
Abstract
The absence of a suitable animal model for HIV infection is one of the major obstacles to the development of a preventive HIV vaccine. Vaccines showing good response in animal studies may fail in human efficacy trials. We have demonstrated DC-mediated in vitro sensitization of autologous T cells against three HIV constructs. The in vitro sensitized T cells were able to demonstrate a polyfunctional T-cell response, as well as central and effector memory T cells, and virus lysis in a virus inhibition assay, three potentially protective responses. However, none of the constructs could induce all three responses. Also there were variations from volunteer to volunteer. These may be due to genetic and other factors. This study provides evidence of an in vitro system that can be used to assess the immune response against a candidate vaccine, and may also provide the opportunity to modify vaccine constructs to achieve the goal of developing an ideal vaccine.
Introduction
Materials and Methods
HIV constructs
Of the three HIV constructs used in this study, multi-epitope vaccine (MEV) and pVAXgag were DNA plasmid constructs, and IVC-4 was a pseudotyped HIV, and all were developed at the National AIDS Research Institute, Pune, India.
The MEV construct was a plasmid DNA with a pVAX backbone containing gene sequences expressing 19 potential CTL epitopes from the conserved immunodominant regions of HIV-1 Gag (n=11), Nef (n=7), and Env (n=1), that were selected based on our previous study (48). The construct was designed by Dr. Bhattacharya (Department of Molecular Virology, National AIDS Research Institute, Pune, India) by selecting the common overlapping amino acid sequences of the conserved immunodominant regions spaced by flexible linker sequences. The designed construct was manufactured by Eurofins Medigenomix GmbH, Ingelheim, Germany. The pVAXgag construct contained a full-length gag gene derived from HIV-1 subtype C, isolated from a recently-infected Indian male patient with a history of heterosexual transmission (GenBank accession number AY4484419). The IVC-4 was env pseudotyped virus 4.J22 (GenBank accession number EU 908219.1) developed at Department of Molecular Virology, National AIDS Research Institute, Pune, India, which was produced by co-transfection of env+ pSVIIIenv or env+ pcDNA 3.1/V5-His-TOPO with env-defective HIV-1 backbone vector (pSG3ΔEnv) (39).
Antigenic peptides to assess the immune response
Pools of overlapping peptides (kindly donated by the National Institutes of Health [NIH] AIDS Research and Reference Reagent Program) for the Gag (cat. no. 3993), Nef (cat. no. 5189), Env (cat. no. 9499), and Pol (cat. no. 6208) regions were used to assess the immune response generated after in vitro sensitization.
Samples
PBMCs were taken from six uninfected healthy volunteers (D-1 to D-6) aged 25 to 37 y. Due to the constraints of the cell numbers, the experiments were carried out in three volunteers each for pVAXgag and IVC-4. The B-cell lines (BCLs) were developed from the PBMCs by transformation using the Epstein-Barr virus (EBV) (36), and used as antigen-presenting cells in the ELISPOT assays.
Generation of DCs
The DCs were generated from blood monocytes as previously described (42) with minor modifications. Briefly, monocytes were isolated from PBMCs using magnetic beads (human untouched monocytes kit, cat. no. 113.50D; Dynabeads, Oslo, Norway), and cultured at a concentration of 1–2×106 cells/mL in complete RPMI medium (RPMI 1640 containing 10% fetal bovine serum [FBS]; Sigma-Aldrich, St. Louis, MO; 2 mM L-glutamine, 25 mM HEPES, and penicillin and streptomycin) in the presence of GM-CSF (250 ng/mL) and IL-4 (100 ng/mL) for 5 d to derive immature dendritic cells (iDCs), which were subsequently loaded with antigens.
Antigen loading and maturation of iDCs
Both the vaccine constructs pVAXgag and MEV were transfected separately into the iDCs using the Amaxa human dendritic cell nucleofector kit (cat. no.VPA-1004; Lonza, Cologne, Germany) following the manufacturer's instructions. In the case of IVC-4, 2×106 iDCs (in 500 μL complete medium) were infected overnight with 1 mL of the IVC-4 stock (titer: 16,80,000 relative luminescence units [RLU]/mL) at 37°C in a 5% CO2 atmosphere, and washed (3×) on the following day to remove the residual virus. Infection of iDCs was confirmed by in house nested polymerase chain reaction (PCR) for the gag region using primer1-primer2 and primer3-primer4 for the first and second rounds, respectively (24). The antigen-loaded iDCs were matured using a mixture of cytokines containing IL-1β (10 ng/mL), IL-6 (100 ng/mL), TNF-α (50 ng/mL), and prostglandin-E2 (1 μg/mL) by incubating for 2 d.
Monocytes, iDCs, and mDCs specific markers
The differentiation of monocytes (on day 0) into iDCs (on day 5) and matured DCs (mDCs) (on day 7) was confirmed with a FACS Aria flow cytometer (Becton Dickinson, Franklin Lakes, NJ) using FACS Diva software. The monoclonal antibodies used were: CD14-PE (monocyte-specific marker), CD11c-PE (myeloid marker), CD83-FITC (DC-specific marker), CD86-FITC (DC-specific marker), HLA-DR-PE (DC-activation marker), CD80-FITC (DC-specific marker; BD Pharmingen, San Diego, CA), and DC-SIGN-PE (DC-specific marker; R&D Systems, Minneapolis, MN).
Expression of HIV-antigens on mDCs
Expression of the HIV antigens on the mDCs transfected with pVAXgag and MEV was confirmed using flow cytometry. The mDCs were stained with 1:10 diluted pooled HIV-1-antibody-positive sera (n=10) for 30 min at room temperature, washed (3×), and stained with anti-DC-SIGN-PE (R&D Systems) antibody, and anti-human-IgG-FITC (Bangalore Genei, Bangalore, India) conjugate (1:10 dilution) for additional 30 min. After removing the unbound antibody by washing, the cells were acquired and analyzed against non-transfected mDCs as control on a FACS Aria flow cytometer using FACS Diva software. The DC-SIGN-expressing cells (5000 events) were gated and analyzed for expression on HIV antigens. Upon transfection with pVAXgag, 7.9% and 19.3% of mDCs from D-2 and D-3, respectively, demonstrated expression of HIV antigens. The percentages of mDCs from D-1, D-2, D-3, D-4, and D-6 that were transfected using MEV constructs were 18.8, 11.2, 10.8, 15, and 4.2, respectively.
In vitro sensitization of T cells
Freshly isolated autologous PBMCs were mixed with antigen-loaded mDCs (DC:PBMC ratio of 1:10) and incubated for 10 d at 37°C in 5% CO2 in complete medium supplemented with interleukin-2 (IL-2, 20 U/mL). The cultures were replenished with fresh complete medium containing IL-2, and incubated for another 10 d. The HIV-specific response was assessed on day 20.
Assessment of the HIV-specific immune response
The antigen-specific cell-mediated response was assessed by interferon-γ (IFN-γ) secretory ELISPOT assay, and the pattern of intracellularly-secreted cytokines (ICS), in response to antigenic stimulation using flow cytometry.
IFN-γ secretion
The T-cell response was estimated as the number of IFN-γ-secreting cells in response to pools of overlapping HIV peptides (5 μg/mL of each peptide; NIH AIDS Research and Reference Reagents Program). The T cells sensitized with MEV were stimulated using the pools of peptides representing the epitopes coded for by the construct. In the case of IVC-4, the peptide pools from the Gag, Gp41, Gp120, protease, integrase, and reverse transcriptase regions were used, whereas only the Gag peptide pool was used in the case of pVAXgag. Briefly, the peptides were presented to sensitized T cells (1×105 T cells/well) using autologous B lymphoid cells (BCLs) pulsed with peptides for 2 h at 37°C in 5% CO2 at a ratio of 10:1, transferred to ELISPOT plates pre-coated with primary antibody for IFN-γ (10 mg/mL), and further incubated for 16–24 h at 37°C in 5% CO2. The following day, the IFN-γ-producing cells were visualized with a biotinylated anti–IFN-γ secondary antibody, followed by an enzyme-substrate complex (avidin-bound biotinylated horseradish peroxidase, Vectastain; Vector Laboratories, Burlingame, CA), and the chromogen substrate (AEC; Sigma-Aldrich). The spots were counted as spot-forming units (SFU)/106 T cells using an automated ELISPOT reader. The sensitized T cells without stimulation (mock) were used to estimate the background response. Other control included no cell controls and phytohemagglutinin-P (PHA-P; Sigma-Aldrich) as positive control. The response to peptides from Yersinia pestis (a kind gift from Dr. D.N. Rao, AIIMS, New Delhi) was measured to ascertain the specificity of the response. In the case of the MEV construct, peptides coded by HIV genes that are not included in the construct were used to ascertain specificity of response.
Estimation of intracellularly-secreted cytokines
Sensitized CD4+ and CD8+ T cells were analyzed for secretion of IFN-γ, TNF-α, IL-2, and perforin in response to HIV antigens. Pools of Gag, Nef, and Env peptides (in case of MEV), Gag (in case of pVAXgag), and Gag, Gp41, Gp120, protease, integrase, and reverse transcriptase (in case of IVC-4) were used. Phorbol 12-myristate 13-acetate (PMA)-ionomycin (I) was used as a positive control, while unstimulated T cells were used as negative control. PBMCs (1×106) were incubated for 2 h with peptides (2 μg/mL) at 37°C in 5% CO2, followed by the addition of brefeldin A (10 μg/mL; Sigma-Aldrich) and incubation for 4–6 h. Subsequently the cells were washed (3×) with wash buffer (PBS+0.5% FCS). The cells were then permeabilized using 200 μL of permeabilizing solution II (BD Biosciences) at room temperature for 10 min, washed (3×) with wash buffer, and stained with anti-CD3-PECy5, anti-CD4-APCCy7, anti-CD8-PETR, anti-IFN-γ-PECy7, anti-TNF-α-PE, anti-IL-2-APC, and anti-perforin-FITC antibodies (BD Pharmingen) for 30 min at room temperature. The cells were fixed using 3% formaldehyde and acquired and analyzed on a FACS Aria using FACS Diva software. The results were expressed as net percentage of cytokine-positive CD4+ or CD8+ T cells (net percentage=percentage of Ag-specific – percentage of negative control). The peptide-specific response was considered positive if the response was at least twofold higher than the negative control (33,50).
Characterization of phenotypes of sensitized T cells
Phenotypes of sensitized T cells such as effector (CD45RA+CCR7−), effector memory (CD45RA−CCR7−), central memory (CD45RA−CCR7+), and naïve (CD45RA+CCR7+), were assessed on the basis of surface expression of CD45RA (a naïve T-cell marker), and CCR7 (a cell-homing marker). The cells were stained for surface markers with CD3-FITC, CD4-PETR, CD8-APCCy7, CCR7-PECy7, and CD45RA-APC, washed three times, and fixed with 3% formaldehyde. The cells were then acquired and analyzed on a FACSAria using FACS Diva software.
HIV inhibition assay
The ability of sensitized T cells (effector) to kill HIV-infected autologous CD4+ T cells was assessed using a functional in vitro assay as described previously (11,41) with minor modifications. Freshly isolated CD4+ T cells (separated with Dynabeads), called target cells in this assay, were infected with primary HIV-1 isolate (IVC-16) at an MOI of 0.1 for 4 h. The residual virus was removed by three washings using RPMI supplemented with 2% FCS. The target cells were resuspended in complete RPMI medium containing IL-2 (50 U/mL. Then 5×104 infected/uninfected CD4+ T cells were plated in triplicate in 96-well U-bottom plates. The sensitized T cells were then added to the wells at 2:1, 1:1, and 0.5:1 effector:target ratios. The cultures were incubated for 10 d at 37°C in a 5% CO2 environment in complete RPMI medium containing IL-2. The culture supernatants were harvested on days 3, 7, and 10, and tested for p24 antigen using a commercially available ELISA kit (Vironostika; Biomerieux, Eindhoven, The Netherlands). The percent virus inhibition was calculated as [1 – (p24 in test/p24 in infected control) ×100].
Results
In vitro sensitized T cells demonstrated antigen-specific IFN-γ secretion
DC-mediated in vitro antigen presentation efficiently induced antigen-specific IFN-γ in PBMCs from all donors with variable magnitudes of response. In case of IVC-4-sensitized T cells the average frequencies (Fig. 1) of Gag-specific IFN-γ-producing T cells were lower compared to MEV- and pVAXgag-sensitized T cells. Nef- and Env-specific responses were higher in MEV-sensitized T cells than in IVC-4-sensitized T cells. The responses against the unrelated peptides (i.e., Yersinia pestis) were of low magnitude, indicating specificity of the responses.
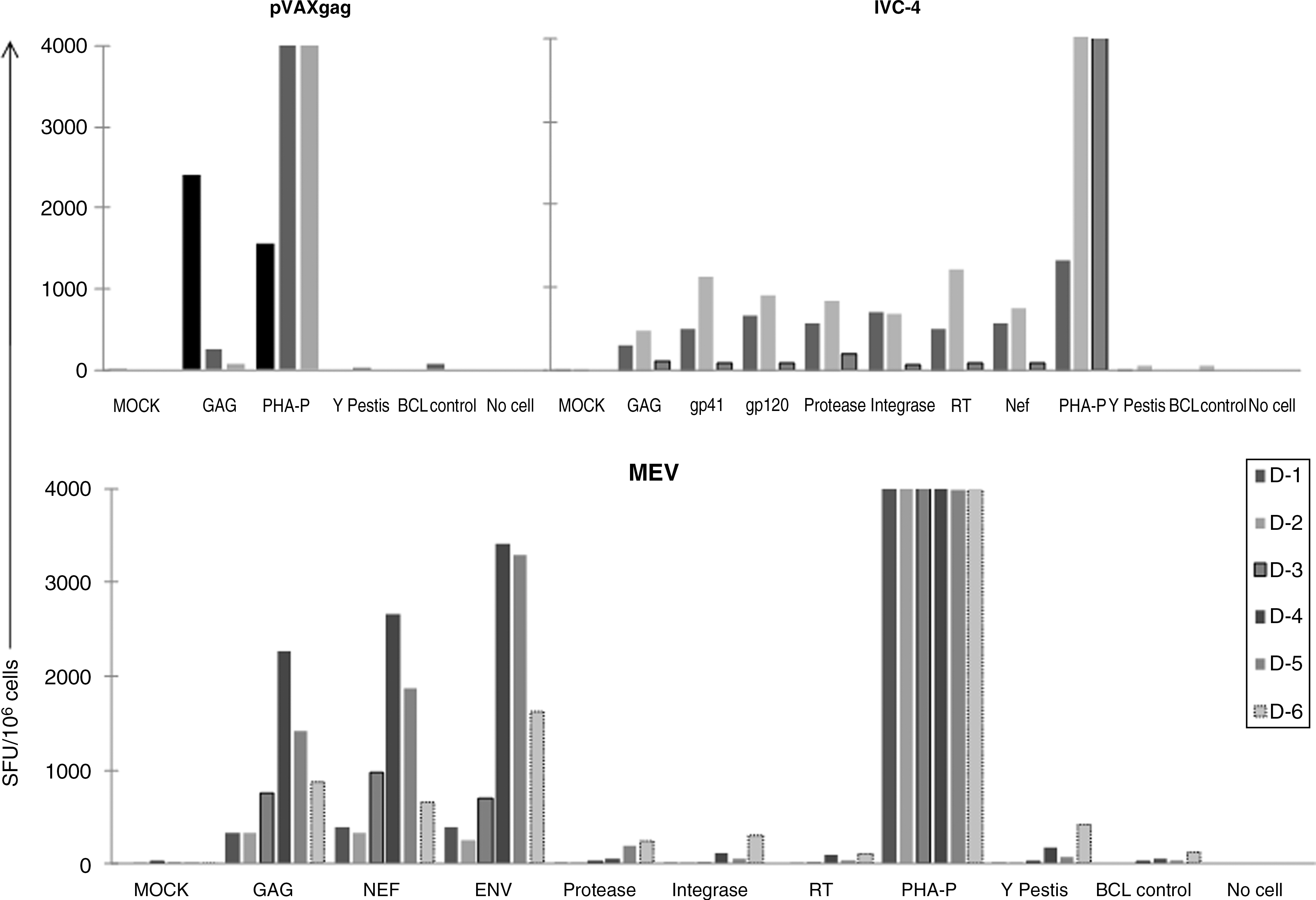
IFN-γ secretion by the sensitized T cells. The antigen-specific IFN-γ secretion was determined using an ELISPOT assay with pools of overlapping peptides from the Gag, Env, protease, integrase, reverse transcriptase, and Nef regions. MEV-, pVAXgag-, and IVC-4-sensitized T-cell responses are shown. The negative controls included MOCK (cells without any stimulation), BCL control, and No cell control, while PHA-P was used as the positive control. The unrelated response was determined against peptides from Yersinia pestis. The responses are plotted as spot-forming units (SFU) per million T cells.
ICS by in vitro sensitized CD8 T cells
Intracellular secretion of cytokines was observed predominantly in CD8+ T cells sensitized with MEV and pVAXgag, while the IVC-4-sensitized CD4+ and CD8+ T cells demonstrated intracellular secretion of cytokines (Table 1). The magnitude of the response was higher against the Gag antigen compared to the other antigens. The MEV-sensitized CD8+ T cells demonstrated better responses to nef antigen compared to IVC-4-sensitized T cells.
Secretion of cytokines by sensitized CD8 and CD4+ T cells in response to Gag, Nef, and Env peptides by the individual donor is demonstrated as percentage of cells expressing the cytokine. The range of response is denoted in the brackets.
RT, reverse transcriptase; MEV, multi–epitope vaccine; TNF–α, tumor necrosis factor–α; IFN–γ, interferon–γ; IL–2, interleukin–2.
Polyfunctional CD8+ T-cell response
The polyfunctionality of CD4+ and CD8+ T cells was determined for the secretion of any two or more of IFN-γ, TNF-α, IL-2, and perforin. A polyfunctional response was seen only in the CD8+ T cells sensitized with MEV and pVAXgag. Frequencies of MEV-sensitized polyfunctional CD8+ T cells ranged between 2.6 and 6.7% (Fig. 2). Only 2.6% of CD8+ T cells demonstrated secretion of all four cytokines. The amount of pVAXgag-sensitized CD8+ T cells secreting all four cytokines was 0.6%. No polyfunctional response was demonstrated by IVC-4-sensitized T cells.

The polyfunctional T-cell response. CD8+ T cells sensitized with pVAXgag (from all three donors, D-1, D-2, and D-3), and MEV (from donors D-1 and D-3) demonstrated secretion of multiple cytokines in response Gag peptides. The labels in the pie charts indicate the percentages of cells expressing the cytokines.
Generation of effector memory and central memory T cells upon in vitro sensitization
The MEV-sensitized CD4+ T cells and pVAXgag- and IVC-4-sensitized CD4+ as well as CD8+ T cells were of effector memory phenotype (CCR7−CD45RA−) (range 88.4–98%). Effector (CCR7−CD45RA+) CD4+ and CD8+ T cells were observed only in D-3 sensitized with pVAXgag. The presence of central memory (CCR7+CD45RA−) CD4+ T cells (range: 47.2–58.8%) was quite high in the MEV-sensitized T cells (Table 2).
Maturation phenotype as effector (E), naïve (N), effector memory (EM), and central memory (CM), of the CD4 and CD8+ T cells sensitized with MEV, pVAXgag, and IVC-4.
MEV, multi-epitope vaccine.
HIV suppression by sensitized CD8+ T cells
The pVAXgag and IVC-4 sensitized T cells from D-1 and D-3, and MEV-sensitized T cells from D-2, D-5, and D-6 were assessed in vitro for their functional ability to control HIV replication in autologous HIV-infected CD4+ T cells (Fig. 3). MEV-sensitized T cells from two donors (D-5 and D-6) demonstrated an excellent ability (>90%) to inhibit HIV replication on day 7 at all three ratios tested. The pVAXgag-sensitized T cells from D-1 and D-3 demonstrated 43% and 54% inhibition, respectively, of HIV replication on day 10 at an effector:target ratio of 2:1. The HIV inhibition by the IVC-4-sensitized T cells at effector:target ratios of 1:1 and 2:1 on day 7 and day 10 ranged from 5–32% and 36–64%, respectively.
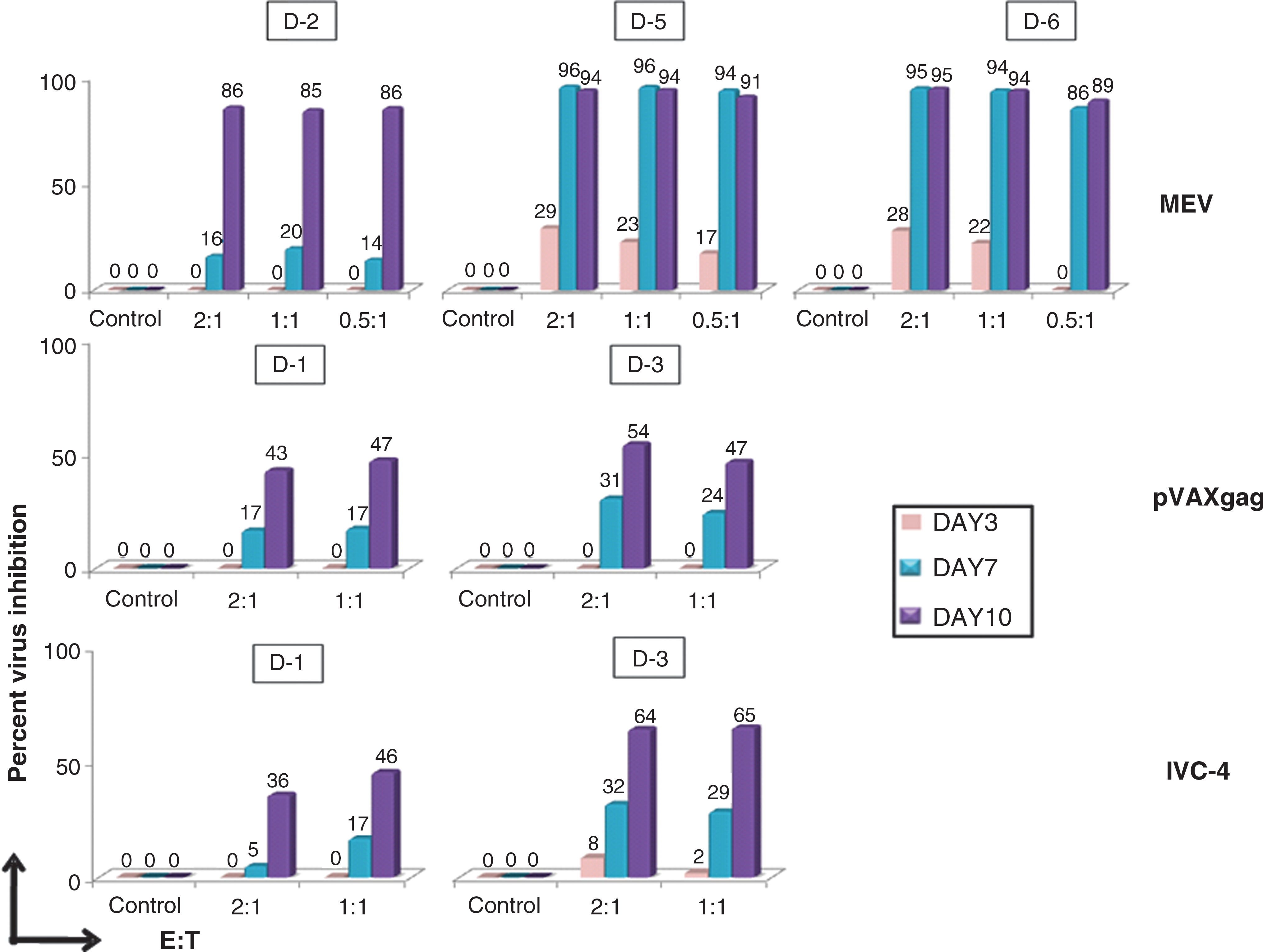
Inhibition of HIV replication by sensitized T cells. The functional ability of the sensitized T cells was determined in an in vitro assay using autologous CD8+ T cells using different E:T ratios (three in the case of MEV, and two in the case of pVAXgag and IVC-4). The culture supernatants were collected at three time points (days 3, 7, and 10) and tested for p24 antigen. The virus inhibition is plotted as the percentage of virus inhibition on the y axis. Color images available online at
Discussion
Sensitization of immune cells in vitro to generate an antigen-specific adaptive immune response has been attempted against various tumor and viral antigens with variable degrees of success (5,10,20,49,51,52). Most of these studies have used dendritic cells (DCs) as potent antigen-presenting cells (APCs) for sensitization of T cells to induce an antigen-specific immune response in vitro. DCs provide a critical interface between innate and adaptive arms of the immune system, and are well known as the only APCs capable of activating naïve T cells in vitro. As professional APCs, DCs play a central role in initiating the adaptive immune response against pathogens. A DC-based assay for in vitro sensitization of CD4+ T cells has recently been reported (34). Methods to assess the immune response in vitro would be of significant value for pre-clinical assessment of candidate vaccines. In vitro sensitization of human peripheral blood lymphocytes may provide clues about the immunogenicity of a vaccine candidate in humans. The use of such in vitro assays has been reported previously for evaluation of the human immune response against several viral and tumor antigens (4,6,21,32,49). Additionally, several in vitro studies have demonstrated the application of DCs as APCs to augment T-cell immunity to HIV infection (27,31).
We report successful in vitro sensitization of peripheral blood mononuclear cells using autologous DCs as the APCs. In the current study, three different antigenic constructs were used in the in vitro sensitization experiments and the responses generated are described. Among these three constructs, one construct represented nearly the full genome of HIV in a pseudotyped virus, and other two constructs were DNA plasmids that expressed the HIV-1C gag gene from an Indian patient with recent HIV infection (pVAXgag), and 19 HIV-1 subtype C CTL epitopes commonly recognized by HIV-infected Indian individuals (MEV) (48). We have used plasmid DNA constructs that were transfected into DCs using a high-throughput transfection system, which allows the DNA to directly enter the nucleus. This system has the advantage of faster and improved expression of viral proteins. IVC-4, the pseudotyed HIV, was in itself a means of delivery system, since it can undergo only one replication cycle and cannot produce progeny viral particles, but still expresses the HIV proteins on the infected cell.
The in vitro sensitization using a DC/T-cell co-culture assay used in our studies demonstrated HIV-specific T-cell responses in nearly all donors. However, the magnitude of the response was variable, which may be attributable to the genetic make-up of the donor affecting T cells and/or APC function, and thus could represent a natural phenomenon.
It has become increasingly evident that rather than the magnitude, the quality of the immune response is a crucial factor in determining the protective response (46). The immunogenicity of HIV vaccines is measured by the number of antigen-inducing IFN-γ-secreting cells in the respondents; however, recent evidence has suggested that the number of polyfunctional T cells may be a better indicator. Recently an HIV vaccine candidate has shown induction of polyfunctional T cell responses in healthy volunteers (16,18). Vaccine-induced polyfunctional T cells have been shown to provide protection against various pathogens (1,9,15,30,38). HIV-infected long-tem nonprogressors (LTNP) have also demonstrated higher frequencies of polyfunctional CD8+ T cells than progressors (3). Additionally, strong polyfunctional CD4+ and CD8+ T-cell responses in blood and rectal mucosa have been recently reported in HIV elite controllers (12,13). It has also been shown that patients who maintain CD4+ and CD8+ T cells that have the capacity to produce both IFN-γ and IL-2 typically experience a milder HIV disease course than those who have a greater proportion of single-cytokine-positive responses (19,53). The two DNA vaccine constructs pVAXgag and MEV induced polyfunctional CD8+ T cells.
In this study, we observed development of effector memory CD4+ T cells as the main population by all three HIV constructs. Surprisingly, the MEV construct alone demonstrated development of central memory CD4+ T cells. Memory T cells are important for generating long-term immunity and vaccine-induced viral control. Two distinct phenotypes of memory T cells, effector memory (TEM) and central memory (TCM) cells (44,45), have discrete functions. The effector memory T cells migrate to the inflamed peripheral tissues and display immediate memory function (25,43), while central memory T cells that reside in the secondary lymphoid organs are thought to ensure the long-term maintenance of antiviral responses due to their long half-life and self-renewal capacity (26,28). Correlation of effector memory and central memory T cells with plasma viral load have also been demonstrated (37). DNA vaccines have been shown to generate long-lived central and effector memory T cells (29,40). Hence vaccines that are designed to induce such long-lasting memory T cells are important. In our study, the MEV construct demonstrated development of central memory CD4+ T cells, which may imply that a vaccine could induce long-term protection against HIV. Recently, Rosa et al. reported vigorous polyfunctional and long-lasting T-cell responses against a multi-epitope DNA vaccine in mice (40). A Phase IIb vaccine trial using DNA prime (including envA, envB, envC, gagB, polB, and nefB) that was boosted with recombinant adenovirus serotype-5 (rAd5) HIV-1 with matching inserts also reported the generation of polyfunctional T-cell responses in healthy volunteers (7). The multi-epitope DNA vaccine construct MEV used in our study also demonstrated generation of polyfunctional T-cell responses.
Recent technological advancements have enabled more quantitative and detailed analysis of virus-specific immunity in humans. However, assays to assess the antiviral activity of such responses are needed. Fauce et al. (11) reported a simple and reproducible in vitro autologous co-culture assay that provides a practical method to evaluate the HIV-specific effector function of CD8+ T cells. In addition, use of bi-specific antibodies for expansion of CD8+ T cells makes this assay feasible for use in situations where there are limited cell numbers. We applied this assay to study the ability of in vitro-sensitized T cells to suppress viral replication in autologous CD4+ T cells.
All three constructs were able to induce sensitized T cells with the ability to suppress viral replication. To our knowledge, we have shown for the first time that these in vitro-sensitized CD8+ T cells can lyse virally-infected CD4+ T cells, indicating the functionality of the response.
All three HIV constructs were able to sensitize autologous T cells in vitro, and the nature of the response varied from donor to donor and from construct to construct (Table 3). However, further studies on larger numbers of samples would be helpful in drawing more solid conclusions. The presence of central and peripheral memory cells and polyfunctional T cells, and the ability to suppress virus growth are considered important in the control of the virus in vivo. However, none of the vaccine constructs could induce all three types of responses: (1) the MEV demonstrated generation of central memory T cells, (2) the pVAXgag and MEV demonstrated generation of polyfunctional T-cell responses, and (3) IVC-4 demonstrated responses by CD4+ as well as CD8+ T cells. The MEV construct that contained CTL epitopes demonstrated the generation of CD4+ central memory T cells. Several studies have reported various factors that are associated with the development of CD8+ T-cell memory responses (35,54). In contrast, very few studies have addressed the requirements for the generation of CD4+ T-cell memory responses (17,47). One possibility that might explain the generation of CD4+ central memory T cells is that the CTL epitopes had overlapping sequences and contained long stretches of peptides. These long stretches of peptides might have been processed and presented by the MHC class II pathway by DCs, and helped generate CD4+ central memory responses. However, further exploratory studies are needed to confirm this.
Antigen-specific IFN-γ secretion, intracellular cytokine secretion by T cells with polyfunctional response, the phenotype of sensitized T cells as effector memory (EM) or central memory (CM) cells, and the ability of sensitized T cells to inhibit HIV replication by all three constructs.
DC, dendritic cell; MEV, multi-epitope vaccine; IFN-γ, interferon-γ; IL-2, interleukin-2.
Although the correlates of protection have not been identified in HIV infection, based on the evidence accumulated from numerous studies, an ideal vaccine should be able to induce T-cell responses that are polyfunctional in nature and long-lasting, as well as efficient in the suppression of viral replication. None of the three constructs used here induced all the desired responses (i.e., polyfunctional T cells, virus suppression, and generation of central memory T cells). It may be possible to fine tune these constructs to achieve this. Alternatively, the use of two constructs together or in a prime and boost strategy may help in achieving this. The novel in vitro sensitization system used in our study may provide an opportunity to modify vaccine constructs to achieve the goal of developing an ideal vaccine. The in vitro sensitization may also provide evidence that will influence decision making about when to move vaccines from animal to human clinical trials.
Footnotes
Acknowledgments
We appreciate the help of Dr. Srikanth Tripathy at the Department of Clinical Sciences, National AIDS Research Institute, for providing us the pVAXgag construct. We thank Dr. D.N. Rao for giving us the Yersinia pestis peptides. We acknowledge the NIH AIDS Research and Reference Reagent Program for providing the Gag (cat. no. 3993), Nef (cat. no. 5189), Env (cat. no. 9499), and Pol (cat. no. 6208) peptides.
Author Disclosure Statement
No competing financial interests exist.