
Abstract
Marek's disease (MD) is a lymphoproliferative disease of domestic chickens caused by a highly cell-associated alpha herpesvirus, Marek's disease virus (MDV). Clinical signs of MD include depression, crippling, weight loss, and transient paralysis (TP). TP is a disease of the central nervous system that affects MD-susceptible chickens 8–11 days post-infection (dpi), normally resulting in recovery 1–3 d after the onset of clinical signs. In this study we inoculated chickens from lines 72 (MD-susceptible) and 63 (MD-resistant) with a very virulent plus strain of MDV at 2 wk of age, and collected brain samples from birds with and without TP at 5, 11, and 21 dpi for gene expression profiling and histological analysis. Data revealed that chickens inoculated with MDV had higher levels of IL-6, IL-10, IL-18, IFN-α, IFN-β, IFN-γ, MHC I, and CD18 in their brains at 11 dpi compared to the uninfected control birds. In addition, the expression levels of IL-6, IL-10, IFN-α, IFN-β, and IFN-γ were significantly higher in the brains of the birds showing clinical signs of TP than in asymptomatic chickens. Comparative analysis between the two chicken lines showed that the expression levels of IL-6, IL-10, IFN-β, IFN-γ, IL-18, CD18, and MHC I were significantly higher in the brains of the birds from line 63 with TP than those of line 72 exhibiting neurological disorders. A differential expression pattern was observed for some of the tested genes at different time points post-inoculation. Histological analysis showed lymphocytic meningitis, perivascular cuffing, and neuronal degeneration within the brains of birds from both susceptible and resistant lines exhibiting TP at 11 dpi. Vaccination prevented development of TP and other MD-associated clinical symptoms. These observations are suggestive of an underlying immunological mechanism for viral-induced neurological dysfunction, and the differential responses of the two chicken lines to MDV infection.
Introduction
In addition to the induction of T-cell lymphomas, MDV infection results in immunosuppression, transient paralysis (TP), and eventual death (12,21,26,28). The classic MD-induced TP is generally followed by an abrupt and complete recovery of the afflicted birds within 2–3 d post-paralytic signs (14). A more severe form of neurological disorder, termed acute TP, is also exhibited in chickens infected with highly virulent MDV strains (33). Acute TP is characterized by an irreversible flaccid paralysis of the neck and limbs 9–11 dpi that generally leads to mortality 1–3 days after the onset of neurological signs (11,33).
In addition to the pathotype of the virus, genetic background of the host also plays a role in the severity of the neurological disorder (10). Development of TP has been linked to a recessive trait controlled by major histocompatibility complex (MHC) genes. The G-B1 line with a B13/B13 genotype was found to be resistant, whereas a G-B2 line with a B6/B6 MHC haplotype was susceptible to development of TP (6,23,29,30). These studies, however, use a virulent strain of MDV that is capable of inducing only the classical form of TP.
Histological studies have revealed that vasculitis, perivascular cuffing of mononuclear cells, vacuolation, and edema are common features observed in the brain lesions of TP-afflicted birds. Comparative analysis indicates that most of the brain lesions of birds with clinical signs of TP are similar to those of asymptomatic MDV-infected chickens (6,15,16,30). Severe and diffuse vasogenic edema, however, was a consistent finding only in the brain lesions of birds with clinical signs of TP (30). Despite the extensive investigation of pathogenic changes associated with MD-induced TP, its underlying molecular mechanism is still largely unknown.
Cytokines have been implicated in the neuropathogenesis of viral encephalitis. Detection of considerable levels of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), RANTES (CCL5), and interferon-γ-induced protein 10 (IP-10) has been reported in human non-permissive microglial cell infection with herpes simplex virus (19). It has been postulated that enhanced expression of the proinflammatory cytokines TNF-α and IFN-γ is associated with neuroinvasion of simian immunodeficiency virus (SIV) and the development of viral-induced encephalitis (22). A progressive decrease in the expression of IL-4 and IL-10 (anti-inflammatory cytokines) was found to be inversely correlated with an increase in the transcriptional activities of proinflammatory cytokines (TNF-α and IFN-γ), and histopathological changes with negative consequences in Japanese encephalitis virus infection in mouse brain (27). And finally, the expression of proinflammatory cytokines has been implicated in the pathogenesis of AIDS dementia complex (35).
Results from two relevant studies investigating the biological functions of a limited number of genes in association with TP indicate that cytokines and other immune-related genes might also play a crucial role in the induction of pathological lesions seen in the brain tissues of TP-afflicted birds. Data from Jarosinski et al. (13) showed a large increase in the expression of inducible nitric oxide synthase (iNOS), interferon-γ (IFN-γ), IL-1β, IL-6, and IL-8 in the brains of chickens infected with a very virulent plus (vv+) strain of MDV. This study also showed that in-vivo levels of nitric oxide (NO) were significantly higher in vv+MDV-infected chickens with neurological disorders than asymptomatic birds infected with the same virus. A study by Abdul-Careem et al. (1) also found an increase in the transcriptional activity of IL-6, IL-12, and IFN-γ in the brains of infected birds compared to uninfected control chickens. Additionally, the expression of IL-6, IL-18, and IFN-γ was shown to be significantly higher in the brain tissues of birds with TP than chickens showing no clinical signs of TP. The authors of both studies speculate that proinflammatory cytokines and other immune-related genes may play a major role in neurological disorders associated with MD.
The purpose of current study was to investigate the potential involvement of cytokines and chemokines by examining the roles of an extensive panel of immune-related genes in MDV-induced TP in susceptible and resistant chicken lines harboring the same MHC haplotype (B2/B2), that develop the acute form of TP after inoculation with a vv+strain of MDV.
Materials and Methods
Chickens
The specific-pathogen-free chickens used in this study were from two highly inbred lines of 63 (MD-resistant) and 72 (MD-susceptible) birds (2). These birds were from unvaccinated breeder hens and carried no maternal antibodies to MDV or herpesvirus of turkeys (HVT). The chicks were hatched at the Avian Disease and Oncology Laboratory poultry facility, and housed in modified Horsfall-Bauer isolation units for the duration of the experiment.
Virus and vaccine
A vv+strain of MDV, 648A-p8 (passage 8), which is propagated and maintained in our laboratory, was used in this experiment (32). The commercially-available vaccine CVI988/Rispens was used for the vaccination of the birds.
Experimental design
One-day-old chicks from each line were randomly distributed into 4 groups of 25 birds each in separate isolators (A, B, C, and D). Birds in group A were vaccinated with CVI988 at 1 wk of age per the manufacturer's recommended dose and challenged with 5000 PFU of 648A 1 wk post-vaccination. Birds in groups B and C were vaccinated and challenged, respectively, at 2 wk of age. Birds in group D served as negative controls. Group C, infected with 648A, was performed in duplicate (25 birds per isolator, 50 total). At 5 dpi, three birds from each of the four groups of each line were euthanized by CO2 inhalation and necropsied. At 11 dpi, six birds from the 648A-inoculated chickens (three with the acute form of TP, and three without any obvious neurological disorder), and three birds from each of the remaining groups of each line were also euthanized by CO2 inhalation and necropsied. At 21 dpi, six birds from 648A-inoculated chickens (three without any prior neurological disorder, and three from recovered TP-afflicted birds), and three birds from each of the remaining groups of each line were euthanized and necropsied. The total brain tissues were collected and immediately stored in RNAlater (Ambion, Austin, TX) to prevent RNA degradation. For histological studies at 11 dpi, six more infected birds (three with TP and three without TP), and three more uninfected control birds of each line, were also necropsied and portions of the brain tissues (the cerebellum and cerebrum) were collected.
Histopathology
The collected brain samples were immersed in 10% neutral buffered formalized saline solution. After dehydration in graded ethanol solutions, the samples were embedded in paraffin, sectioned, and mounted on glass slides for hematoxylin and eosin staining.
RNA isolation
Total RNA was isolated from the homogenized brain tissues of three birds of each of the above-mentioned groups at 5, 11, and 21 dpi (three biological replications) by the Qiagen (Valencia, CA) RNeasy mini kit according to the manufacturer's instructions. Residual DNA was digested by DNase treatment of RNA samples using Ambion's RNase-free DNase kit. The levels of gene expression in the brain tissues of control birds were used as reference or baseline for calculation of fold changes in gene expression in MDV- or Rispens-inoculated chickens.
Real-time RT-PCR
Real-time PCR analysis of the relative quantification of chicken gene expression transcripts was carried out at the Research Technology and Support Facility of Michigan State University in East Lansing, Michigan. Briefly, 2.5 μL of a 1:20 dilution of the oligo dT-based RT product from 4 μg of total RNA isolated from brain tissues was used for each reaction. Then 300 nM of each for specific sense and anti-sense primers were used in the presence of 5 μL SYBR Green PCR master mix (Applied Biosystems, Foster City, CA). The amplification program was as follows: 50°C for 2 min, 95°C for 10 min, 40 cycles at 95°C for 15 sec, followed by 60°C for 1 min. All the reactions were run in duplicate in a 7900HT Sequence Detection System (Applied Biosystems). The primers for chicken genes were designed using MacVector software (Accelrys, San Diego, CA). All the primers were synthesized by Operon Biotechnologies, Inc. (Huntsville, AL). The primer sequences and their amplicons are listed in Table 1.
Statistical analysis
The web-based software, Relative Expression Software Tool (REST), was used for relative quantification of chicken gene expression and significance of fold changes. The significance of the fold changes in gene expression is determined by a pair-wise fixed reallocation randomization test, and plotted using standard error (SE) estimation via a complex Taylor algorithm (
Results
Clinical observation
In the course of the experiment designed to test the response of two genetically-diverse chicken lines with the same MHC haplotype (B2/B2) to a vv+strain of MDV, a high mortality rate was observed between 8 and 15 dpi. More than 40% of MDV-infected birds in line 72 and close to 20% of birds in line 63 died from an acute form of TP by 15 dpi. The syndrome was characterized by an initial depression followed by a rapid progression of neck and limb paralysis, and eventual death within 24–48 h after the onset of the neurological disorder.
Histopathological observations
The MDV-infected chickens of both susceptible and resistant lines with clinical signs of TP had moderate to severe lymphocytic meningitis, perivascular cuffing by lymphocytes, gliosis, and neuronal degeneration in the cerebrum and cerebellum sections of their brains. The asymptomatic infected chickens of line 72 exhibited a mild form of lymphocytic meningitis, with perivascular cuffing by lymphocytes, gliosis, and neuronal degeneration. The MDV-infected birds from line 63 with no obvious clinical disorder, however, had only lymphocytic meningitis and perivascular cuffing within their brains. The uninfected control chickens had no histological lesions (Fig. 1). The vaccinated or vaccinated/challenged birds exhibited no MD-associated clinical signs and no histological analysis was performed.
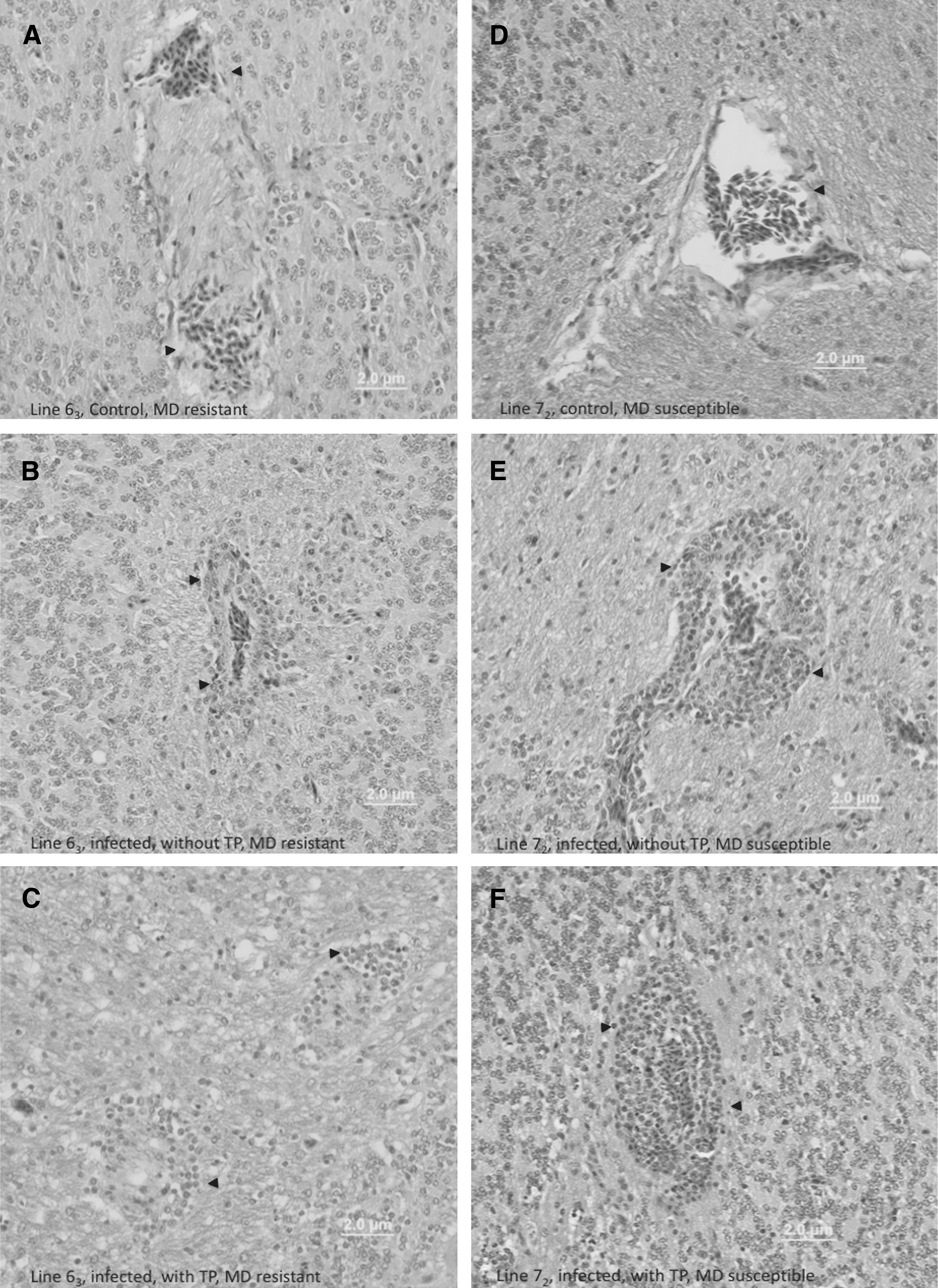
Histological brain lesion of 648A-inoculated and control chickens from line 63 (
Gene expression analysis
The relative quantification and fold changes in gene expression were based on REST software analysis. Changes were normalized to the expression levels of the housekeeping gene β-actin. The numerical values of fold changes were converted to bar graphs, and statistical significance was determined using the same software (Figs. 2, 3, and 4). Comparative fold changes in the expression levels of the tested genes between the susceptible and resistant lines indicated differential immune responses to MDV infection. The MDV-inoculated chickens of both lines, regardless of their clinical signs, had higher levels of IL-6, IL-10, IL-18, IFN-α, IFN-β, IFN-γ, MHC I, and CD18 in their brain tissues at 11 dpi compared to the uninfected control birds. In addition, the transcriptional activities of IL-6, IL-10, IL-18, CD18, IFN-α, IFN-β, and IFN-γ were significantly elevated in the brains of the birds with neurological disorders compared to asymptomatic MDV-infected birds. Furthermore, comparative analysis revealed that the expression levels of IL-6, IL-10, IL-18, CD18, IFN-β, IFN-γ, and MHC I, were significantly higher in the brain tissues of birds from line 63 with TP than those of line 72 exhibiting clinical signs of TP (Fig. 3). An indirect correlation between the expression patterns of IL-10 and MHC II was evident in the chickens of line 72. High expression levels of CD18, the β subunit of the β2 integrin family of adhesion molecules, in both susceptible and resistant lines was suggestive of leukocyte activation and transmigration into the brains of MDV-infected birds. Finally, the expression of Bcl2, an anti-apoptotic gene, was downregulated in all treated groups of line 72, and in MDV-infected groups of line 63. This suppression was more severe in TP-afflicted birds of line 72 than all the other treated groups. In the vaccinated/challenged group of line 63, however, expression of Bcl2 was increased. The transcriptional activity of Bcl2 was not detected in the vaccinated group of line 63.
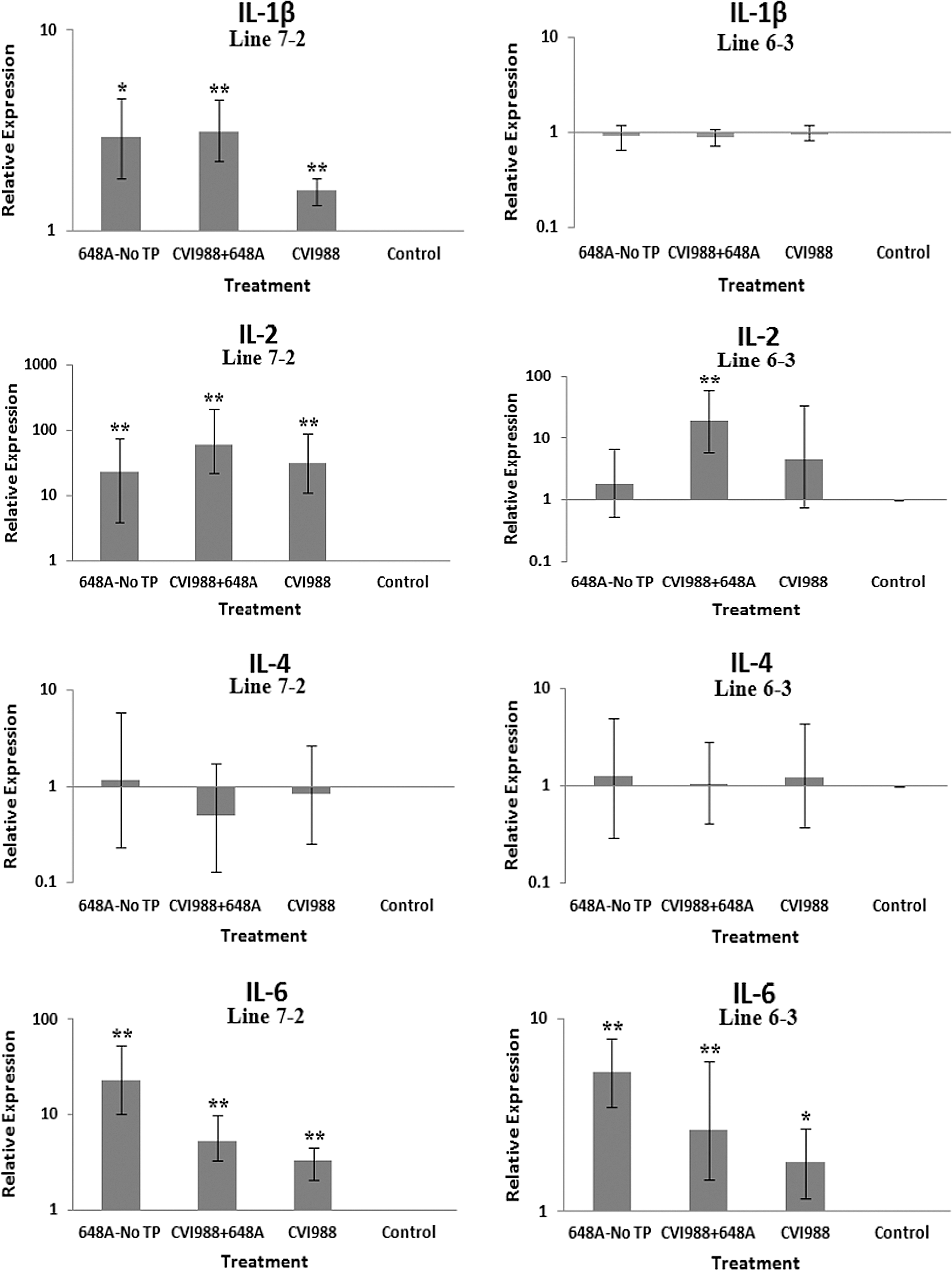
Bar graphs depicting fold changes in cytokines, chemokines, and other immune-related gene expression patterns at 5 dpi. The bar graphs on the left side of the panels depict gene expression patterns in the brain tissues of birds from line 72. The graphs on the right side of the panels show the expression levels of the corresponding genes in the brain tissues of birds from line 63. The relative quantification and fold changes in gene expression were determined by using the expression levels in the control birds as reference or baseline. The fold changes were normalized to the expression levels of the housekeeping gene b-actin. The numerical values of fold changes were converted to bar graphs, and statistical significance was determined using REST (*p < 0.05; **p < 0.01; 648A, virus; CIV1988, vaccine; TP, transient paralysis).
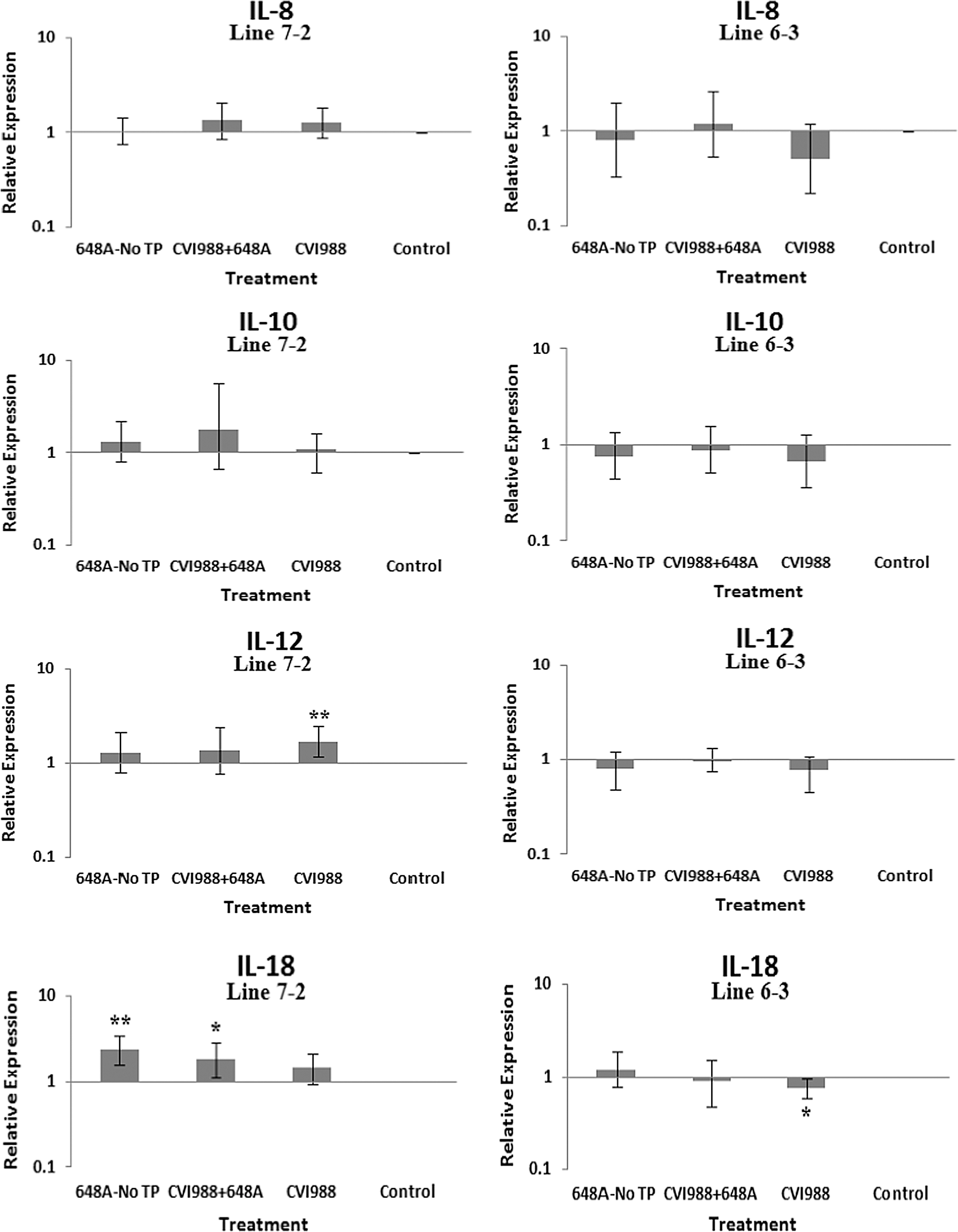
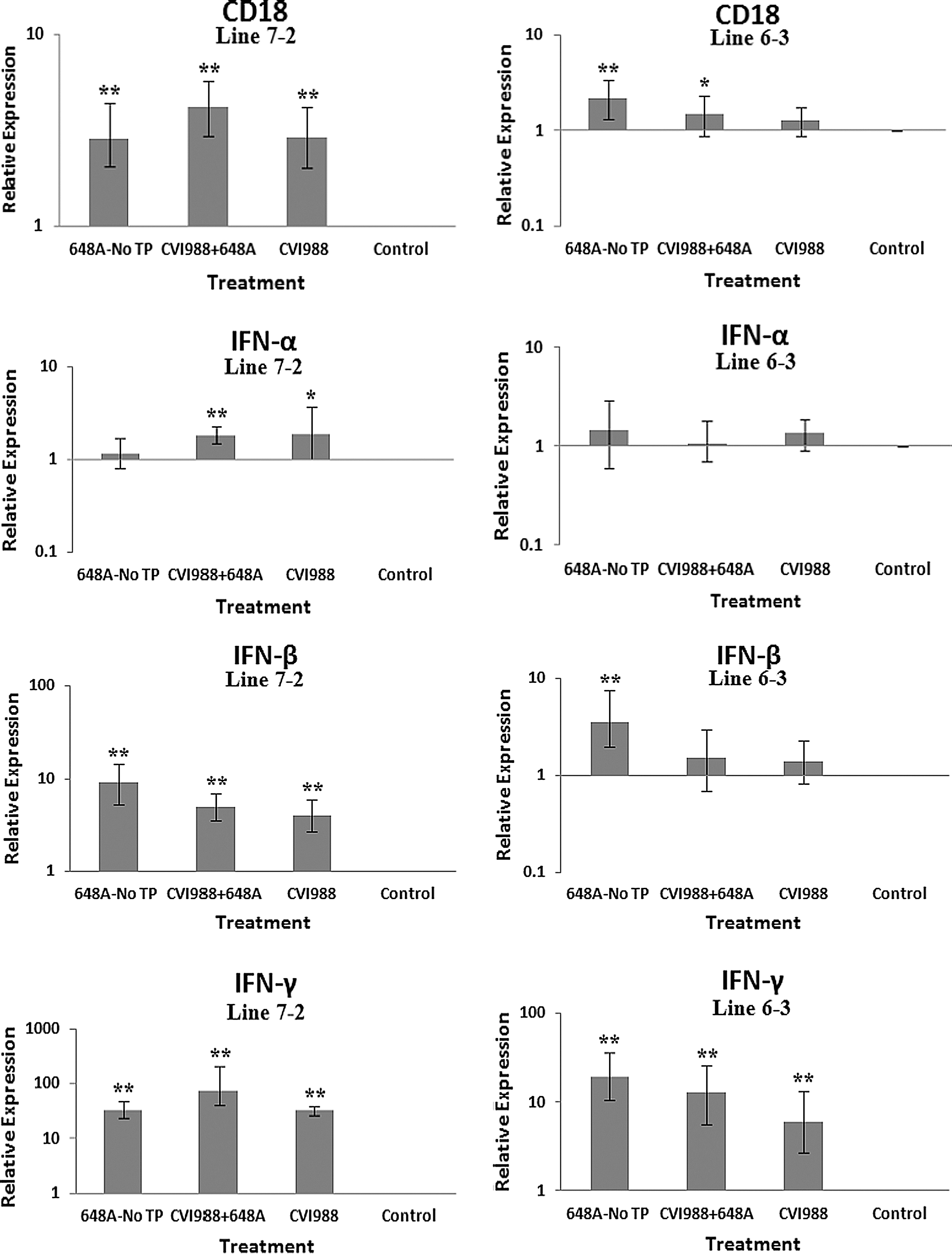

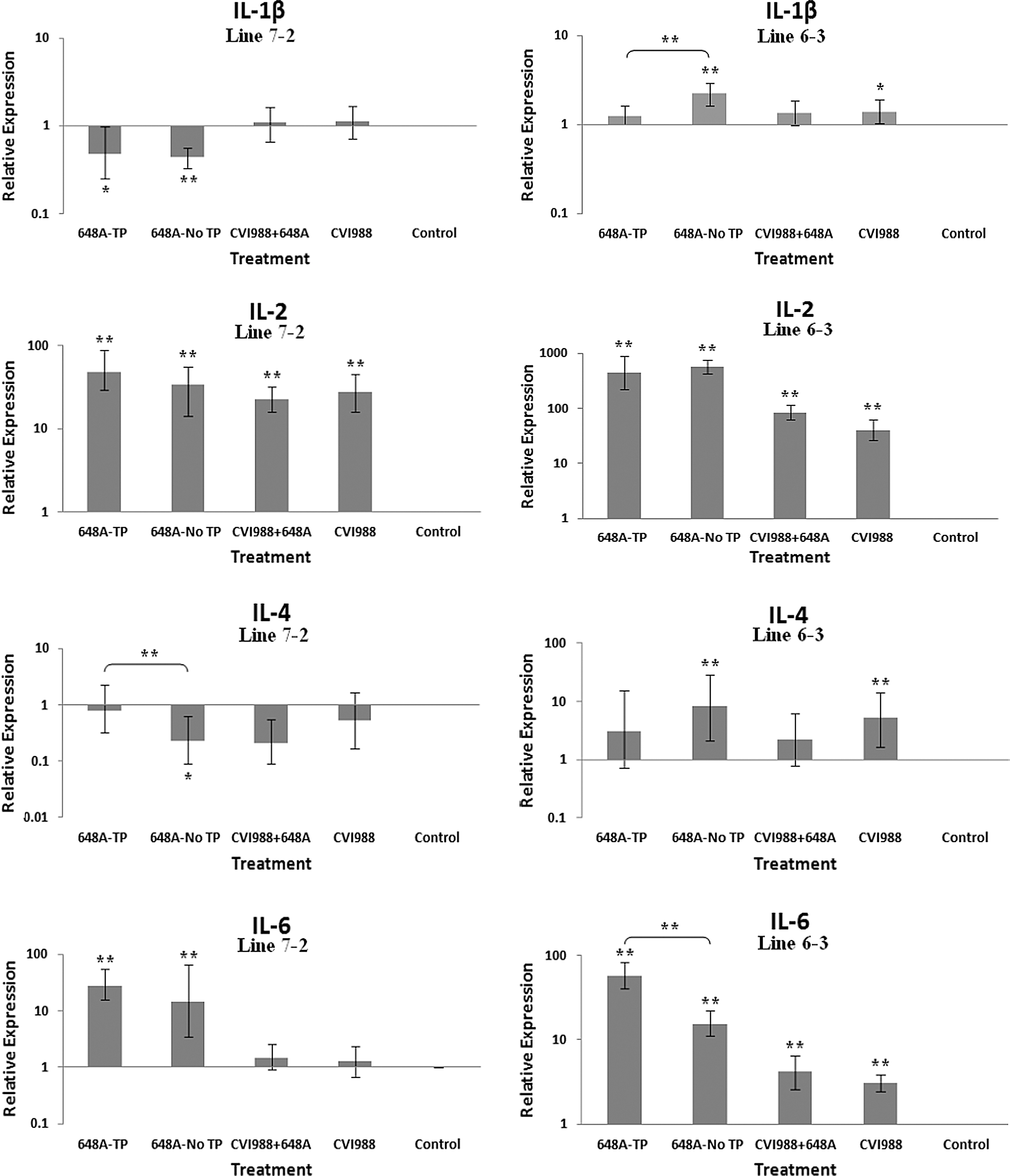
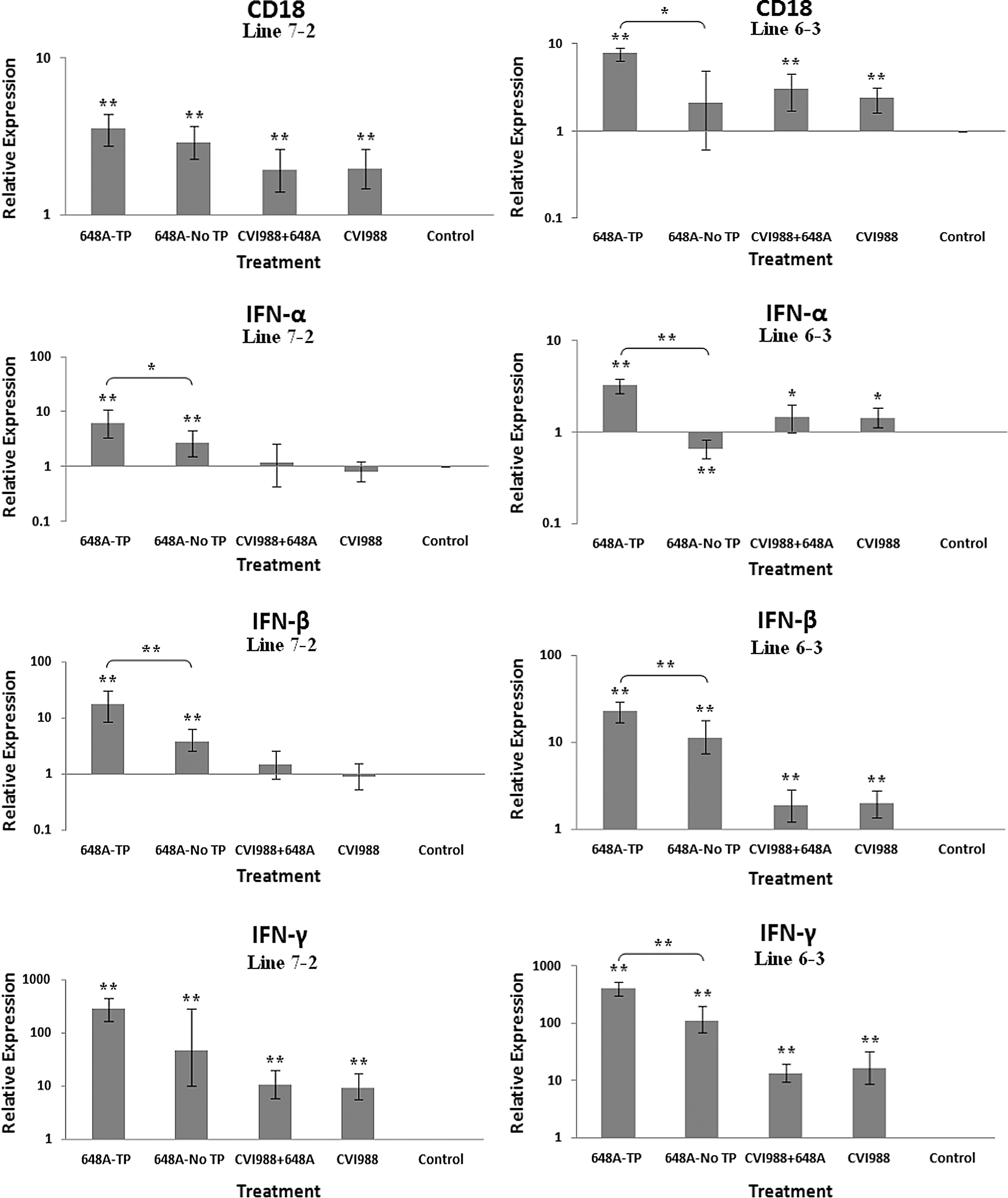
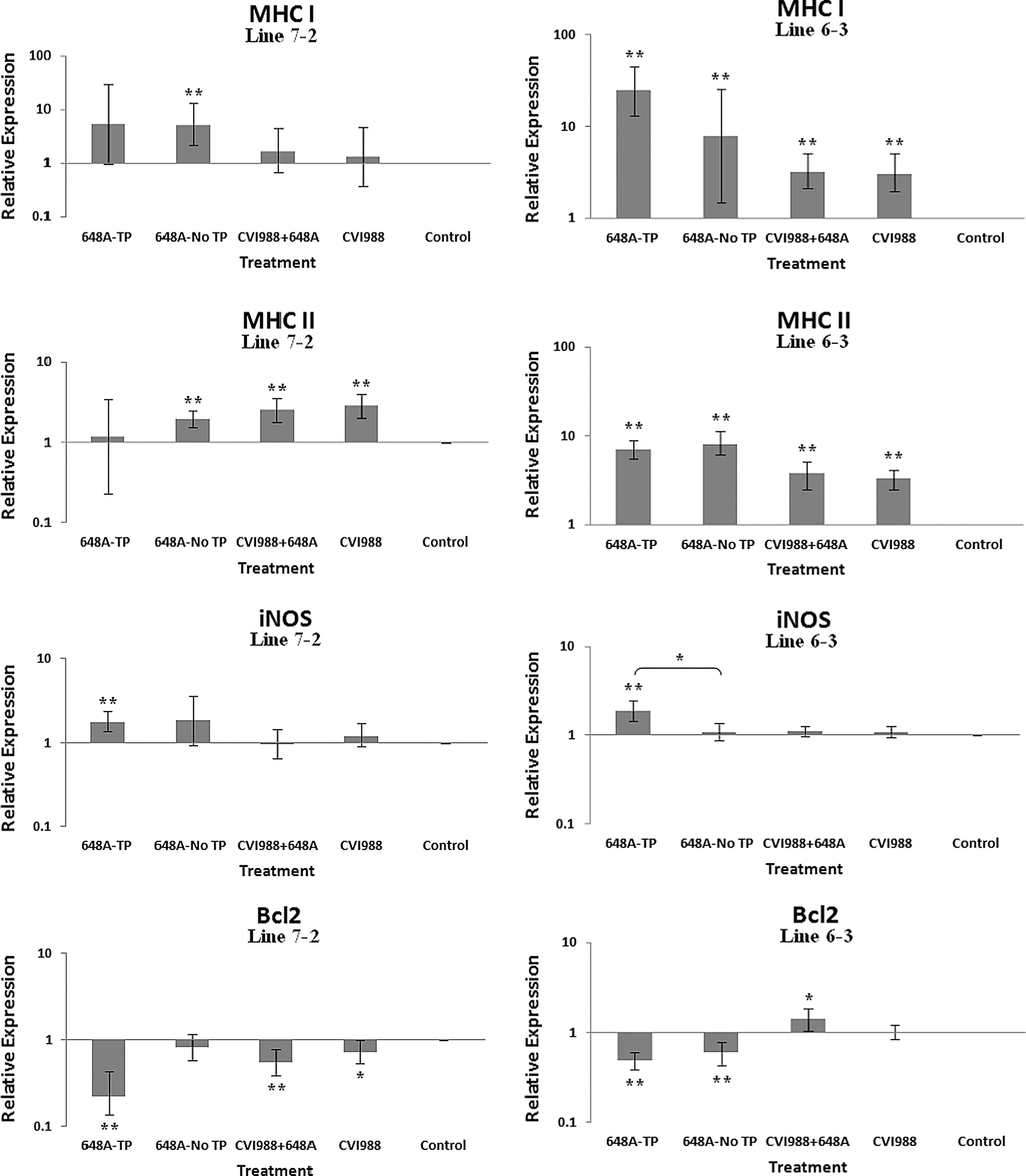
Bar graphs depicting fold changes in cytokines, chemokines, and other immune-related gene expression patterns at 11 dpi. The relative quantification and fold changes in gene expression were determined by using the expression levels in the control birds as reference or baseline. More than 40% of MD-susceptible and close to 20% of the resistant birds developed an acute form of TP between 9 and 11 dpi. The numerical values of fold changes were converted to bar graphs and statistical significance was determined using REST software (*p < 0.05; **p < 0.01; 648A, virus; CIV1988, vaccine; TP, transient paralysis).

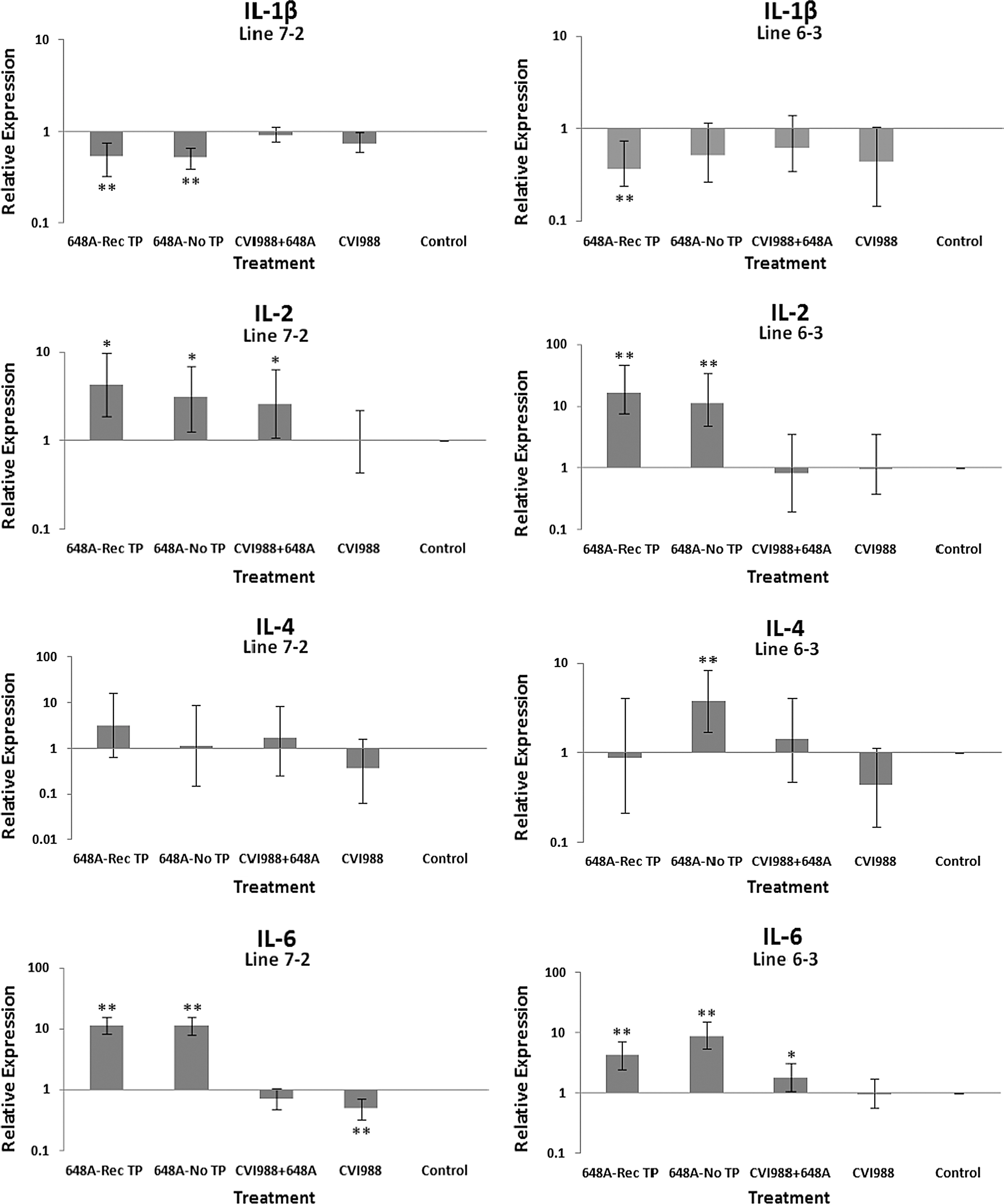
Bar graphs depicting fold changes in cytokines, chemokines, and other immune-related gene expression patterns at 21 dpi. The relative quantification and fold changes in gene expression were determined by using the expression levels in the control birds as reference or baseline. The numerical values of fold changes were converted to bar graphs and statistical significance was determined using REST software (*p < 0.05; **p < 0.01; 648A, virus; CIV1988, vaccine; TP, transient paralysis; Rec TP, TP-afflicted birds that recovered from MDV-induced neurological disorders between 9 and 11 dpi).
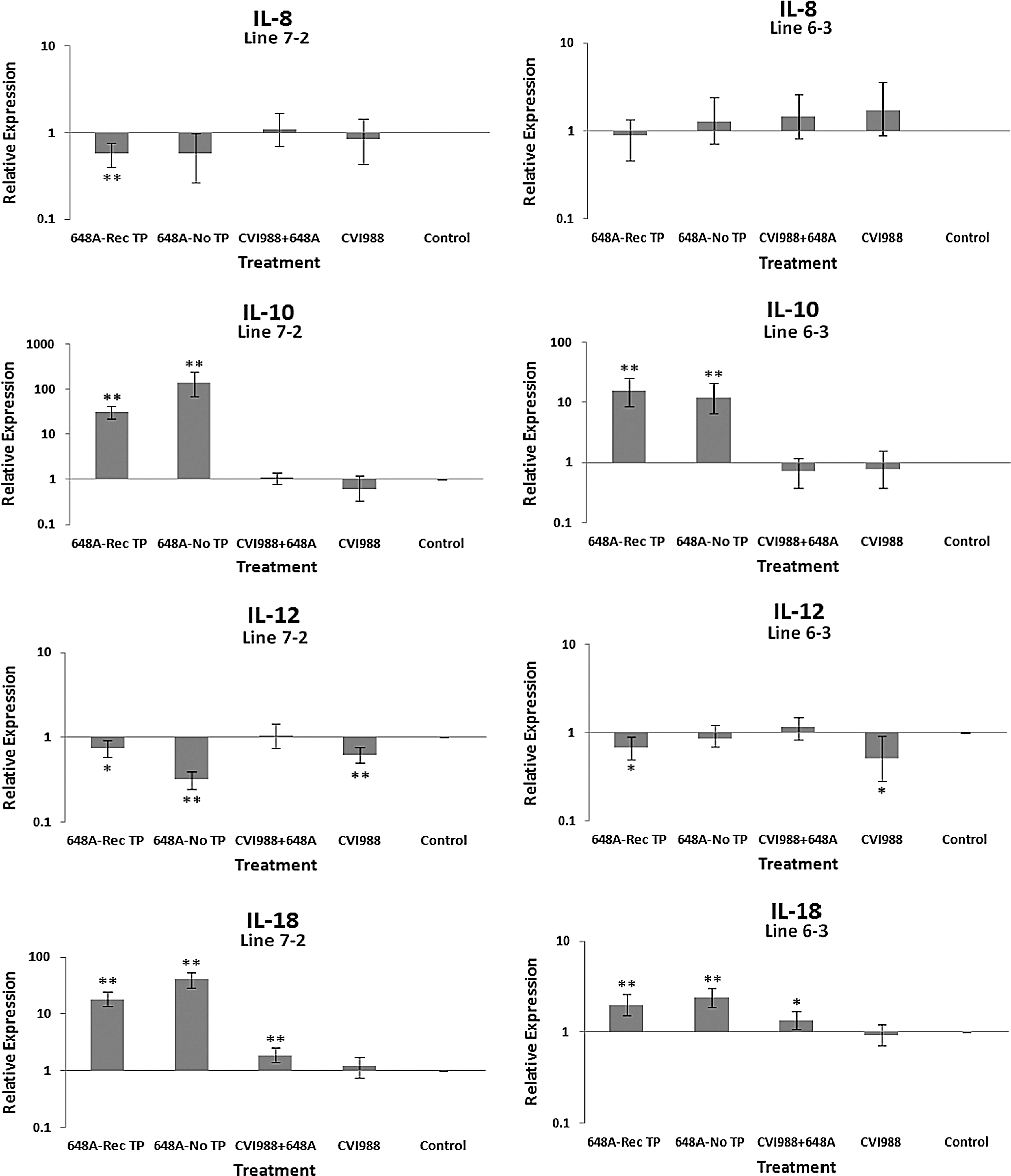
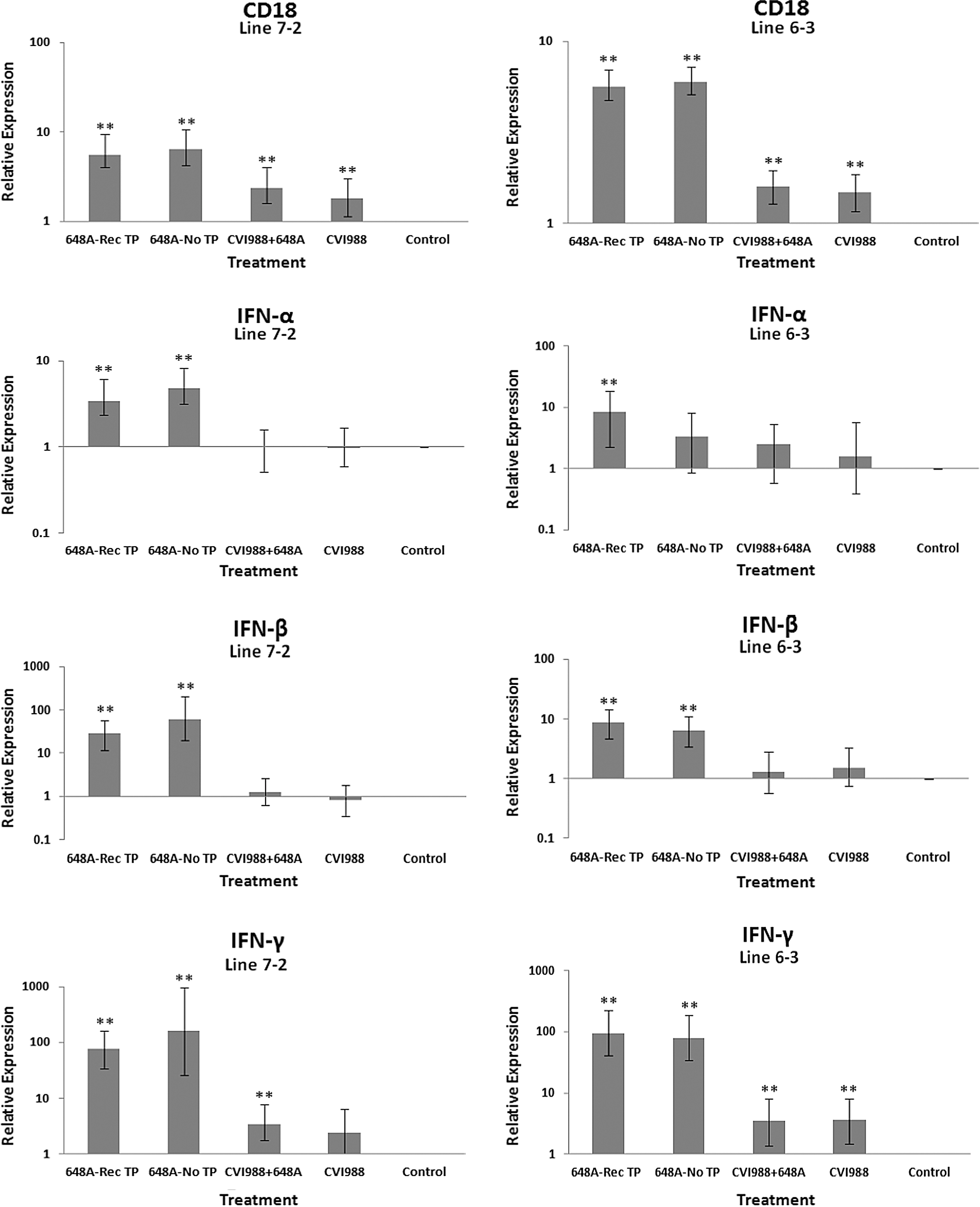
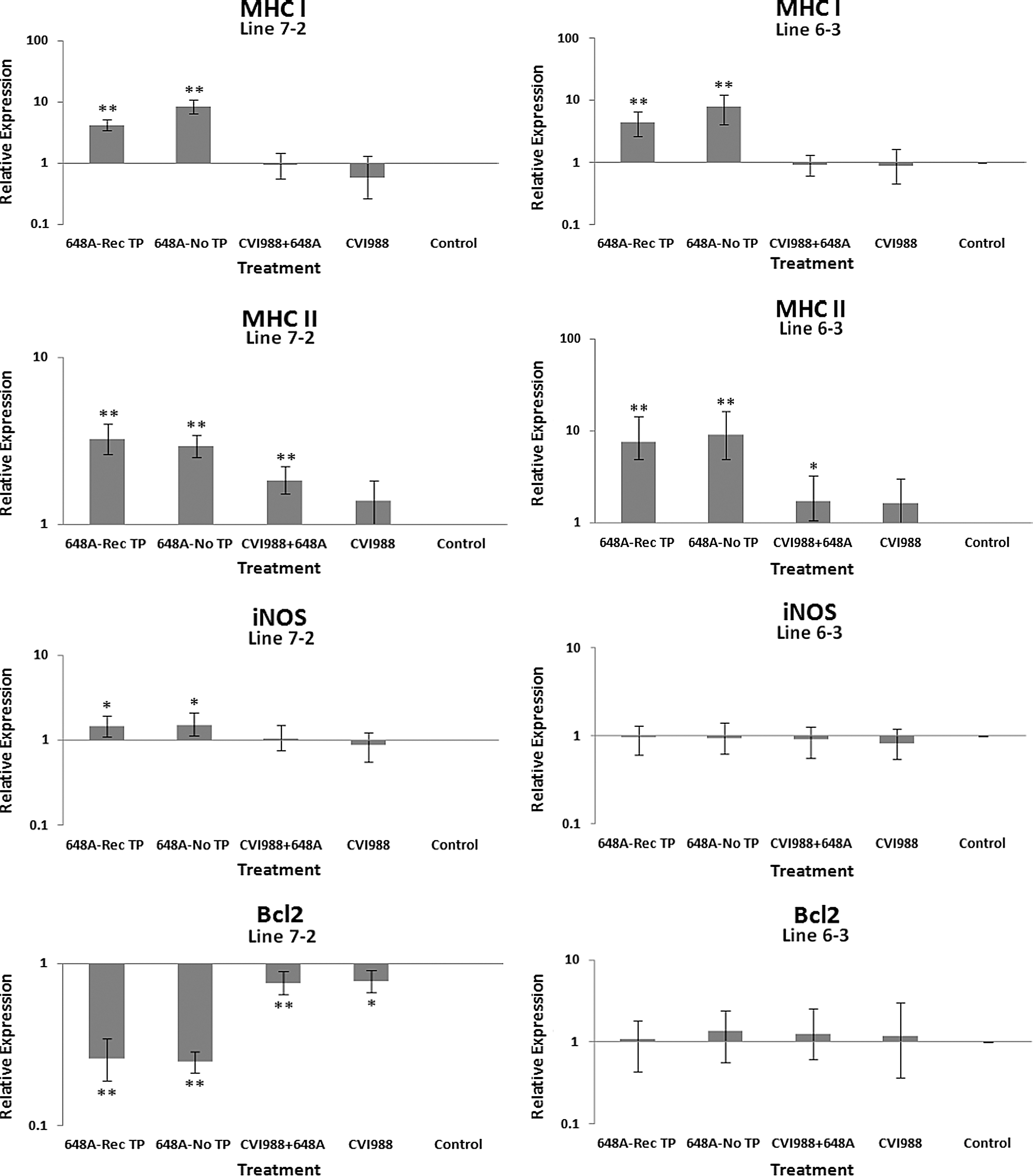
Comparative analysis of gene expression at three time points (5, 11, and 21 dpi) revealed changes in expression patterns during different phases of MDV infection. IL-1β was upregulated in the brains of the infected birds of line 72 at 5 dpi (cytolytic infection), but was downregulated at 11 and 21 dpi (latent infection and reactivation, respectively). This proinflammatory cytokine showed no transcriptional activities at 5 dpi, minor upregulation at 11 dpi, and downregulation at 21 dpi in the brains of the treated groups of line 63. IL-2 was highly upregulated at 11 dpi in both chicken lines, with reduced activities seen at 5 and 21 dpi. IL-4 showed a differential pattern of expression in both lines at all three time points. IL-6, another proinflammatory cytokine, was abundantly expressed in both chicken lines during cytolytic infection, followed by a similar pattern of expression at 11 and 21 dpi. It is clear, however, that the expression level was higher in the infected birds of line 63 than those of line 72. IL-8 was minimally active in line 63, with minor suppression in the infected birds of line 72 at 11 dpi. Minor changes were observed in the expression of this heterophil chemoattractant in both lines at 5 and 21 dpi. The expression of IL-10 and its inverse correlation with the expression pattern of MHC II at 11 dpi was mentioned above. Its transcriptional activity was extended to 21 dpi in 648A-infected chickens of both lines. IL-12, a cytokine associated with a Th-1 type of immune response, showed minor changes in expression during three phases of MDV infection. IL-18, a potent inducer of IFN-γ, was actively expressed in both lines at 11 dpi, with some extending to 21 dpi. At 5 dpi, however, it was only upregulated in inoculated birds of line 72. CD18, a β subunit of β2 integrin adhesion molecules, showed transcriptional activities during all three phases of infection, with the highest expression levels in line 72 at 5 dpi, and line 63 at 21 dpi. IFN-α was minimally activated, while IFN-β and IFN-γ were both highly expressed during the three phases of MDV infection, with variations seen among the inoculated birds of either line. MHC I expression was the highest at 11 dpi in line 63 among all the inoculated chickens of either line. MHC II, on the other hand, showed high levels of activity during the cytolytic, latent, and reactivation phases of infection in all inoculated birds, with the exception of birds in line 72 at 11 dpi. The pattern of expression in these latter birds was inversely correlated with that of IL-10 at the same time point. There was little transcriptional activity for iNOS in the inoculated chickens of either line throughout the study. Finally, the expression of Bcl2 was clearly suppressed in most inoculated chickens of line 72, indicating apoptotic-induced cell death in the brain tissues of 648A-infected birds.
Discussion
MDV is a highly cell-associated α-herpesvirus that causes various clinical syndromes, including T-cell lymphomas and neurological dysfunction. TP-susceptible chickens show progressive development of depression, ataxia, and recumbency. TP-afflicted birds usually recover within 24–48 h after the onset of clinical signs (4,24). An acute form of MDV, however, is induced by vv+strains of MDV, and is characterized by an irreversible generalized paralysis, leading to mortality shortly after the onset of neurological disorders (11,33). Earlier studies also suggest that TP is a recessive trait controlled by genes within or closely linked to the major histocompatibility complex (23,33).
To better understand the underlying mechanisms of MDV-induced TP, we analyzed the expression profiling of a panel of cytokines and other immune-related genes in the brain tissues of MDV-infected susceptible and resistant chicken lines with and without neurological disorders. In addition, histological analysis of the brain samples revealed the extent and severity of TP-associated lesions. The lesions observed in both the cerebellum and cerebrum consisted of lymphocytic meningitis, lymphocytic perivascular cuffing, neuronal degeneration, and gliosis. Interestingly, the lesions in the brains of birds showing clinical signs of TP were similar to those of infected birds exhibiting no clinical signs. As previously shown, the lesions in the brains of the symptomatic birds were severe, more diffuse, and generally associated with edema (31). Furthermore, TP-associated lesions of the MD-susceptible birds were more severe and extensive than those of the resistant line. No TP was observed in the vaccinated or vaccinated/challenged birds, and consequently no histological analyses were performed.
Transient infiltration of lymphocytes and macrophages into the TP-associated brain lesions is likely contributing to the clinical signs seen in the afflicted chickens. After the initial entry of MDV by some yet unknown mechanism and viral replication, it is likely that a specific viral transcriptional product (e.g., vIL8) acts as a chemoattractant that activates the endothelial cells, leading to vascular permeability and eventual attachment and trafficking of T cells and macrophages into the brain. A more likely scenario, however, is that microglial cells, the resident macrophages of the CNS, respond to a non-permissive MDV infection (3) by expressing considerable amounts of cytokines and chemokines, that leads to the activation of endothelial cells and upregulation of adhesion molecules, and consequently attachment and transmigration of leukocytes into the brain. These cells are mainly terminally differentiated T cells, macrophages, and plasma cells, that will eventually enter the cell death program in their enclosed microenvironment after exerting their effector functions (9).
Whether the cytokines or chemokines detected in the brains of MDV-infected chickens are the cause or effect of cell migration and the consequent induction of lesions and neurological dysfunction is not clear and needs further investigation. Silencing or blocking the biological functions of these soluble proteins could provide insight into the underlying mechanism behind TP.
In this report, we analyzed the expression patterns of 16 immune-related genes, including proinflammatory and anti-inflammatory cytokines, iNOS, MHC I, MHC II, and Bcl2, in the brains of MDV-infected susceptible and resistant chickens.
IL-1β, a proinflammatory cytokine, was somewhat downregulated in the infected chickens of line 72 with or without clinical signs. Minimal expression of IL-1β was detected in the vaccinated or vaccinated/challenged birds of the same line. In line 63, however, the infected birds exhibiting no clinical signs had higher expression levels of IL-1β than symptomatic vaccinated or vaccinated/challenged birds. Higher expression levels of this proinflammatory cytokine in the brains of the asymptomatic infected birds of line 63 is an indication of a more robust immune response to MDV infection than that seen in line 72. Inoculated birds of line 72 at 5 dpi had higher levels of IL-1β than those of line 63. The expression of this proinflammatory cytokine was suppressed at 21 dpi in the inoculated birds of both lines (Figs. 2 –4). No constitutive expression of IL-1β was detected in the control groups of either line at any time point post-inoculation.
IL-2, a Th-1-type cytokine, acts as a mediator of growth and functional activity of B and T cells (36). The transcriptional activity of this peliotropic cytokine followed a similar pattern in the corresponding groups of the treated birds of susceptible and resistant lines at 11 and 21 dpi. Comparative analysis between similarly treated groups of both lines showed higher expression levels in line 63 than in line 72. Once again, this was suggestive of a stronger immune response in the resistant line 63 than the susceptible line 72. At 5 dpi, however, higher activation of IL-2 is observed in the inoculated birds of line 72 than those of line 63.
IL-4, an anti-inflammatory cytokine, was downregulated in all four groups of treated chickens of line 72 At 11 dpi. In contrast, its expression was upregulated in all treated groups of line 63. Interestingly, the level of expression was lower in the symptomatic infected and vaccinated/challenged birds than in the other two groups. Suppression of this anti-inflammatory cytokine is generally inversely correlated to the upregulation of the proinflammatory cytokines TNF-α and IFN-γ, which leads to a Th-1-type of immune response. Minor transcriptional activities of IL-4 were observed at 5 and 21 dpi in either line.
IL-8, a strong chemoattractant and activator of neutrophils (17), is another cytokine that was suppressed in treated groups of line 72 compared to the corresponding groups of line 63 at 11 and 21 dpi. Downregulation of this essential chemokine was likely a viral-induced mechanism to suppress the effector function of macrophages that play critical roles to control MDV infection.
The expression patterns of the proinflammatory cytokines IL-6 and IL-18 were similar in the corresponding treated groups of both lines, with exception of line 63 at 5 dpi. The transcriptional activities of both genes, however, were higher in line 63 than line 72 at 11 dpi. In the study by Jarosinski et al. (13), however, the expression levels of IL-6 and IL-18 were higher in MD-susceptible line P2a than the resistant line N2a infected with a vv+MDV strain. The expression patterns of the proinflammatory cytokines IL-6 and IL-18, however, were in good agreement with those observed by Abdul-Careem et al. (1).
IL-12 is associated with a Th-1-type of immune response, and was upregulated in the symptomatic or asymptomatic infected groups of line 72 at 11 dpi. Not much activity was detected in the vaccinated/challenged group of the same line. It was rather unexpected to see suppressed expression of IL-12 in TP-afflicted chickens of line 63 compared to the corresponding group in line 72. Line 63 is relatively resistant to MD, and generally mounts a more vigorous immune response to MDV than line 72. IL-12 expression was suppressed in the inoculated chickens of both lines at 21 dpi.
In contrast to a previous study by Jarosinski et al. (13), there was minimal activity of iNOS in the brains of the treated groups of either line at any time point. This could be due to the highly pathogenic strain of MDV used in the study, which may have suppressed the expression of iNOS.
IL-10 is a cytokine associated with a Th-2-type immune response, and was significantly upregulated in the brains of symptomatic birds of both lines. It was also expressed at relatively high levels in the brain tissues of asymptomatic infected, and to some extent vaccinated and vaccinated/challenged birds, of line 72 at 11 dpi. These corresponding two latter groups in line 63, however, showed minor to no transcriptional activities of IL-10. Higher transcriptional activities of IL-10 were also observed in 648A-infected birds of both lines at 21 dpi. IL-10 is an anti-inflammatory cytokine that is known to suppress T-cell responses via a potent biological action on antigen-presenting cells, including dendritic cells and macrophages (8,25). IL-10 also serves as a potent mechanism for the downregulation of MHC II and co-stimulatory molecules (20). It is no surprise that the IL-10 expression level at 11 dpi is inversely correlated with the expression of MHC II in the MD-susceptible line 72. MHC II expression was also upregulated in the treated groups of both lines at 5 and 21 dpi. MHC I, on the other hand, was progressively upregulated from vaccinated to symptomatic groups in both lines at 11 dpi. Once again, like most other genes tested, higher expression of MHC I was exhibited in line 63 than in line 72. MHC I plays a critical role in adaptive immune responses by displaying peptides derived from intracellular pathogens to cytotoxic T cells (CTLs). CTLs specific for the MHC-peptide complex will recognize and destroy the infected cells in the process. At 5 dpi, a higher expression of MHC I was observed in the 648A-infected birds of line 63 than in those of line 72. At 21 dpi, however, its expression was suppressed in the recovered TP-afflicted birds in comparison to asymptomatic 648A-infected birds in either line.
The expression of IFN-α and IFN-β (type 1 IFNs), and the proinflammatory IFN-γ (type 2 IFN), followed similar patterns in both lines at all time points. CD18 is a β subunit of β2 integrin family of adhesion molecules that in its heterodimeric form (CD11/CD18) plays an essential role in inflammation by regulating leukocyte adhesion, transmigration, and function (18). Higher transcriptional activity of CD18 in all treated groups of both chicken lines, specifically the TP-afflicted groups, was an indication of activation and transmigration of leukocytes into the brain. A higher expression level of CD18 was also observed in the treated groups of line 72 at 5 dpi, and the treated groups of both lines at 21 dpi. A significant downregulation of the anti-apoptotic gene Bcl2 in the brains of infected birds of both lines with or without clinical signs was a clear indication of cell death in the brains of afflicted chickens. Depression of the expression of Bcl2 was more severe in symptomatic birds of line 72 than in the other groups of either line. Downregulation of Bcl2 in the brain tissues of 648A-infected birds of line 72 was also evident at 21 dpi.
In conclusion, the data presented here show that despite the similarities in the response to MDV challenge in the CNS of MD-susceptible and resistant birds, there are clear differences between the two lines. Histological analysis revealed that pathogenic changes in the brains of MD-susceptible birds are more severe, with extensive perivascular cuffing, in addition to gliosis and neuronal degeneration. Gene expression analysis provided further insights into the possible underlying mechanism of TP, and the differential immune responses to MDV challenge of the CNS in these two chicken lines.
Although the proinflammatory cytokines were significantly upregulated in the brains of both chicken lines, the expression levels of these genes were generally higher in the treated groups of the resistant line than in those of the susceptible line. It is interesting to see a downregulation of IL-4 in all treated groups of line 72. We speculate that the decrease in the expression level of this anti-inflammatory cytokine has negative consequences for the pathological changes associated with MDV-induced TP. Finally, higher transcriptional activities of the anti-inflammatory cytokine IL-10 in all treated groups of line 72, and its inverse correlation with the expression of MHC II, is suggestive of a possible immunological mechanism behind the highly susceptible nature of line 72 to TP specifically, or MDV infection in general.
Footnotes
Acknowledgments
The authors would like to thank Barbara Riegle, Christine Harman, and Barry Coulson, for their excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.