
Abstract
Existing therapies for chronic viral infections are still suboptimal or have considerable side effects, so new therapeutic strategies need to be developed. One option is to boost the host's immune response with cytokines. We have recently shown in an acute ex vivo HIV infection model that co-administration of interferon (IFN)-α and interleukin (IL)-7 allows us to combine the potent anti-HIV activity of IFN-α with the beneficial effects of IL-7 on T-cell survival and function. Here we evaluated the effect of combining IFN-α and IL-7 on viral replication in vivo in the chronic lymphocytic choriomeningitis virus (LCMV) and acute Friend retrovirus (FV) infection models. In the chronic LCMV model, cytokine treatment was started during the early replication phase (i.e., on day 7 post-infection [pi]). Under the experimental conditions used, exogenous IFN-α inhibited FV replication, but had no effect on viral replication in the LCMV model. There was no therapeutic benefit of IL-7 either alone or in combination with IFN-α in either of the two infection models. In the LCMV model, dose-dependent effects of the cytokine combination on T-cell phenotype/function were observed. It is possible that these effects would translate into antiviral activity in re-challenged mice. It is also possible that another type of IFN-α/β or induction of endogenous IFN-α/β alone or in combination with IL-7 would have antiviral activity in the LCMV model. Furthermore, we cannot exclude that some effect on viral titers would have been seen at later time points not investigated here (i.e., beyond day 34 pi). Finally, IFN-α/IL-7 may inhibit the replication of other viruses. Thus it might be worth testing these cytokines in other in vivo models of chronic viral infections.
Introduction
IFN-α plays a key role in the host's natural defense against viral infections: it is primarily part of the innate immune response, but it has also a definite role in the generation of the adaptive immune response (2,3). Despite this broad activity, IFN-α alone has only limited success in treating chronic viral diseases.
Complementing IFN-α's activity with a cytokine acting primarily on T cells may be beneficial in the generation of an efficient immune response against chronic viral infections. An ideal such partner might be interleukin (IL)-7, which is important for sustained proliferation and survival of thymocytes, and which regulates peripheral T cell homeostasis (4,5). Furthermore, IL-7 is critical for generating T-cell memory (6). In phase I and I/IIa clinical trials, recombinant human IL-7 substantially increased the number of circulating T cells in cancer and HIV-infected patients (7 –10). Notably, the increase pertained to CD4+ and CD8+ T cells of both the naïve and memory cell subsets (11). Previously, IL-2 has been studied in humans for enhancing cellular immune responses. However, two phase III trials, ESPRIT and SILCAAT, failed to show any benefit of IL-2 immunotherapy in HIV-infected individuals treated with antiretroviral therapy (12). Recently, it has been found that IL-2 specifically expands regulatory T cells (Tregs), a T-cell subset with immunosuppressive activity, that might explain the lack of clinical benefit seen in the ESPRIT and SILCAAT trials (13). Such an expansion of Tregs does not occur with IL-7. Importantly, IL-7 was well tolerated by the patients in the phase I and I/IIa trials, which is a major advantage for potential future applications (11).
We have previously shown that treatment of human tonsillar lymphocyte aggregate cultures infected ex vivo with HIV with IFN-α and IL-7 permitted combining the anti-HIV activity of IFN-α with the beneficial effects of IL-7 on T-cell survival and function (14). These findings encouraged us to evaluate the immunomodulatory and antiviral effects of this cytokine combination in vivo. While humanized mice are very promising for the study of HIV infection, we preferred to evaluate this cytokine combination in mouse models with a fully functional immune system. We chose the lymphocytic choriomeningitis virus (LCMV) and Friend retrovirus (FV) infection models. LCMV is a natural pathogen of mice (15). It replicates to high titers in several organs. The main antiviral immune mechanism constraining LCMV is based on the induction of virus-specific CD8+ T cells (16,17). LCMV can cause either acute or persistent infection, depending on the viral strain, route of infection, and inoculation dose.
FV infection is the result of a co-infection with the Friend murine leukemia virus (F-MuLV) and the spleen focus-forming virus (SFFV). F-MuLV is a replication-competent apathogenic helper virus, whereas SFFV is a defective pathogenic virus (18). Depending on the mouse strain, FV infection results in an acute infection with either development of erythroleukemia and death, or subsequent recovery and persistent infection (19). Recovery from acute FV infection requires virus-specific CD8+ T cells and virus-neutralizing antibodies, whereas effector CD4+ T cells are the critical T-cell subset constraining persistent FV (20 –22).
Thus, we studied the in vivo effect of IFN-α/IL-7 on viral replication in the chronic LCMV model, with cytokine treatment starting during the early replication period (i.e., on day 7 post-infection [pi]), and in the acute FV model.
Materials and Methods
Mice and cytokines
Mice were obtained from the Jackson Laboratory. C57BL/6 mice were ordered via Harlan Laboratories, the Netherlands. A.BY and B10.A were ordered via Charles River, Germany, and crossed in the BZL, University Hospital Zurich, to obtain (B10.A×A.BY) F1 mice (H-2a/b). All experiments with mice were approved and conducted according to local guidelines and laws.
Peginterferon alfa-2a (PEG-IFN-α2a; Pegasys®) and Peginterferon alfa-2b (PEG-IFN-α2b; PegIntron®) were kindly provided by Hoffman LaRoche and Schering-Plough, respectively. Human interferon alpha A/D (IFN-αA/D; Universal Type I IFN) was purchased from Pestka Biomedical Laboratories. Recombinant glycosylated human IL-7 (CYT107) was provided by Cytheris.
Real-time quantitative PCR
RNA was isolated from blood and spleens using the QIAamp RNA Blood Mini Kit (Qiagen, Valencia, CA), per the manufacturer's recommendations. Reverse transcription and real-time quantitative (q)PCR were performed essentially as previously described (23), with the exception that commercially available primers and probes for the IFN-stimulated genes (ISGs) interferon regulatory factor 7 (IRF7) and myxovirus resistance 1 (Mx1) were used (Assays-on-Demand; Applied Biosystems, Carlsbad, CA). The mean normalized gene expression (MNE) for every sample was determined using the software application Q-Gene (calculation procedure for MNE 2) (24); phosphoglycerate kinase (Pgk) was used as the housekeeping gene.
Infection experiments
For the LCMV model, female C57BL/6 mice approximately 8 wk of age were infected intravenously (IV) with 2×106 plaque-forming units (PFU) of LCMV Cl-13 (kindly provided by Andreas Bergthaler). Starting on day 7 pi, mice were treated subcutaneously (SC) for 4 wk either once weekly with PEG-IFN-α2b (20 μg per injection), twice weekly with IL-7 (10 μg per injection), both cytokines, or were mock-treated with PBS (n=6 for the PBS group and n=5 for all other groups, Supplementary Fig. 1A; see online supplementary material at
In another experimental setup of the LCMV model, female C57BL/6 mice approximately 8 wk of age were infected as described above. Starting on day 7 pi, mice were treated SC for 4 wk either with 1×20 μg of PEG-IFN-α2b plus 2×10 μg of IL-7 per week (group I), 2×20 μg of PEG-IFN-α2b plus 3×10 μg of IL-7 per week (group II), 2×20 μg of PEG-IFN-α2b plus 2×100 μg of IL-7 per week (group III), or were mock-treated with PBS (n=4 for the PBS group and n=5 for all other groups, Supplementary Fig. 1B; see online supplementary material at
For the FV experiments, we used (B10.A×A.BY) F1 mice (H-2a/b), which have an intermediate susceptibility to FV (i.e., they develop splenomegaly and subsequent erythroleukemia more slowly than fully-susceptible mice). Female mice 8–28 wk of age (mice of different ages were evenly distributed over the groups) were infected IV with 3×103 SFFU of FV (free of lactate dehydrogenase-elevating virus [LDV]; see below). Starting on day 4 pi, mice were treated daily SC until day 8 with IFN-αA/D (4×104 U per injection), or three times every other day with IL-7 (10 μg per injection), or with a combination of both cytokines; the mock-treated controls received PBS (n=6 for all groups, Supplementary Fig. 1C; see online supplementary material at
In another experimental setup of the FV model, female (B10.A×A.BY) F1 mice approximately 24 wk of age (mice of different ages were evenly distributed over the groups) were infected as described above, and treated with either IFN-αA/D (2×104 U per injection) on days 4, 5, and 6 pi, IL-7 (10 μg per injection) on days 7, 9, and 11 pi, or with a combination of both cytokines; the mock-treated controls received PBS (n=6 for all groups, Supplementary Fig. 1D; see online supplementary material at
Immunostaining and flow cytometry
To assess the proliferative response of lymphocytes from uninfected female C57BL/6 mice to treatment with different doses of IL-7, blood was collected from the heart and lysed with ACK cell lysing buffer, and cryopreserved splenocytes were used. The cells were incubated with anti-CD4, anti-CD8, and anti-B220 antibodies (clones H129.19, 53-6.7, and RA3-6B2, respectively), fixed/permeabilized with Cytofix/Cytoperm (BD Biosciences Pharmingen, Franklin Lakes, NJ), and subsequently stained with an antibody against Ki-67.
The frequency of LCMV-specific CD8+ T cells was determined by first re-stimulating splenocytes with 10 μg/mL of LCMV glycoprotein (gp)33 (kindly provided by Andreas Bergthaler) for 4 h in the presence of GolgiPlug (BD Biosciences Pharmingen); control cells were not re-stimulated. The cells were then incubated with anti-CD8 antibody, fixed/permeabilized, and subsequently stained for intracellular IFN-γ. To quantify the frequencies of CD4+ and CD8+ T cells, splenocytes were stained with anti-CD4 and anti-CD8 antibodies. The frequencies of proliferating T cells were assessed by incubating splenocytes with anti-CD4 and anti-CD8 antibodies, fixation/permeabilization, and staining for intracellular Ki-67. Cell surface expression of CD127 (= IL-7 receptor α chain) was determined by co-staining of splenocytes for CD4, CD8, and CD127. In the case of IFN-γ production and expression of Ki-67 and CD127, the mean fluorescence intensity (MFI) was also evaluated.
All antibodies were from BD Biosciences Pharmingen. Acquisition was made on cells defined as lymphocytes based on their FSC and SSC properties on a FACSCalibur (BD Biosciences Pharmingen), or a CyAn ADP (Beckman Coulter, Brea, CA). Data were analyzed with FlowJo software (Tree Star, Ashland, OR).
Statistical analysis
Differences between cytokine and control treatments were tested for significance by one-way analysis of variance (ANOVA) with Dunnett's multiple comparison test, using GraphPad Prism 5.0 software. p Values <0.05 were considered statistically significant.
Results
Screening of different types of IFN-α and IL-7 dose-finding experiments
Human IFN-α exists in 13 subtypes with distinctive biological properties, whereas only one recombinant IFN-α protein (i.e., the allelic variants IFN-α2a and IFN-α2b), is used therapeutically (27,28). We evaluated three different interferons: the pegylated versions of the two IFN-α2 variants (PEG-IFN-α2a; Pegasys, Hoffman LaRoche; and PEG-IFN-α2b, PegIntron, Schering-Plough), which have improved pharmacokinetics and pharmacodynamics compared to the unmodified IFN-α2 variants (29), as well as the hybrid IFN-αA/D, which exhibits high activity on mouse cells, a property not observed with the parental molecules (30).
We reasoned that a pegylated version of IFN-α would reduce the number of doses needed, and thus might be better suited for administration over an extended period such as in our chronic LCMV model. Thus we tested PEG-IFN-α2a and PEG-IFN-α2b in comparison to universal IFN-αA/D for their capacity to upregulate the IFN-stimulated genes IRF7 and Mx1 in uninfected C57BL/6 mice. Both PEG-IFN-α2b and IFN-αA/D significantly upregulated IRF7 and Mx1 mRNA expression in blood and spleen 6 h post-treatment, whereas PEG-IFN-α2a did not have any effect (Fig. 1). We therefore decided to use PEG-IFN-α2b in the chronic LCMV mouse model, and the universal IFN-αA/D in the acute FV mouse model.
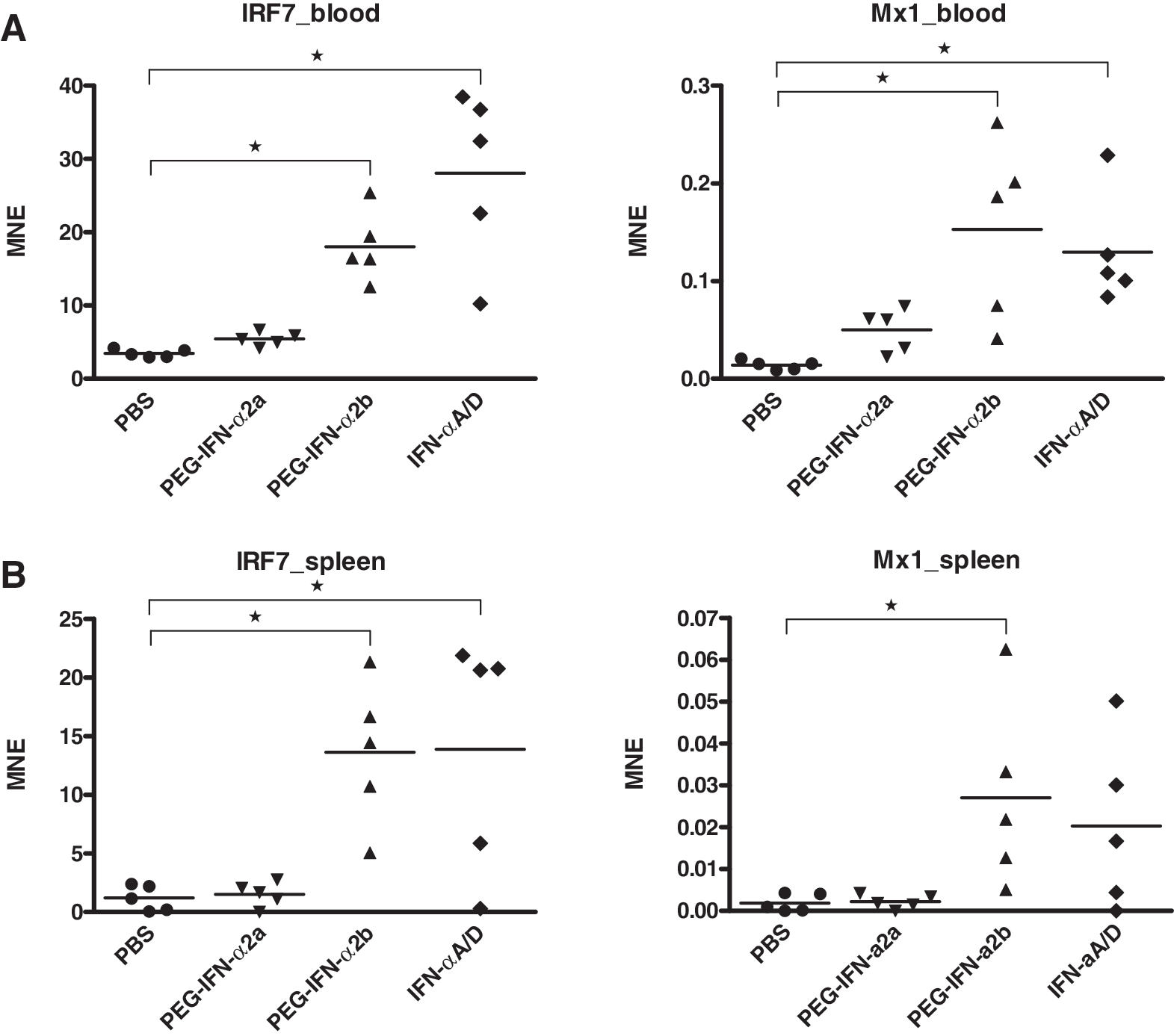
Screening of different types of IFN-α. Female C57BL/6 mice approximately 8 wk of age were treated SC either with 20 μg of PEG-IFN-α2a or PEG-IFN-α2b, or with 4×104 U of IFN-αA/D. Expression of IRF7 (left panels) and Mx1 (right panels) was measured 6 h later in blood (
For IL-7, we also performed dose-finding experiments. C57BL/6 mice were injected with 1, 10, or 100 μg of IL-7, and 72 h later proliferation of T cells in blood and spleen was determined by immunostaining/flow cytometry using Ki-67 as a proliferation marker. While significant increases of the percentages of Ki-67+ cells were only obtained with 100 μg of IL-7, there was also a clear increase of the frequency of proliferating B220+ cells and CD8+ T cells with 10 μg of IL-7; for CD4+ T cells, there was a 1.6- and 1.4-fold increase on average in blood and spleen, respectively (Fig. 2). Thus, we decided to use IL-7 initially at 10 μg per injection with two administrations per week in the chronic LCMV model, and repeated administration in the acute FV model.
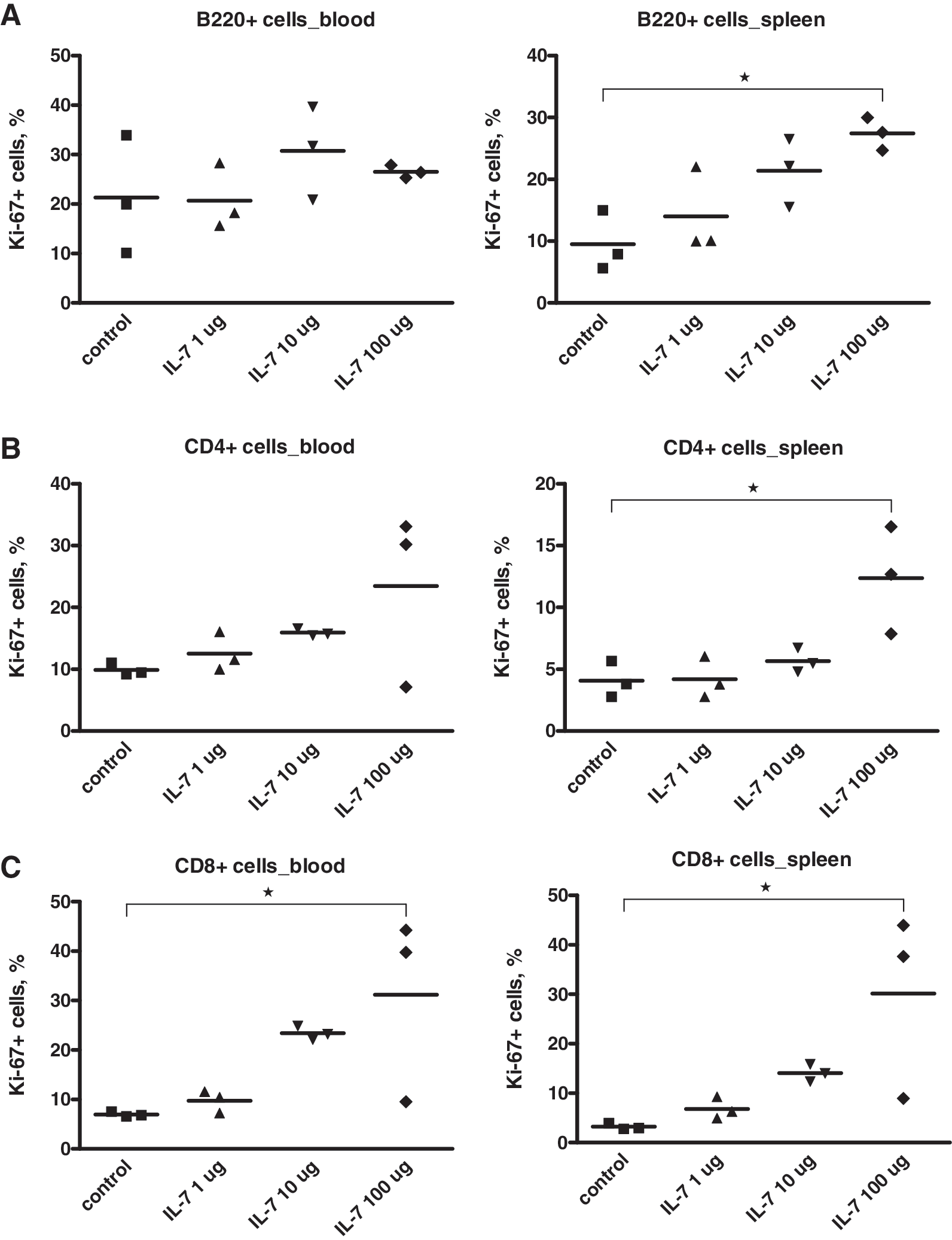
IL-7 dose-finding experiments. Female C57BL/6 mice approximately 8 wk of age were treated IP with 1, 10, or 100 μg of IL-7 per injection; control mice were mock-treated with PBS. Proliferation of B cells (
No effect of IFN-α and/or IL-7 on viral load in the LCMV model despite biological effects of the cytokine combination on T cells
We studied the effect of IFN-α and/or IL-7 on viral replication in vivo in the chronic LCMV model (LCMV strain clone 13), in which the function of CD8+ T cells gets impaired within 1 week pi, depending on the virus dose (31). We started administration of cytokines on day 7 pi, with the aim to prevent the ongoing functional exhaustion of LCMV-specific CD8+ T cells. Mice were treated for 4 wk either once weekly with PEG-IFN-α2b, twice weekly with IL-7, with both cytokines, or mock-treated with PBS (Supplementary Fig. 1A; see online supplementary material at
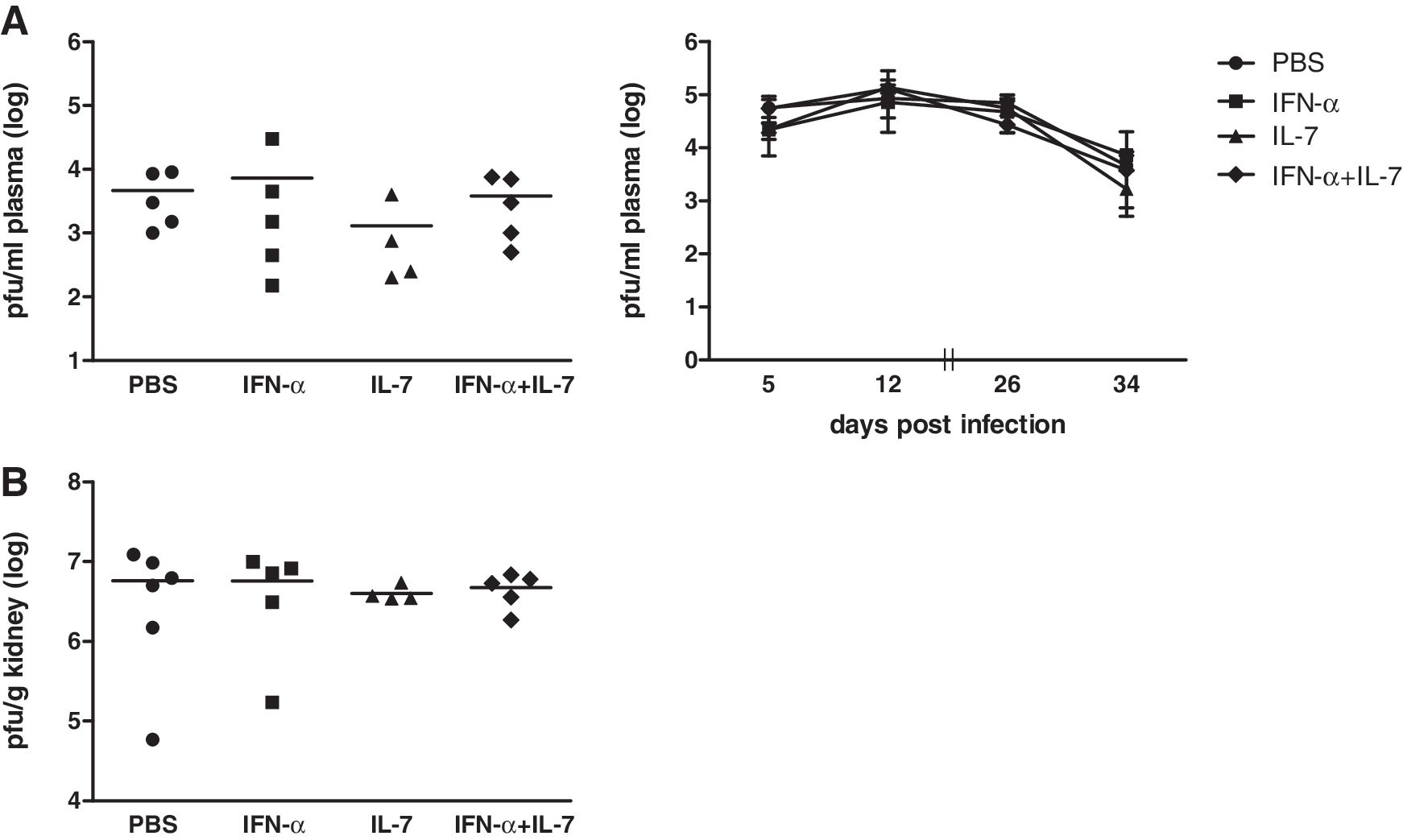
IFN-α and/or IL-7 have no effect on viral load in the LCMV model. Female C57BL/6 mice approximately 8 wk of age were infected IV with 2×106 PFU of LCMV strain Cl-13. Mice were treated starting on day 7 pi either once weekly with 20 μg of PEG-IFN-α2b, twice weekly with 10 μg of IL-7, both cytokines, or mock-treated with PBS. Viral titers were determined with an immunofocus assay of plasma on days 5, 12, 26, and 34 pi (
Since in the previous experiment with 20 μg of PEG-IFN-α2b once weekly, and/or 10 μg of IL-7 twice weekly, we had not seen any effect on viral load, we increased the dose and/or number of administrations of both cytokines in the combination. Mice were treated with either 1×20 μg of PEG-IFN-α2b plus 2×10 μg of IL-7 per week (group I, same dosing regimen as that of the previous experiment), 2×20 μg of PEG-IFN-α2b plus 3×10 μg of IL-7 per week (group II), or 2×20 μg of PEG-IFN-α2b plus 2×100 μg of IL-7 per week (group III) (Supplementary Fig. 1B; see online supplementary material at
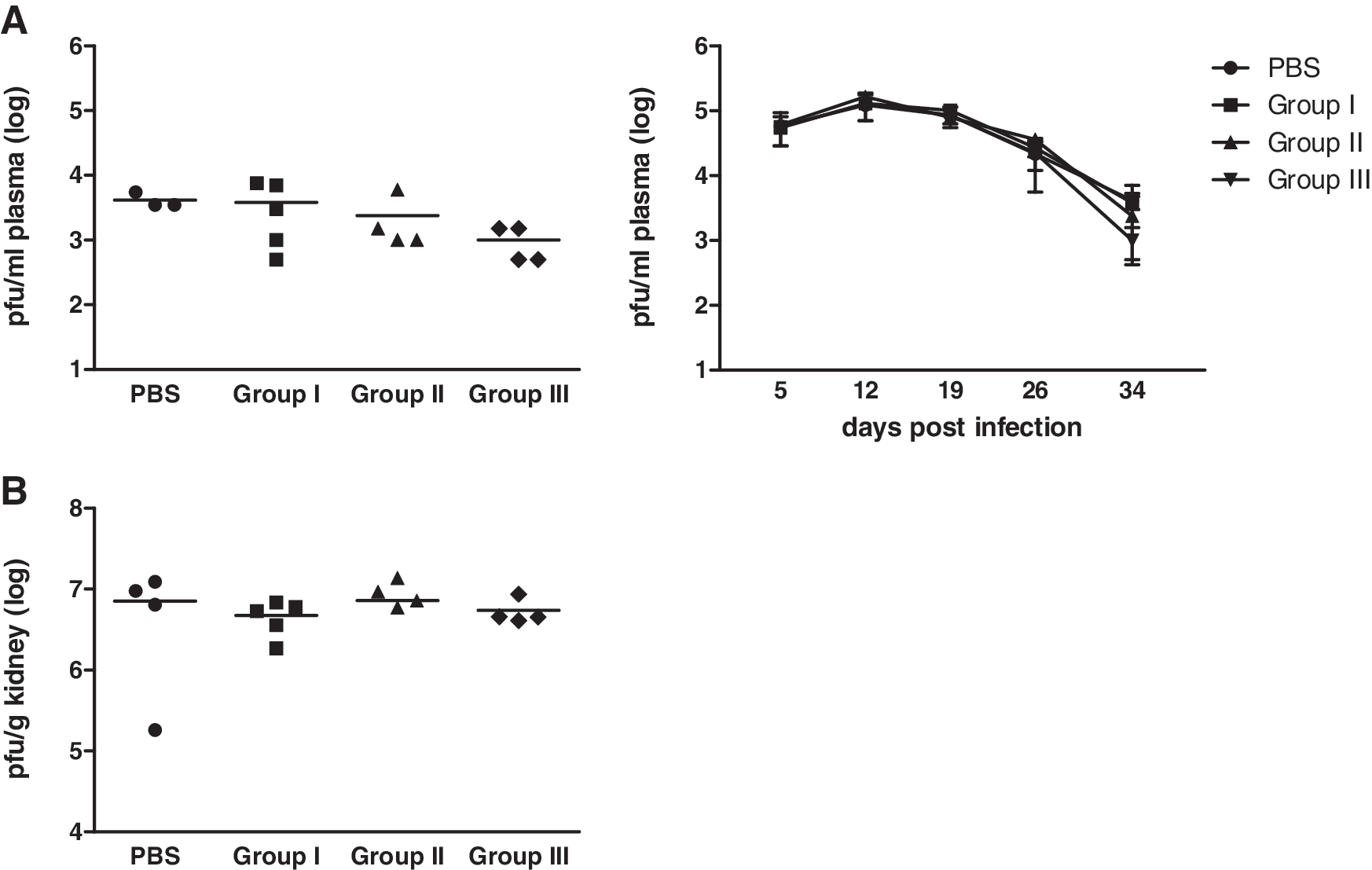
Increasing the dose and/or number of administrations of IFN-α and/or IL-7 does not result in viral control. Mice were infected as described in the legend to Fig. 3 and treated starting on day 7 pi, either with 1×20 μg of PEG-IFN-α2b plus 2×10 μg of IL-7 per week (group I), 2×20 μg of PEG-IFN-α2b plus 3×10 μg of IL-7 per week (group II), 2×20 μg of PEG-IFN-α2b plus 2×100 μg of IL-7 per week (group III), or mock-treated with PBS (PBS). Viral titers were determined with an immunofocus assay from plasma on days 5, 12, 19, 26, and 34 pi (
To assess whether the cytokine combination would have biological effects despite its lack of effect on viral replication, we analyzed the following parameters of splenic T cells by immunostaining/flow cytometry: (1) IFN-γ production of CD8+ T cells after re-stimulation of splenocytes with LCMV gp33, (2) frequency of CD4+ and CD8+ T cells, (3) frequency of Ki-67+ CD4+ and CD8+ T cells, and (4) frequency of CD127+ CD4+ and CD8+ T cells. For the markers IFN-γ, Ki-67, and CD127, the MFI was also assessed. In group III, all parameters were significantly increased compared to control animals (Fig. 5). The frequency of Ki-67+ T cells was also significantly increased in group I, whereas the percentage of CD127+ T cells was also significantly increased in group I and group II. The percentages of total T cells and of LCMV-specific CD8+ T cells were only significantly higher in group III. The MFI data correlated with these findings (data not shown).
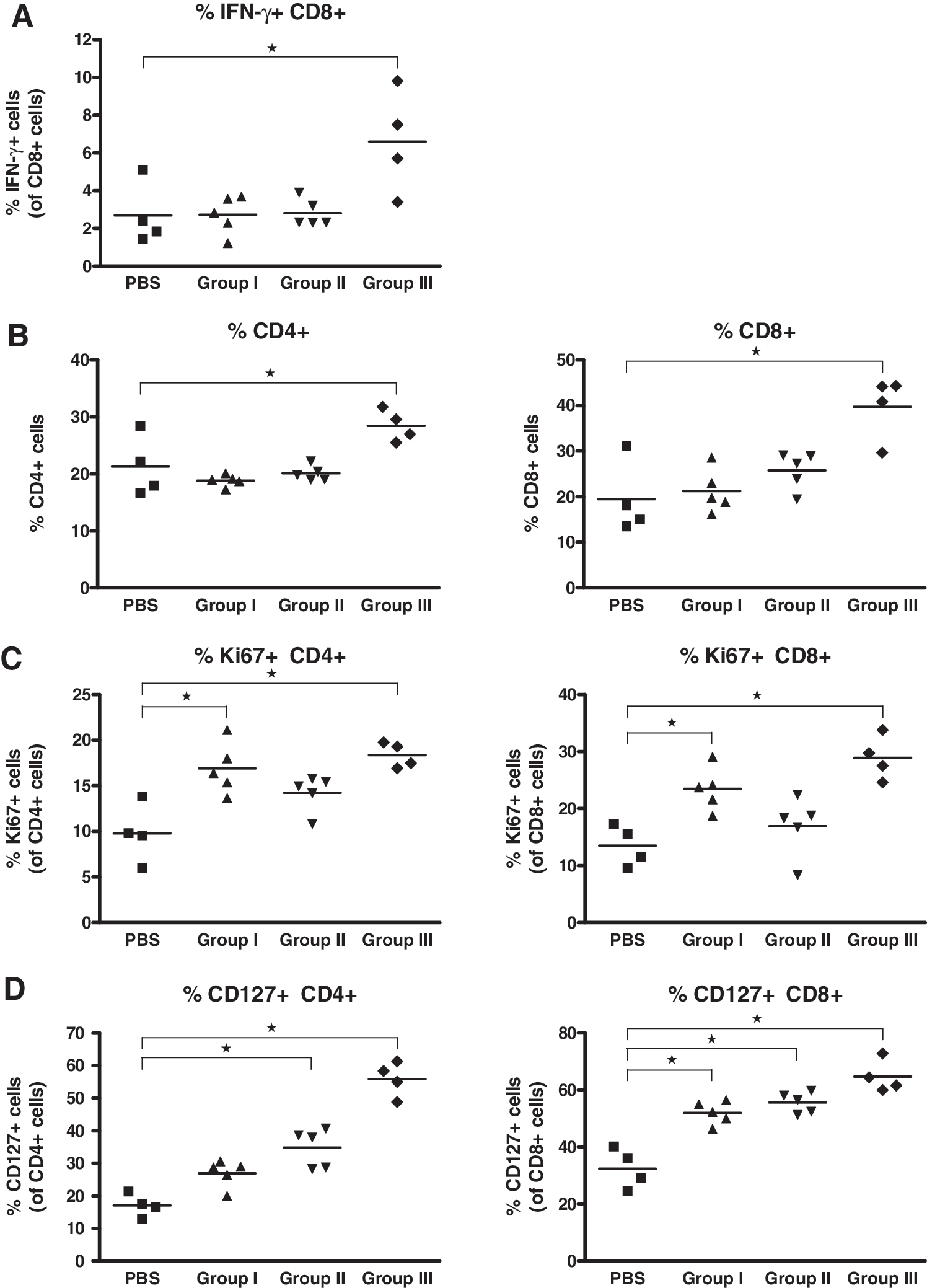
The combination of IFN-α and IL-7 modulates T-cell phenotype and/or function. Splenocytes were isolated from the spleens of the mice, the viral titers of which are shown in Fig. 4, and analyzed by immunostaining/flow cytometry for the frequency of CD8+ T cells producing IFN-γ after re-stimulation with LCMV gp33 (
Thus, the combination of IFN-α and IL-7 in the LCMV model dose-dependently increased antigen-specific and overall frequency, proliferation, and IL-7Rα expression of T cells, but these effects did not translate into suppression or decrease in viremia.
IFN-α alone, but not IL-7 or IFN-α combined with IL-7 inhibits FV
Given that IFN-α and/or IL-7 had no effect on viral replication in our chronic LCMV model, we decided to test the cytokines for their antiviral effect in another infection model. We chose the FV model, given that others and our group have previously shown antiviral effects of IFN-α and IL-7 in this model (32
–35). In the acute FV model (Supplementary Fig. 1C; see online supplementary material at
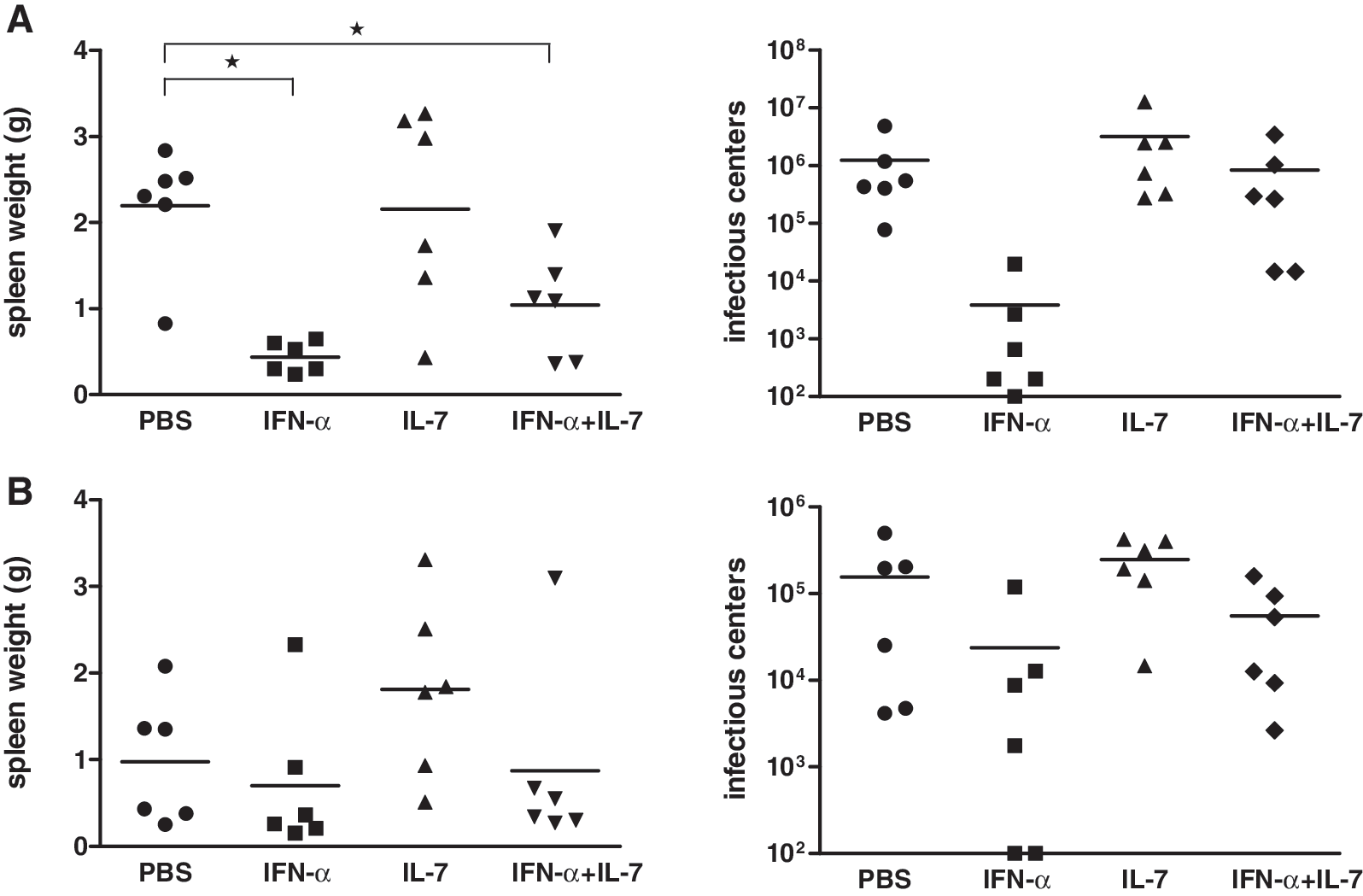
IFN-α alone, but not IL-7 or IFN-α combined with IL-7, inhibits FV. Female (B10.A×A.BY) F1 mice were infected IV with 3×103 SFFU of FV. (
Discussion
We have recently shown that treatment of human tonsillar lymphocyte aggregate cultures with IFN-α and IL-7 combines the potent anti-HIV activity of IFN-α with the beneficial effects of IL-7 on T-cell survival and function in an acute ex vivo HIV infection model (14). Here, we evaluated the effect of this cytokine combination on viral replication in vivo in two different mouse infection models (i.e., the LCMV and FV models). The main findings are: (1) dose-dependent effects of the cytokine combination on T-cell phenotype and/or function, but no effect of IFN-α and/or IL-7 alone on viral replication in the LCMV model, and (2) inhibition of FV replication with IFN-α, but no effect with IL-7, and strikingly a partial reversion of the anti-FV effect of IFN-α when IFN-α and IL-7 were combined.
To define the appropriate type of IFN-α for the treatment of LVMC- and FV-infected mice, we first compared three different types of IFN-α (i.e., PEG-IFN-α2a, PEG-IFN-α2b, and the hybrid IFN-αA/D) for their capacity to upregulate IFN-stimulated genes (ISGs) in uninfected C57BL/6 mice. PEG-IFN-α2b and IFN-αA/D, but not PEG-IFN-α2a, upregulated IRF7 and Mx1 expression. The distinct effects of PEG-IFN-α2a versus PEG-IFN-α2b are surprising, since they differ only in one amino acid at position 137 (37). As for IL-7, dose-finding studies revealed that 10 μg of IL-7 resulted in an increase of proliferating cells, in particular of CD8+ T cells; this increase became significant with 100 μg of IL-7. Thus, PEG-IFN-α2b, IFN-αA/D, and IL-7 clearly had biological effects in C57BL/6 mice.
We first tested IFN-α and IL-7 alone and in combination for their antiviral effect in mice infected with the LCMV clone 13 (38), whereby we started the cytokine treatments 7 days pi (i.e., early during replication). Strikingly, we did not observe any effect of IFN-α, IL-7, or their combination on LCMV Cl-13 titers, neither in blood at any of the time points investigated nor in kidneys at 34 days pi. Increasing the dose and/or number of administrations of IFN-α and/or IL-7 to up to 2×20 μg of PEG-IFN-α2b plus 2×100 μg of IL-7 per week also did not result in viral control. We did not examine viral titers beyond day 34 pi; thus we cannot exclude that some effect on viral titers would have been seen at later time points.
The lack of any effect of IFN-α on LCMV titers was surprising since previous reports showed at least some antiviral activity of IFN type I against LCMV. Treatment of C57BL/6 mice with polyclonal IFN-α/β antiserum caused increased titers of LCMV Cl-13 over an observation period of 5 d (39). The importance of IFN-α/β for clearance of persistent LCMV infection was also shown by the same group in a study using LCMV-WE carrier mice that lacked the receptor for IFN-α/β in adoptive transfer experiments (40). Differences in the experimental design between these previous studies and our study, such as strain and/or dose of LCMV, and obviously serotherapy (39) or the use of IFN-α/β receptor-deficient mice (40), as opposed to administration of IFN-α may explain the discrepant outcomes. The dose of 100 PFU of LCMV Cl-13 used in the study by Moskophidis et al. (39) resulted in acute infection, but may not have caused exhaustion of CD8+ T cells. We cannot exclude that another type of IFN-α would have been more efficacious in inhibiting LCMV replication, and help promote IL-7's stimulating immunological effects. We have recently shown that murine IFN-α subtypes differ greatly in their antiviral efficacy in vivo, at least in the acute FV model: the IFN-α subtypes α1, α4, or α9, but not α6, significantly reduced viral loads (32).
Two very recent studies also investigated the immunotherapeutic and antiviral effects of IL-7 on chronic LCMV Cl-13 infection (41,42). In all studies, including ours, IL-7 treatment was started at around 8 days pi (i.e., at the beginning of the contraction phase of the LCMV-specific CD8+ T cells). However, only the study by Pellegrini et al., in which the mice were treated once daily with 10 μg of rhIL-7 for 3 wk, showed significant effects on viral load. They found accelerated virus clearance from the spleen and liver by days 29 and 36 pi, respectively, and virus elimination from chronic reservoirs such as the kidney and brain by day 60 pi (42). The effect of IL-7 on viremia was not shown in this study. In the other study, in which rhIL-7 was administered daily at 5 μg/mouse between either days 8 and 15 pi, days 15 and 25 pi, or days 8 and 30 pi, there was only a trend of accelerated viral control with the latter experimental setup (41). Notably, we treated mice with a maximum dose of 2×100 μg per week. Together, these data point to dose- and/or dosing-dependent clearance of LCMV. However, the necessity to administer IL-7 daily is surprising considering the biological properties of the glycosylated rhIL-7; it has a longer half-life than the previous unglycosylated form of IL-7 [t½ of 6.5–9.8 h (43)], and is thus given only once weekly to patients in various ongoing clinical trials (
In contrast to the lack of an antiviral effect, the combination of IFN-α and IL-7 had effects on T-cell function/phenotype in our LCMV-infected mice. IFN-α/IL-7 dose-dependently increased (1) the frequency of antigen-specific CD8+ T cells as assessed in a recall antigen assay, (2) the frequency of CD4+ and CD8+ T cells, (3) CD127 expression on T cells, and (4) T-cell proliferation. Similarly, Nanjappa et al. did not observe significant reductions in viral titers with IL-7, despite the presence of immunotherapeutic effects (41). One may speculate that the immunological effects seen in our study were insufficient in their magnitude and/or were generated too late to constrain LCMV replication. We cannot exclude that this cytokine treatment applied early after acute LCMV infection might have an effect when re-challenging the mice, or that antigen-specific cells transferred to LCMV-naïve mice might attenuate LCMV infection.
Given that IFN-α and/or IL-7 had no effect on viral replication in our LCMV model, we decided to test the cytokine combination in the FV model. We have previously shown that exogenous IFN-αA/D and some murine IFN-α subtypes, as well as endogenous IFN-α induced by polyinosinic-polycytidylic acid, reduce acute FV replication (32 –34). Furthermore, Lu et al. have shown that IL-7 significantly prolongs the survival time of FV-infected mice and decreases viral replication (35). Indeed, in the present study IFN-α inhibited FV replication, whereas IL-7 alone had no effect and even partially reversed the antiviral effect of IFN-α. Notably, the target cells of FV are granulocytes, monocytes, lymphocytes, and erythroid precursor cells (33). Thus, we reasoned that in the FV model, IL-7 might activate lymphocytes and thereby possibly generate more FV target cells for virus infection. To address this issue, we postponed the cytokine treatment for 4 d, to day 7. However, this change in the experimental design did not result in an effect of IL-7 alone or combined with IFN-α on viral replication. Furthermore, the frequency of FV-specific CD8+ T cells was not augmented by IFN-α and/or IL-7 (data not shown).
A number of differences in the experimental design might explain the conflicting results of our group and those of Lu's group, who reported an antiviral effect with IL-7 alone. First, in the latter study, a different mouse strain (DBA/2 mice) was used. Of note, the authors showed that these mice have substantially lower NK cell activity, which is restored by IL-7 treatment (35). Second, Lu et al. used a different preparation of IL-7, and a different dosage and application schedule. Finally, numerous mouse-passaged FV isolates were recently reported to also contain LDV, and co-infection with LDV was shown to delay FV-specific CD8+ T-cell responses during acute infection (45). Thus, LDV contamination might have contributed to the beneficial effects of IL-7 seen in the study by Lu et al.
In conclusion, while we could not show an antiviral effect of IFN-α and/or IL-7 in our chronic LCMV infection model with cytokine treatment started during the early replication phase, and of IL-7 in acutely FV-infected mice, it may well be that this cytokine combination would inhibit the replication of other viruses, and be of therapeutic benefit in other infection models. In support of this hypothesis, Parker et al. recently reported that IFN-α therapy-induced lymphopenia in SIV-infected rhesus macaques is counteracted by IL-7, suggesting that addition of IL-7 to the standard IFN-α/ribavirin therapy in HIV/HCV co-infected patients may be beneficial (46). Thus, it might be worth testing the addition of IL-7 to IFN-α in other in vivo models of chronic viral infection.
Footnotes
Acknowledgments
We thank Andreas Bergthaler and David Pinschewer for help with establishing the LCMV model and providing LCMV gp33, Maya Borer (Roche) for providing Pegasys, Yvonne Meier (Schering-Plough) for providing PegIntron, Cytheris for providing IL-7 and critically reading the manuscript, and Pal Johansen for help with the IV infections and cytokine treatments.
A.A. was supported by the Theodor and Ida Herzog-Egli foundation. U.H. was supported by the Swiss National Science Foundation (SNSF) with fellowship 323530-123717. R.F.S. was supported by the SNSF #1003A-118391. This work was financially supported by Cytheris.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.