from cell lysates for viral real-time quantitative PCR.
RNA extraction and real-time quantitative PCR
Viral RNA was extracted from cell lysates with the QIAamp® Viral RNA Mini Kit (Qiagen, Hilden, Germany), according to the manufacturer's instructions. Extracted RNA was stored at −70°C until use. The primer used for reverse transcription was Oligo d(T)18, and cDNA was synthesized by incubation of the reaction mixture for 1 h at 37°C according to the M-MLV Reverse Transcriptase manual (Promega, Madison, WI).
PCR amplification was performed in glass capillary tubes using the LightCycler® system (Roche, Mannheim, Germany) with cDNA as the template. SYBR Green I (Takara Bio, Shiga, Japan) contained Taq polymerase and the ions necessary for the PCR reaction. Each reaction contained 1 μL of cDNA, 5 μL of SYBR Green I, 0.1 μL of each forward and reverse primer, and was made up to 10 μL with water. The first step of amplification was denaturation for 30 sec at 95°C, followed by 40 cycles of amplification at 55°C for 5 sec, and 72°C for 10 sec. Quantitative PCR targeting the 2009 A (H1N1) influenza virus M gene was performed with the primer sets 5′-ACTACC ACCAATCCACTA-3′/5′-GTCATCTTTCAGACC AGC-3′. During the amplification process, fluorescence emission was monitored by the Ct values. The Ct threshold value represents the cycle number from which a significant increase in fluorescence was detected. Ten-fold dilutions of the reconstructed plasmid pMD-M were used as standard samples. The Ct threshold values were lower for more concentrated samples of plasmid DNA template. A standard curve was constructed plotting the logarithm of the concentration of each dilution of plasmid on the x-axis, against the Ct values of the plasmid samples on the y-axis. From the standard dilution curve created, the concentrations of the unknown samples were calculated according to their Ct values. After calculation of sample concentration, the data were converted to the logarithm of the relative copy numbers per milliliter of cell lysate based on the sizes of the RT-PCR reaction system and the Q-PCR reaction system. The detection limit of this assay was 104 copies per milliliter.
Animals
Female Hartley strain guinea pigs weighing between 250 and 300 g were obtained from the laboratory animal center at the National Institute for the Control of Pharmaceutical and Biological Products (Beijing, China). During challenge studies, the guinea pigs were housed in an animal device in a BSL-3 laboratory. The animals were allowed free access to food and water. Prior to each manipulation, the guinea pigs were anesthetized by the intramuscular injection of ketamine (30 mg/kg of body weight). Animal experimentation was approved by the Institutional Animal Care and Use Committee of the Beijing Institute of Microbiology and Epidemiology.
Mx and OAS-1a protein expression analysis
The responsiveness of guinea pigs to human IFN-ω was assessed by analyzing the expression of Mx and 2,5′-oligoadenylate synthetase 1a (OAS-1a) using reverse transcriptase PCR (RT-PCR) or Western blot. The guinea pigs were treated with intranasal PBS or 5.0×105 IU/kg of IFN-ω on day 0 post-infection, and individual guinea pigs were euthanized on days 1, 2, and 3 post-infection. Lung samples were collected from the animals, and half of each sample was homogenized in 1 mL of PBS for RT-PCR or Western blot. For Mx expression, total RNA from lung tissues was extracted using Trizol® reagent (Invitrogen, San Diego, CA), and cDNA was synthesized as described above. The 30-cycle PCR was performed using Mx primer sets 5′-GCTCTGTGCTGGAAGCACTGTCT-3′/5′-GACCACCACCAGGTTGA-3′. β-Actin primer sets 5′-CAACCGCG AGAAGATGAC-3′/5′-AGGGTACATGGTGGTGCC-3′ were used as an internal control. For OAS-1a expression, the homogenate was supplemented with protease inhibitors (1 mM PMSF, 1 μg/mL leupeptin, 1 μg/mL pepstatin, 2 μg/mL aprotinin, 10 mM NaF, and 1 mM Na3VO4). Following centrifugation at 13,000 g for 10 min at 4°C, the supernatant was collected. The protein concentration of each sample was determined by bicinchoninic acid protein assay. Equal amounts of protein were separated by 12% SDS-PAGE, transferred to nitrocellulose, and incubated with antibodies against Oas-1a (E-12, sc-49835). Following incubation with the HRP-conjugated secondary antibody, proteins were detected using the Western blot luminol reagent. Anti-β-actin (sc-1616-R) served as a loading control. All antibodies and luminol reagent were purchased from Santa Cruz Biotechnology.
IFN treatment and viral challenge in guinea pigs
During challenge studies, guinea pigs were equally divided into three groups: an IFN-α group, an IFN-ω group, and a PBS group. For the IFN group, a dose of recombinant human IFN-α or IFN-ω corresponding to 5.0×105 IU/kg of the average weight of all animals was given intranasally in 300 μL of PBS, with 150 μL instilled into each nostril from day 1 prior to intranasal inoculation with influenza virus, to day 5 post-infection. For viral challenge, 105 TCID50 of Ca/04 influenza virus in 300 μL of PBS was inoculated intranasally into each animal on day 0. The control group was treated with PBS with the same manipulation, and the animal experiments were done twice.
To determine the ability of virus to replicate in the lower respiratory tract, lung tissues from 3 animals of each group were harvested every day from day 1 post-infection to day 6 post-infection. Prior to the collection of lung samples, the animals were deeply anesthetized as described above and exsanguinated. Lung tissues for titration were removed and immediately frozen at −70°C. For viral titer analysis, half of each lung sample was homogenized in 1 mL of PBS. Tissue homogenates were clarified by centrifugation, and viral titers were expressed as TCID50 in MDCK cells according to the Spearman-Karber formula.
To determine the ability of the virus to replicate in the upper respiratory tract, 1 mL of PBS was introduced into the cut trachea of each guinea pig, and the liquid from the nostrils was harvested for virus titration.
Statistical analysis
GraphPad Prism 5.0 software was used for statistical analyses, and two-way analysis of variance (ANOVA) was used for analyzing the significance. p Values<0.05 were considered statistically significant.
Results
Effects of human IFNs on 2009 A (H1N1) influenza virus in vitro
Cytopathic effect (CPE) was observed in Ca/04 and BJ/501 influenza virus-infected Caco-2 cells and MDCK cells. Specific antibody against H1N1 influenza virus HA could react with Ca/04 and BJ/501 influenza virus strains in both types of cells with similar immunostaining profiles (Fig. 1). The CPE in Caco-2 cells was much less apparent than in MDCK cells, while there were no significant differences in genomic copies between the two cell lines.
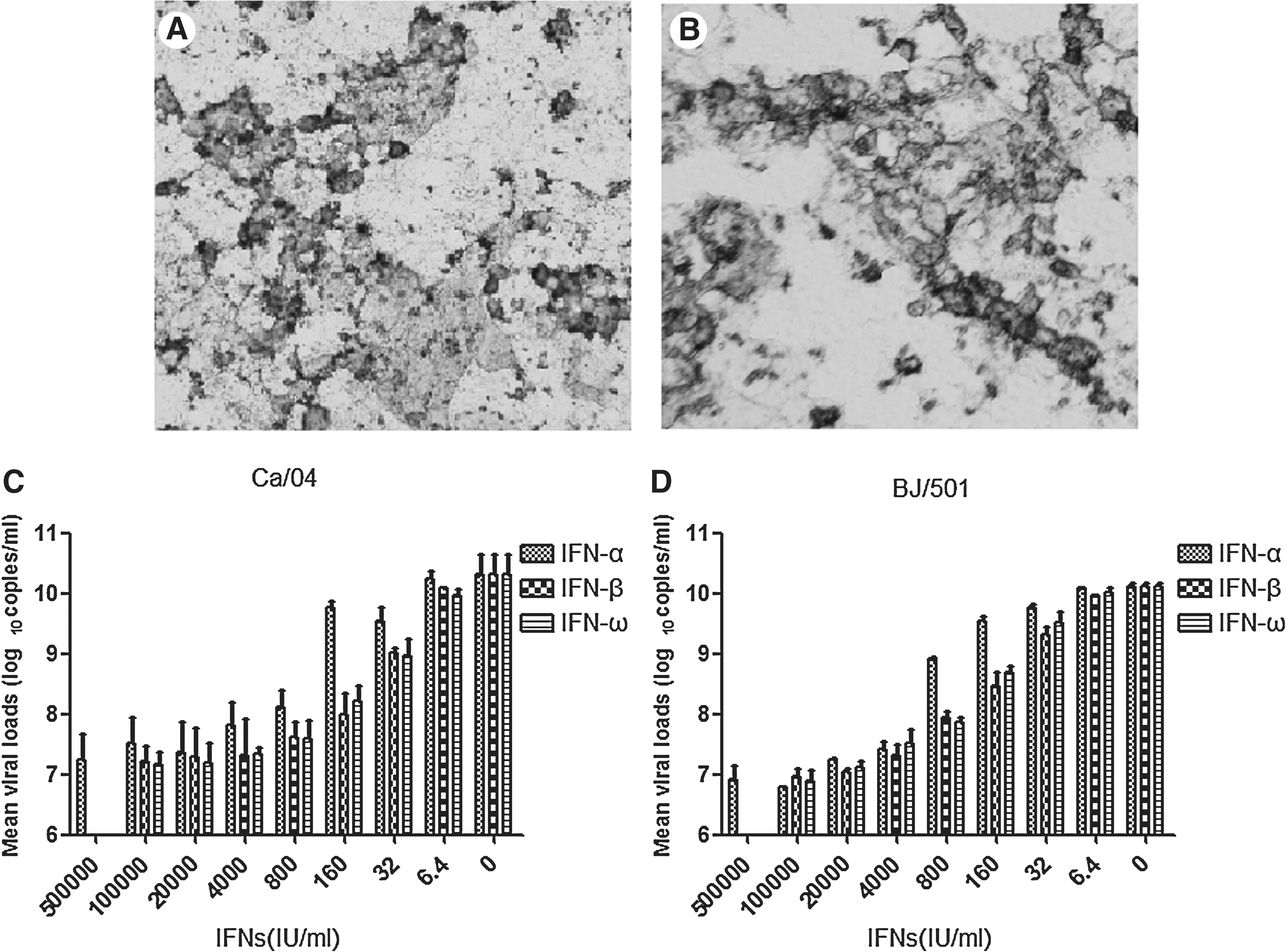
IFN-α, IFN-β, and IFN-ω showed a dose-dependent inhibition of the production of pandemic 2009 A (H1N1) influenza virus in Caco-2 cells. (
Pretreatment with IFN-α, IFN-β, and IFN-ω showed a dose-dependent inhibition of the production of virus in Caco-2 cells, as detected by real-time PCR (Fig. 1). However, in cells pretreated with high concentrations of IFNs and without visible CPE, IFN pretreatments did not completely reduce virus titer. This result indicated that IFNs were not able to block virus infection though they reduced virus replication. In cultures pretreated with different IFNs, the viral loads were reduced by 5000-fold at 1.0×105 IU/mL of IFNs compared to the untreated controls, where only virus was added.
When comparing the inhibitory effects of different IFNs on Ca/04 influenza virus, IFN-β and IFN-ω were significantly more potent than IFN-α at 160 IU/mL (p<0.05), with no significant difference at other concentrations. Similar results were observed for BJ/501 influenza virus, for which IFN-β and IFN-ω were more potent than IFN-α at 800 and 160 IU/mL (p<0.05). IFN-β and IFN-ω showed similar inhibitory activity against both strains at every concentration of IFNs (p<0.05).
Induction of Mx and OAS-1a with intranasal IFN-ω
It has been demonstrated that exogenous IFN-α treatment can induce an antiviral response in guinea pigs (5). To determine whether exogenous human IFN-ω was capable of inducing an antiviral response in this species, induction of Mx and OAS-1a was examined following a single treatment with recombinant human IFN-ω. Mx protein is an IFN-inducible nuclear protein and is critical for recovery from infection by influenza viruses (15,16). OAS-1a belongs to interferon-induced proteins that play a putative role in mediating resistance to virus infection, control of cell growth, differentiation, and apoptosis (17). As early as day 1 following intranasal treatment with 5.0×105 IU/kg of recombinant human IFN-ω, a robust induction of Mx mRNA and OAS-1a protein was observed in IFN-treated animals compared to PBS-treated animals (Fig. 2). A lower level of Mx mRNA and OAS-1a was observed on day 2 and day 3 post-infection, with no Mx mRNA and OAS-1a seen in PBS-treated animals during this period.

Expression of Mx and OAS-1a in lung tissues of guinea pigs after intranasal IFN-ω treatment. Mx and OAS-1a protein expression in the guinea pig lungs were examined by RT-PCR or Western blot following a single treatment with either PBS or 5.0×105 IU/kg of IFN-ω. β-actin was used as an internal control.
The protective effect of human IFN-ω against 2009 A (H1N1) influenza virus in guinea pigs
Having demonstrated that exogenous recombinant human IFN-ω treatment could induce an antiviral response in guinea pigs, we next examined the ability of IFN-ω treatment to inhibit the replication of Ca/04 influenza virus in guinea pigs, with IFN-α treatment as positive control and PBS treatment as negative control. The animals were treated intranasally with IFN-α, IFN-ω, or PBS prior to and following virus inoculation on day −1 to day 5 post-infection. On day 0, approximately 6 h following treatment, the animals were inoculated intranasally with 105 TCID50 of Ca/04 influenza virus. Three guinea pigs in each group were killed in order to evaluate viral loads in nasal washes and lung tissues from day 1 to day 6 post-infection. The viral titers in nasal washes were low (not exceeding 103.0 TCID50/mL) from day 1 to day 4 post-infection, and could not be detected on day 5 and day 6 post-infection. There was no significant difference in the reduction of viral titers observed in nasal washes between IFN-treated animals and PBS-treated animals (p>0.05, Fig. 3), perhaps because of the low viral titers of Ca/04 influenza virus in the guinea pigs' nasal washes. In the lungs of infected animals, the viral titers reached 105.5 TCID50/mL, and there was a significant reduction of viral titers, by 1000-fold, in the IFN-treated animals compared to the PBS-treated animals (p<0.05, Fig. 3). IFN-ω showed a similar inhibitory effect against Ca/04 influenza virus to IFN-α. In the PBS-treated group, the viral peak in lung tissues appeared on day 3 post-infection and declined afterwards. In the IFN-treated group, the viral titers in lung tissues (not exceeding 102.5 TCID50/mL) decreased from day 1 after virus exposure. This finding demonstrated that IFN treatment, though unable to block Ca/04 virus infection, could reduce virus replication in the lung tissues.
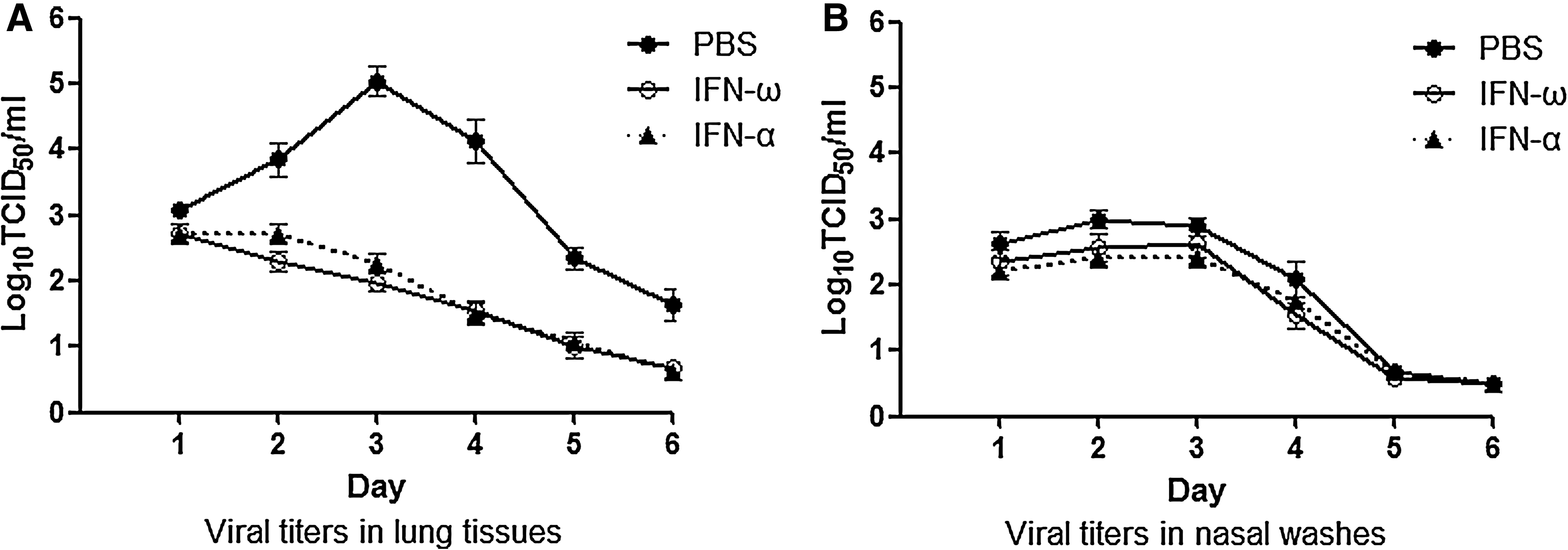
Replication of Ca/04 influenza virus in PBS-treated and IFN-treated guinea pigs. Following daily treatment with intranasal human IFN-ω or IFN-α from days −1 to 5 post-infection, and viral challenge on day 0, three guinea pigs in each group were killed at day 1 to day 6 post-infection. Viral titers from lung tissues and nasal washes of infected animals were assayed by TCID50 in MDCK cells by use of the Spearman-Karber formula. The detection limit of this assay was 0.5 log10 TCID50/mL. The experiments were done twice.
Discussion
IFNs are key players in early host responses to viral infections. Previous studies have shown that intranasal or oral administration of IFN-α protects mice, ferrets, and guinea pigs from influenza virus challenge (3 –5). In the late 1960s and early 1970s leukocyte IFN became available in nasal spray preparations in Moscow pharmacies for use as both prophylaxis and treatment of influenza. However, the reported effects on influenza were minimal, and the side effects from intranasal administration were considerable (18,19). In the current study, recombinant human IFN-ω was evaluated for its protective efficacy against 2009 A (H1N1) influenza viruses in vitro and in guinea pigs.
The recombinant human IFN-ω used in this work is presently in phase I clinical studies for HCV treatment, and its antiviral activity against vesicular stomatitis virus, severe acute respiratory syndrome-coronavirus, and enterovirus 71 was evaluated in our laboratory. To evaluate the inhibitory ability of IFN-ω towards 2009 A (H1N1) influenza virus, several human cell lines were first screened for their sensitivity to this virus, including Caco-2, Hep-2, A549, and MRC-5 cells, because MDCK cells did not respond to the addition of human IFN-ω. 2009 A (H1N1) influenza virus caused an obvious CPE only in Caco-2 cells. The viral genomic copies were detected at the same level in both Caco-2 and MDCK cell lines, which indicated that the virus had similar growth profiles in both cell lines. Li et al. tested the differential susceptibility of different cell lines to swine-origin influenza A [pandemic 2009 A (H1N1) influenza virus], in which Caco-2 and MDCK cells showed good susceptibility to swine-origin influenza A (H1N1) virus (20).
In this study, in cultures that were pretreated with different IFNs, the viral loads reached a 5000-fold reduction compared to the untreated control. The antiviral activities of IFN-β and IFN-ω are superior to IFN-α at some concentrations, and the difference in potency may be a result of differential glycosylation. In contrast to IFN-α (produced in E. coli), the IFN-β (produced in CHO cells) and IFN-ω (produced in yeast) in this work are glycosylated. A previous study indicated that glycosylation has an important effect on the activity of IFNs (9).
Mice, ferrets, and guinea pigs have been used as animal models for 2009 A (H1N1) influenza virus infection and developed respiratory disease and lung pathology (3 –5). In this study the choice of guinea pigs as an animal model for 2009 A (H1N1) influenza virus was based on several considerations. First, although the ferret is considered to be an ideal animal model for influenza virus infection, ferrets are not available as an animal model in our institute. Second, we challenged BALB/c mice with 2009 A (H1N1) influenza virus in pre-experiments and infection was not detected in every challenged mouse. However, infection was detected in every challenged guinea pig. Furthermore, guinea pigs are responsive to human IFN-ω treatment. Maines et al. found that infection of ferrets with 2009 A (H1N1) influenza viruses caused increased morbidity and replicated to higher titers in lung tissue compared to seasonal influenza virus (11). Infection of guinea pigs with Ca/04 influenza virus in this study did not result in any weight loss or other pronounced clinical signs at any time point following infection. However, virus could be detected both in nasal washes and in lung tissues of guinea pigs. Higher titers of virus were found in lung tissue than in nasal washes, while minimal pneumonia was observed in lung pathology (data not shown). These observations suggested that 2009 A (H1N1) influenza virus was able to replicate efficiently in the upper and lower respiratory tracts of guinea pigs, though it was not highly pathogenic in this animal model.
Conclusion
In summary, repeated intranasal application of recombinant human IFN-ω significantly reduced the replication of 2009 A (H1N1) influenza virus in lung tissues, and this reduction in titers may be useful in alleviating lung damage. However, this beneficial effect should be weighed against a possible negative impact of IFN-ω treatment on the course of disease.
Footnotes
Acknowledgment
This work was supported by the National Natural Science Foundation of China (81025018).
Author Disclosure Statement
No conflicting financial interests exist.