
Abstract
Human T-cell leukemia virus type I (HTLV-I), a causative agent of adult T-cell leukemia (ATL), is transmitted from mother to child, predominantly by breastfeeding. Oral HTLV-I infection and infection early in life are associated with a subsequent risk of ATL. Although the pathogenic mechanisms of ATL remain largely unknown, the host immune system seems to play an important role in HTLV-I pathogenesis. Previous studies have shown that monocytes from ATL patients had reduced capacity for dendritic cell (DC) differentiation. Therefore, we performed the present study to clarify the mechanisms responsible for the impairment of DC differentiation using HTLV-I-infected breast milk macrophages (HTLV-BrMMø). We found that when CD14+ monocytes were cultured with GM-CSF and IL-4 in the presence of HTLV-BrMMø, they altered the surface phenotype of immature DCs and the stimulatory capacity of T-cell proliferation. The presence of HTLV-BrMMø significantly blocked the increased expression of CD1a, CD1b, CD11b, DC-SIGN, and HLA-DR; however, increased expression of CD1d and CD86 was observed. These effects could be partially replicated by incubation with culture supernatants from HTLV-BrMMø. The impairment of monocyte differentiation might be not due to HTLV-I infection of monocytes, but might be due to unknown soluble factors. Since other HTLV-I-infected cells exhibited similar inhibitory effects on monocyte differentiation to DCs, we speculated that HTLV-I infection might cause the production of some inhibitory cytokines in infected cells. Identifying the factors responsible for the impairment of monocyte differentiation to DCs may be helpful to understand HTLV-I pathogenesis.
Introduction
Although the pathogenic mechanisms of ATL remain largely unclear, HTLV-I-encoded gene products such as Tax or basic leucine zipper factor (HBZ) may be among the key factors ultimately leading to the development of ATL (23). Moreover, the host immune system also seems to play an important role in HTLV-I pathogenesis. It has been shown that HTLV-I-specific T-cell responses are critical for a low viral load to be maintained (7,20). However, HTLV-I-specific cytotoxic T cells (CTLs) can be induced from peripheral blood mononuclear cells (PBMCs) of asymptomatic carriers, but rarely from those of ATL patients or those who fail to respond to ex vivo stimulation (4). Clarifying the reasons why HTLV-I-specific immune responses are not effective in HTLV-I-infected patients is critical to prevent the development of ATL.
Dendritic cells (DCs) are professional antigen-presenting cells (APCs) involved in the capture and processing of antigens and are capable of inducing primary responses of T cells. Therefore, the modifications of DCs during their differentiation process have a direct effect on T-cell responses. Previous studies have shown that monocytes from ATL or HAM/TSP patients had a reduced capacity for DC differentiation in vitro (1,22,29), and the numbers of myeloid DC (myDC) and plasmacytoid DC (pcDC) cells were decreased in ATL patients (15). HTLV-I can efficiently infect monocytes and DCs as well as T cells (19); however, Azakami et al. have reported that there was no close correlation between a high proviral load of PBMCs and the capacity for DC differentiation (5). Furthermore, monocytes that exhibited impaired differentiation to DCs were not always positive for HTLV-I (29). These findings suggest that impaired differentiation of DCs might be induced under infectious disease conditions rather than the infected state of monocytes.
HTLV-I is transmitted primarily in three ways: mother to child (11), sexual transmission (40), and via blood transfusion (38). Among these routes, mother-to-child transmission through breast milk is the most important route to stop, because oral HTLV-I infection and infection early in life are associated with a subsequent risk of ATL (11,14). Previously, we demonstrated that CD14+ breast milk macrophages (BrMMø), the majority of cells in breast milk, particularly in early colostrum, were capable of producing granulocyte macrophage-colony stimulating factor (GM-CSF) spontaneously, and differentiating into CD1+ DCs by stimulation with exogenously added interleukin-4 (IL-4) (16,32,39). Moreover, they could differentiate into DC-like cells by HTLV-I infection (36). An appropriate number of HTLV-I-infected BrMMø efficiently induced T-cell proliferation and transmitted HTLV-I virions to activated T cells (36); that is, HTLV-I-infected BrMMø might serve as a reservoir as well as a transmitter of HTLV-I. Recently, we observed that when CD14+ monocytes were cultured with GM-CSF and IL-4 in the presence of HTLV-I-infected BrMMø, they altered the surface phenotype of immature DCs and the stimulatory capacity of T-cell proliferation. This inhibitory effect could be partially replicated by incubation with culture supernatants from HTLV-I-infected BrMMø. In the present study, considering the possibility that HTLV-I-infected BrMMø might modulate the immune systems in newborns through breastfeeding, we examined how HTLV-I-infected BrMMø affect the process of monocyte differentiation to DCs.
Materials and Methods
Preparation of cells
Fresh BrMMø were isolated from breast milk by Lymphosepar I (Immuno-Biological Laboratories, Gunma, Japan) gradient centrifugation, followed by adherence to polystyrene tissue culture dishes for 1 h at 37°C. Adherent cells were then removed by incubation with 5 mM ethylenediaminetetraacetic acid (EDTA) for 30 min at 4°C and cultured in T-cell culture medium (CTM) (35), composed of RPMI 1640 medium supplemented with 2 mmol/L L-glutamine, 1 mmol/L sodium pyruvate, 0.1 mmol/L non-essential amino acids, a mixture of vitamins, 1 mmol/L HEPES, 100 U/mL penicillin, 100 μg/mL streptomycin, 50 mmol/L 2-mercaptoethanol, and heat-inactivated 10% fetal calf serum. PBMCs were isolated from healthy donors using Lymphosepar I gradient centrifugation. CD14+ monocytes were separated from PBMCs by a human monocyte enrichment kit (Stemcell Technology, Vancouver, B.C., Canada), routinely resulting in >90% purity of CD14+ cells. Cells were cultured in CTM supplemented with 200 ng/mL GM-CSF and 10 ng/mL IL-4 to obtain DCs. HTLV-I-producing cell lines of BrMMø (HTLV-BrMMø) and DC (HTLV-DC) were established as described previously (36). Briefly, BrMMø or immature DCs were infected with 0.5 mL supernatant containing 354 ng/mL HTLV-I p19 from MT-2 (26), an HTLV-I-producing T-cell line, for 18 h at 37°C, and further propagated with CTM. B95-a cells [an Epstein-Barr virus-transformed marmoset B cell line (21)], MOLT-4 cells [human acute lymphoblastic leukemia cell line (25)], and MT-2 cells were cultured in CTM. This study was approved by the Review Board of Nippon Medical School and all human participants gave written informed consent.
Co-culture experiments
CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 either in the presence or absence of the following irradiated (30 Gy) test cells: HTLV-BrMMø (36), MT-2 (26), HTLV-DC, B95-a (21), and MOLT-4 (25), at a 30:1 monocyte:test cell ratio for 3 d. Test cells were labeled with 2×10−6 M PKH67 (green fluorescence) or PKH26 (red fluorescence) (Sigma-Aldrich, St. Louis, MO) to differentiate them from monocytes. Titration of HTLV-BrMMø to monocyte cell ratio of 1:10 up to 1:90 was performed using different numbers of HTLV-BrMMø to examine whether the inhibitory effect of HTLV-BrMMø on monocyte differentiation occurs in a dose-dependent manner. To obtain culture supernatants from various cell lines, 3×104 cells/mL of each cell line were cultured in CTM for 7 d and the supernatants were collected after centrifugation at 500 g for 10 min. CD14+ monocytes were cultured with GM-CSF and IL-4 either in the presence or absence of culture supernatants for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4 for 2 d. Purified HTLV-I virions were prepared from culture supernatants from HTLV-BrMMø (HTLV-BrMMø-sup) using the Fast-Trap lentivirus purification and concentration kit (Millipore, Billerica, MA) according to the manufacturer's instructions. To examine the characteristics of the active substances in HTLV-BrMMø-sup, we heated HTLV-BrMMø-sup to 90°C for 30 min, and subjected HTLV-BrMMø-sup to treatment with 1500 U/mL trypsin (Sigma-Aldrich) at 37°C for 3 h. To determine the molecular weight of the active substance in HTLV-BrMMø-sup, they were fractionated by Amicon Ultra-15 (Millipore).
Antibodies and flow cytometric analysis
Cells were pelleted and re-suspended at a concentration of <5×105 cells in 50 mL PBS with 0.1% NaN3 and 1% human serum containing each monoclonal antibody. For flow cytometric analysis, phycoerythrin (PE)-conjugated antibodies to CD1a, CD1d, CD11b, CD14, CD83, and CD86, as well as fluorescein isothiocyanate (FITC)-conjugated antibody to HLA-DR were purchased from BD Bioscience (Franklin Lakes, NJ). PE-conjugated antibodies to CD1b and CD1c, and allophycocyanin (APC)-conjugated antibody to CD3, were from BioLegend (San Diego, CA). PE-conjugated antibodies to CD40 and DC-SIGN were from eBioscience Inc. (San Diego, CA) and R&D Systems (Minneapolis, MN), respectively. FITC-conjugated antibody to Vα24 was from Immunotech (Marseille, France). After 30 min incubation with each antibody at 4°C, the cells were washed and re-suspended in PBS for analysis by FACScan (BD Bioscience) and Flow Jo analysis software (Tree Star Inc., Ashland, OR). Neutralizing antibodies against IL-10 were purchased from Miltenyi Biotec Inc. (Auburn, CA).
Detection of HTLV-I proviral DNA
CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence of HTLV-BrMMø-sup for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4 for 3 d. DNA was isolated from monocytes using Blood & Tissue Genome Mini kits (Viogene, Sunnyvale, CA). HTLV-I proviral DNA was detected by polymerase chain reaction (PCR) using the following primers for the Tax gene to generate a 244-bp fragment: 5′-TGTTTGGAGACTGTGTACAAGGCG-3′ and 5′-GTTGTATGATTGGCGGGGTAA-3′.
Mixed lymphocyte reaction (MLR) assays
For assessment of T-cell stimulation in allogeneic MLR, both PBMCs and CD14+ monocytes were cultured with HTLV-BrMMø-sup for 18 h. After washing with PBS, cells were further incubated in CTM supplemented with GM-CSF and IL-4 for 2 d. The cells were then irradiated (30 Gy) and used as APCs. Purified allogeneic T cells (5×104 cells/well) were cultured with the indicated number of APCs in 96-well U-bottomed tissue culture plates at 37°C for 5 d. Proliferation was measured by the addition of 0.5 μCi/well 3H-thymidine/well for the last 16 h, and the plates were harvested and counted using a β-counter (1450 Microbeta Trilux; Wallac, Gaithersburg, MD).
Measurement of virus and cytokine production
IL-10 and IL-12 concentrations were measured using the DuoSet ELISA Development Kit (R&D Systems) according to the manufacturer's instructions. HTLV-I p19 core antigens were measured using the HTLV p19 Antigen ELISA Kit (ZeptoMetrx Corporation, Buffalo, NY).
Induction of iNKT cells
PBMCs were cultured with 10 ng/mL α-galactosylceramide (α-GalCer; Enzo Life Sciences Inc., Farmingdale, NY) either in the presence or the absence of HTLV-BrMMø-sup for 7 d. Cell cultures were fed by replacing a half-volume of media every other day with fresh CTM with IL-2 (10 U/mL). No additional α-GalCer was added during cell feeding.
Statistical analysis
Student's t-test was used to determine the significance of differences in means. A p value of<0.05 was considered significant.
Results
HTLV-BrMMø and supernatants from HTLV-BrMMø inhibit monocyte differentiation to immature DCs
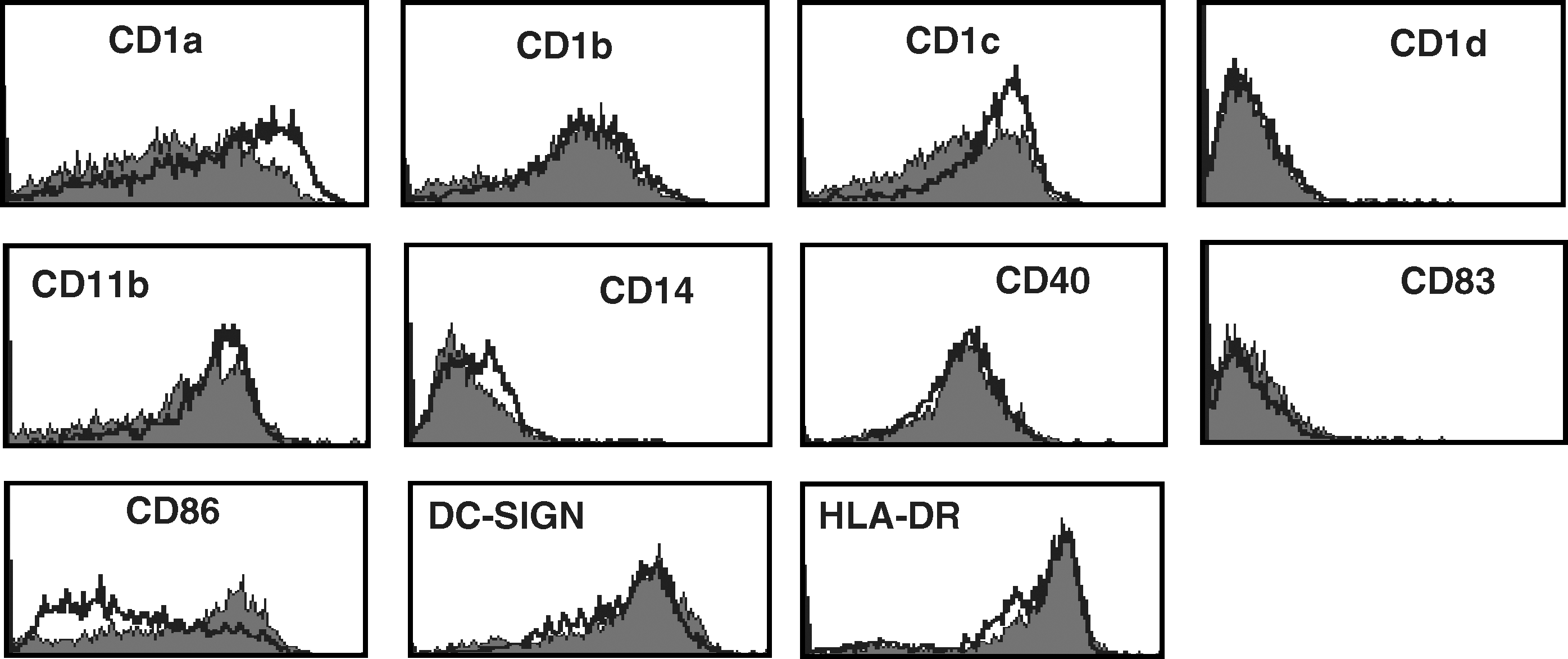
In order to study whether HTLV-BrMMø modulates DC function by regulating the expression of surface molecules, CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence or absence of HTLV-BrMMø for 3 d. Monocytes were characterized by side scatter and forward scatter profiles, followed by gating on PKH-negative cells by FACS analysis. Generally, when CD14+ monocytes are stimulated with GM-CSF and IL-4, they differentiate into immature DCs, displaying increased expression of CD1a, in contrast to decreased or absent CD14 expression. The presence of HTLV-BrMMø significantly blocked the increased expression of CD1a, CD1b, CD11b, DC-SIGN, and HLA-DR, and slightly blocked that of CD1c. Interestingly, increased expression of CD1d and CD86 was observed. The expression levels of CD14 and CD40 were not altered (Fig 1A). This inhibitory effect was not found in co-cultures with uninfected BrMMø (Fig. 1B). When CD14+ monocytes and HTLV-BrMMø were separated by a permeable filter, monocytes failed to acquire CD1a expression (data not shown). These findings suggest that HTLV-BrMMø are capable of modulating the expression of surface molecules on monocytes by secreting cytokines; therefore, we next examined the effect of culture supernatants from HTLV-BrMMø (HTLV-BrMMø-sup) on monocyte differentiation to DCs. CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence or absence of HTLV-BrMMø-sup for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4. Flow cytometric analysis performed on day 3 of culture revealed a significantly decreased expression of CD1a, DC-SIGN and HLA-DR, and increased expression of CD1d and CD86, as observed in co-cultures with HTLV-BrMMø; however, the expression levels of CD1b, CD1c, and CD11b were not altered (Fig. 1C). This inhibitory effect was not found in culture supernatants from uninfected BrMMø (Fig. 1D). These data implied that HTLV-BrMMø were capable of modulating the expression of surface molecules on monocytes, mainly through cell–cell interactions, and partly by secreting cytokines. HTLV-BrMMø-sup induced the dose-dependent inhibition of CD1a expression, and complete loss of CD1a expression resulted after more than 18 h of incubation with HTLV-BrMMø-sup (Fig. 1E).

Effect of HTLV-BrMMø and culture supernatants from HTLV-BrMMø on monocyte differentiation. CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4, (
Characterization of soluble factors responsible for the impairment of monocyte differentiation
Culture supernatants of monocytes with HTLV-BrMMø contained less than 10 ng/mL HTLV-I p19 (Fig 2A), whereas HTLV-BrMMø-sup contained more than 50 ng/mL HTLV-I-p19. HTLV-I can infect monocytes and DCs as well as T cells. Thus, to explore whether the impairment of monocyte differentiation to DCs could be caused by HTLV-I infection of monocytes, we measured HTLV-I proviral load in monocytes by PCR. Although monocytes incubated with HTLV-BrMMø-sup exhibited downregulation of CD1a, genomic DNA from these monocytes did not contain HTLV-I proviral DNA (Fig. 2B). To further examine whether HTLV-I virions were required for the impairment of monocyte differentiation to DCs, experiments were performed in which excessive amounts of purified HTLV-I virions were added to CD14+ monocytes cultured with GM-CSF and IL-4. One μg/mL of purified HTLV-I virions did not cause a loss of CD1a expression (Fig. 2C). Furthermore, when HTLV-BrMMø-sup was treated with ultraviolet irradiation or heating, and the expression levels of CD1a on monocytes remained downregulated. These findings suggest that the impairment of monocyte differentiation does not necessarily require HTLV-I infection of monocytes.
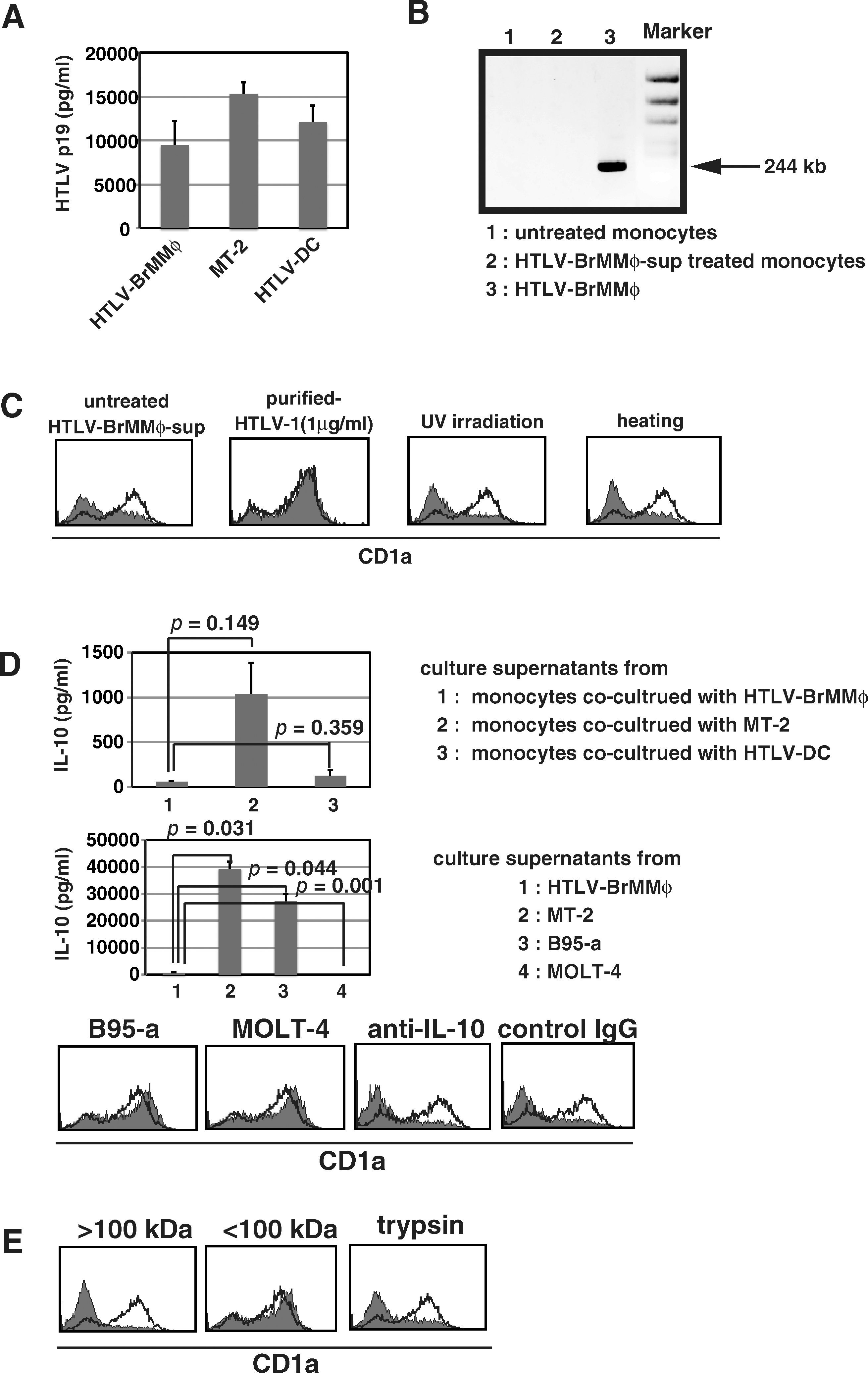
Characterization of soluble factors responsible for the impairment of monocyte differentiation. (
IL-10 is a potent immunosuppressive cytokine that reduces the expression of MHC class II, CD80, and CD86 on DCs, and inhibits the production of proinflammatory cytokines by DCs (2). Monocytes and macrophages are known to produce IL-10 by various stimuli, and Makino et al. have reported that HTLV-I-infected monocytes secreted a large amount of IL-10 (22). Co-cultures of monocytes with irradiated HTLV-BrMMø or HTLV-DC produced less IL-10 than co-cultures of monocytes with irradiated MT-2 for 3 d (Fig. 2D, upper panels). We then measured IL-10 in the 7-d-cultured supernatant of HTLV-BrMMø, in which we found far less IL-10 than in supernatants from 7-d-cultured MT-2 or B95-a (Fig. 2D, middle panels). Nevertheless, the 7-d-cultured supernatant of HTLV-BrMMø containing an undetectable amount of IL-10 significantly inhibited GM-CSF- and IL-4-mediated induction of CD1a expression, although culture supernatants from B95-a, containing more than 30 ng/mL of IL-10, did not inhibit CD1a expression (Fig. 2D, lower panels). As far as we have determined, more than 50 ng/mL of recombinant IL-10 was required to suppress CD1a expression (data not shown). Furthermore, anti-IL-10 neutralizing antibody could not restore CD1a expression (Fig. 2D, lower panels). These observations indicate that IL-10 might be not responsible for the impairment of monocyte differentiation seen in our systems. The inhibitory effect was observed in the high-molecular-weight fraction (>100 kDa) in HTLV-BrMMø-sup, and was not abolished when culture supernatants were heated to 90°C for 30 min or treated with trypsin (Fig. 2C and E), indicating that the active substance is heat- and trypsin-stable.
Effect of HTLV-BrMMø-sup on the function of APCs
Primary T-cell responses are required for the interaction between T cells and APCs. Thus we tested the effect of HTLV-BrMMø-sup on the stimulatory capacity of APCs for T-cell proliferation in an MLR. When monocytes or PBMCs were treated with or without HTLV-BrMMø-sup and used as a source of APCs, the stimulatory activity of both cells was significantly reduced (Fig. 3A).
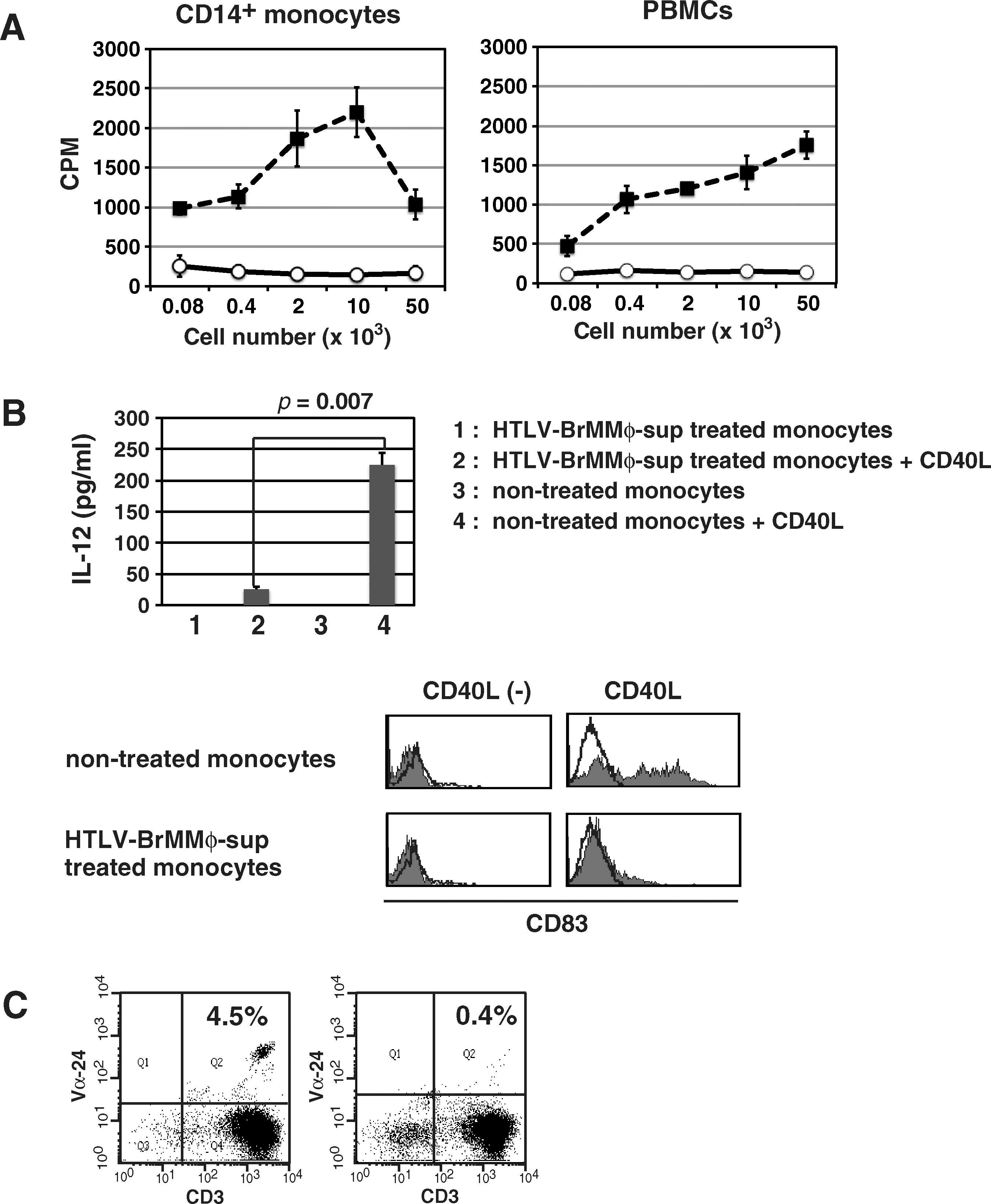
Effect of HTLV-BrMMø-sup on the function of APCs. (
Immature DCs can produce large amounts of cytokines, such as IL-6 and IL-12, when stimulated with LPS or CD40 ligand (CD40L). IL-12 is the most important cytokine in the initiation of Th-1 immunity to virus infection; therefore, monocytes treated with or without HTLV-BrMMø-sup were tested for IL-12p70 production after CD40L stimulation (30 ng/mL; PeproTech, Rochy Hill, NJ). CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence or absence of HTLV-BrMMø-sup for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4 for 7 d, and then stimulated with CD40L. After 2 d, we harvested culture supernatants and measured the level of IL-12p70 by ELISA. It was found that untreated monocytes showed increased secretion of IL-12p70 and upregulation of CD83 in response to CD40L stimulation, while CD40L-induced production of IL-12 and upregulation of CD83 was significantly suppressed in HTLV-BrMMø-sup-treated monocytes (Fig. 3B).
It was observed that the expression level of CD1d on monocytes co-cultured with both HTLV-BrMMø and HTLV-BrMMø-sup was slightly upregulated. Invariant natural killer T (iNKT) cells are a subset of lymphocytes that bridge the innate and adaptive immune systems and are potent regulators of diverse immune responses. iNKT cells can become activated directly via engagement of invariant TCR with glycolipid antigens loaded onto CD1d molecules on APCs, or indirectly via activated APCs; therefore, we examined the effect of HTLV-BrMMø-sup on the stimulatory capacity of APCs for the induction of CD1d-restricted iNKT cells by α-GalCer. PBMCs were cultured in CTM with α-GalCer in the presence or absence of HTLV-BrMMø-sup for 7 d and further cultured with IL-2 for additional 7 d. As shown in Fig. 3C, contrary to our expectations, the percentage of Vα24+ CD3+ cells was significantly decreased in HTLV-BrMMø-sup-treated PBMCs.
Effect of culture supernatants from MT-2 and HTLV-DCs on monocyte differentiation to immature DCs
Co-culture of monocytes with both uninfected BrMMø and culture supernatants from uninfected BrMMø did not affect monocyte differentiation to DCs (Fig. 1B and D). It was speculated that HTLV-I infection of BrMMø might lead to the production of inhibitory factors; therefore, we further tested the ability of culture supernatants to inhibit monocyte differentiation to DCs in other HTLV-I-infected cells, MT-2, and HTLV-DCs. Predictably, both culture supernatants from MT-2 (Fig. 4A) and HTLV-DCs (Fig. 4C) exhibited the same inhibitory effect as those of HTLV-BrMMø on the expression level of surface molecules. These inhibitory effects were not found in culture supernatants from the HTLV-I-uninfected T-cell line, MOLT-4 (Fig. 4B), or immature DCs (iDCs) (Fig. 4D).

Effect of culture supernatants from MT-2 and HTLV-DCs on monocyte differentiation to immature DCs. CD14+ monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence or absence of culture supernatants from MT-2 (
Effect of HTLV-BrMMø-sup on differentiated DCs
It was important to evaluate whether HTLV-BrMMø-sup also affected already-differentiated DCs. We prepared iDCs by culturing CD14+ monocytes in the presence of GM-CSF and IL-4 for 7 d, and then cultured with HTLV-BrMMø-sup for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4 for 2 d. As shown in Fig. 5, treatment of iDCs with HTLV-BrMMø-sup resulted in little significant alteration of the expression of most surface molecules; however, the expression level of CD86 remained upregulated. These data implied that HTLV-BrMMø-sup affected an early process in monocyte differentiation to DCs.
Effect of HTLV-BrMMø-sup on differentiated DCs. Immature DCs (iDCs) were prepared by culture of CD14+ monocytes in the presence of GM-CSF and IL-4 for 7 d with or without HTLV-BrMMø-sup for 18 h. After washing with PBS, the cells were further cultured in CTM supplemented with GM-CSF and IL-4 for 3 d.
Discussion
DCs play a critical role in initiating and regulating immune responses by promoting antigen-specific T-cell activation. The ability of DCs to initiate an immune response depends on their transition from antigen-processing to antigen-presenting cells by upregulating MHC class II and T-cell co-stimulatory molecules on the cell surface (24). This transition is an important process in mounting an immune response because immature DCs not only fail to prime T cells effectively, but also serve to promote tolerance induction (34). Makino et al. have reported that monocyte-derived immature DCs donated by ATL patients were downregulated in the expression of HLA-DR, CD1a, and CD86 (22). Several other studies have also shown the impairment of DC differentiation in HTLV-I-infected individuals. In our in vitro study, we showed that co-culture of monocytes with HTLV-I-infected cells inhibited monocyte differentiation to DCs. This inhibitory effect could be partially recapitulated by incubation with HTLV-BrMMø-sup. Factors leading to DC dysfunction are postulated as follows: first, the direct HTLV-I infection of monocytes; second, the presence of HTLV-I proteins that might modulate DC differentiation; and third, the production of cytokines from HTLV-I-infected cells that might modulate DC differentiation.
First, we tested whether impairment of DC differentiation is required for HTLV-I infection of monocytes. When monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence of UV-irradiated or heated HTLV-BrMMø-sup, the expression levels of CD1a on monocytes remained downregulated. Moreover, we could not detect HTLV-I proviral DNA in any of the samples of monocytes that exhibited impaired differentiation to DCs; therefore, we assumed that the impairment of DC differentiation does not necessarily require HTLV-I infection of monocytes.
The viral protein Tax is known to play a critical role in the genesis of ATL by functioning as a transcriptional activator and an oncoprotein (13). Tax is required for viral replication and is of special importance with respect to HTLV-I-specific immune responses. Jain et al. have shown that Tax promoted the production of proinflammatory cytokines and chemokines involved in the DC activation process. Consequently, Tax can induce the expression of CD40, CD80, CD83, and CD86 on DCs, and enhance the T-cell proliferation capability of DCs (17); therefore, Tax appears not to be directly involved in the impairment of DC differentiation. The HTLV-I accessory protein p12I has been shown to downmodulate the surface expression of HLA-ABC, ICAM-1, and ICAM-2 (6,18). We elevated the expression levels of p12I in CD1a-expressing cells using p12I expression vector (kindly provided by Dr. Franchini); however, no substantial differences were found in the expression level of CD1a between p12I expression vector-transfected cells and control vector-transfected cells (data not shown).
Regarding the impairment of DC differentiation, some studies have proposed mechanisms by which soluble products from various types of cells interfere with DC differentiation. Cytokines produced by tumor cells such as IL-1β, IL-6, IL-10, and vascular endothelial growth factor (VEGF), are known to suppress DC development and function (12,27). Spaggiari et al. have reported that mesenchymal stem cells exhibited an inhibitory effect on DC maturation through the production of prostaglandin E2 (33). Some studies suggested that IL-10 might be one tool to modulate the immune response in HTLV-I-infected patients (9,22). In this study, we observed that culture supernatants from MT-2 and B95-a cells contained a large amount of IL-10, whereas those from HTLV-BrMMø contained little IL-10; however, culture supernatants from B95-a did not inhibit GM-CSF- and IL-4-mediated induction of CD1a expression, whereas those from HTLV-BrMMø showed significant inhibition, and furthermore, anti-IL-10 neutralizing antibody could not restore CD1a expression. These findings suggest that IL-10 is not responsible for the impairment of monocyte differentiation to DCs in our systems. Since culture supernatants from MT-2 and HTLV-DCs exhibited a similar inhibitory effect on monocyte differentiation to DCs, we speculated that HTLV-I infection might cause the production of some inhibitory cytokines in infected cells.
Recently Azakami et al. reported a decreased frequency of iNKT cells in PBMCs from HAM/TSP and ATL patients (5). We also observed that when PBMCs were cultured with α-GalCer in the presence of HTLV-BrMMø-sup, the percentage of iNKT cells was significantly decreased. iNKT cells are characterized by a semi-invariant T-cell receptor (TCR) using a unique TCR Vα24Jα18 chain in humans, and by its recognition of α-GalCer loaded onto CD1d molecules. They exert potent killing activity and rapidly produce cytokines and chemokines that influence many other cells of the innate and adaptive immune systems. Thus, iNKT cells have been shown to play a critical role in immune regulation, ranging from the development of autoimmunity to responses to pathogens and tumors (37); therefore, the low number of iNKT cells in HTLV-I-infected patients might result in the insufficient control of HTLV-I-infected cells, and lead to the development of ATL.
We found that treatment of monocytes with HTLV-BrMMø-sup exhibited potent inhibitory effects on both T-cell and iNKT-cell proliferation. An effective T-cell response reflects the presence of necessary co-stimulatory and proinflammatory cytokines, such as IL-12, whereas suppression of T-cell responses reflects the absence of co-stimulation and the presence of inhibitory cytokines such as IL-10 (10). When monocytes were cultured in CTM supplemented with GM-CSF and IL-4 in the presence of HTLV-BrMMø-sup, the expression level of CD1a, DC-SIGN, and HLA-DR was significantly downregulated. Moreover, HTLV-BrMMø-sup-treated monocytes exhibited decreased production of IL-12 after simulation with CD40L; therefore, the failure of T-cell proliferation in MLR might be due to the deficiency of both co-stimulatory and proinflammatory cytokines. However, since the expression level of CD86 was upregulated and that of CD40 was not altered, we have to examine the stimulatory capacity of monocytes for an antigen-specific T-cell response. In addition, although the expression level of CD1d was also slightly upregulated, induction of iNKT cells was suppressed. It is much more likely that antigen-processing and antigen-presenting functions may be damaged in monocytes during the differentiation process. Further studies are required to elucidate the characteristics of impaired monocytes and the relationship between the inhibitory effects of HTLV-I-infected cells on monocyte differentiation and dysfunction of HTLV-I-specific immune responses.
Treatment of iDCs with HTLV-BrMMø-sup resulted in few significant changes in the expression of surface molecules compared to untreated iDCs, except CD86. CD86 is a co-stimulatory molecule that engages with CD28 on T cells and provides T cells with co-stimulatory signals. We do not know why the expression level of CD86 was upregulated by treatment with HTLV-BrMMø-sup. Although the molecular mechanism underlying CD86 regulation remains unclear, a recent study demonstrated that CD86 is ubiquitinated in DCs via ubiquitin ligase, and that ubiquitination plays a key role in CD86 expression (8); therefore, treatment of monocytes with HTLV-BrMMø-sup might result in inadequate ubiquitination of CD86, which would lead to an increase in CD86 expression.
Hasegawa et al. have reported that orally HTLV-I-infected rats exhibited weaker HTLV-I-specific cellular immune responses with an increased viral load (14). When HTLV-I-infected BrMMø are ingested with maternal breast milk and arrive at the newborn intestinal tract, they might transmit HTLV-I virions to intestinal lymphocytes and monocytes. Moreover, HTLV-I-infected BrMMø might inhibit the proliferation and differentiation of immune cells, including local innate immune responses. These early disturbances in innate immunity of the newborn intestine may result in a failure to develop HTLV-I-specific immune responses.
In the present study, we showed that soluble products from HTLV-I-infected BrMMø exhibited potent inhibitory effects on both T-cell and iNKT-cell proliferation. This indicated functional impairment in both adaptive and innate immunity. Although we have not closely analyzed the soluble products present in culture supernatants from HTLV-I-infected BrMMø, identifying the factors responsible for the impairment of monocyte differentiation to DCs may be helpful for both understanding HTLV-I pathogenesis, and developing new therapeutic approaches.
Footnotes
Acknowledgments
We thank Dr. Watari, Dr. Shinya, and Ms. Owaki for technical support, and Dr. Akiba (Akiba Hospital) for providing breast milk samples. This work was supported in part by grants from the Ministry of Education, Science, Sport, and Culture, from the Ministry of Health and Labor and Welfare, Japan, from the Japanese Health Sciences Foundation, and by the Promotion and Mutual Aid Corporation for Private Schools of Japan.
Author Disclosure Statement
No conflicting financial interests exist.