
Abstract
Incorporation of molecular adjuvants into DNA vaccines is often used to improve the induction of immune responses, but few approaches aim to specifically activate B cells for an enhanced humoral response only. Hemokinin-1 (HK-1) is a factor that activates B cells for proliferation, survival, differentiation into plasma cells, and Ab production. Therefore, we investigated if it may be used as a molecular adjuvant for DNA vaccines to elicit strong humoral and memory responses. The HK-1 coding sequence was sub-cloned as single or triple copies in-frame downstream of S2 HBsAg in the proVAX/S2 construct. Compared to mice immunized with proVAX/S2 or proVAX/S2-HK-1, proVAX/S2-3HK-1 induced a higher level of IgG production, a higher percentage of differentiated antibody-secreting plasma cells, and a higher level of T-cell proliferation. Furthermore, a higher proportion of B cells had the B220+CD27+ phenotype in these groups, and specific antigen re-challenge induced a higher level of total IgG production 60 d after the last immunization, suggesting that the use of HK-1 as an adjuvant promoted immunological memory. Taken together, these results suggest that using HK-1 as an adjuvant molecule could enhance the immunogenicity of HBsAg DNA vaccines, and result in stronger humoral and memory responses. Therefore, HK-1 may lead to the development of a novel humoral-biased molecular adjuvant for an HBsAg DNA vaccine against hepatitis B infection.
Introduction
Compared with recombinant protein vaccines, DNA vaccines can be easily prepared on a large scale with high purity and stability (3). Moreover, they usually stimulate more potent cellular immune responses than protein antigens do (4). In fact, DNA vaccination has been suggested to be a good candidate technology for stimulating responses in non-responders to recombinant HBsAg vaccines, or for therapeutic vaccination for HBV carriers (5). However, DNA vaccines are often found to be relatively inefficient in the induction of antibody responses, and various approaches have been developed to improve them, particularly with respect to the choice of adjuvants (6).
Tachykinin defines a large group of 10 to 20 amino acid (aa) neuropeptides characterized by the presence of a common C-terminal motif, Phe-X-Gly-Leu-Met-NH2 (7). Hemokinin-1 (HK-1) is the latest addition to the tachykinin family, and is predominantly expressed in non-neuronal tissues (8). A recent study has demonstrated the potent impact of HK-1 on the activation of B cells, including their proliferation, survival, differentiation into plasma cells, and antibody (Ab) production (9). These results suggest that HK-1 might be an effective adjuvant for DNA vaccine responses, particularly when stronger humoral responses are needed.
In this work, we cloned the HK-1 coding region in-frame with the S2 antigen of an HBsAg DNA vaccine to assess its adjuvant effectiveness. Our results show that HK-1 is able to enhance the humoral immune response to HBsAg DNA vaccine by modifying the differentiation of plasma cells and promoting the generation of immune memory.
Materials and Methods
Animals, cells, and peptide
Female BALB/c mice aged 6–8 wk were purchased from the Animal Institute of the Chinese Medical Academy (Beijing, China), maintained under a 12-h light:dark cycle, and fed with pathogen-free food and water. An NIH/3T3 (ATCC CRL-1658™) cell line was maintained in Dulbecco's modified Eagle's medium (DMEM; GibcoBRL, Grand Island, NY), supplemented with 10% fetal calf serum (FCS), and penicillin/streptomycin (GibcoBRL), in a humidified incubator set at 37°C and 5% CO2. The HBsAg-derived peptide S208-215 (ILSPFLPL; H-2Kb-restricted) (10) was synthesized by GL Biochem Co., Ltd. (Shanghai, China).
Plasmid construction
The coding sequence containing the preS2 and S regions of HBV was cloned into the proVAX vector, constructed in this laboratory previously with a cytomegalovirus (CMV) promoter and a β-hCG leader sequence (11), and the new plasmid was designated as proVAX/S2. One or a series of three copies of the HK-1 coding sequence, each separated by five glycine residues, was inserted downstream of proVAX/S2 in-frame with the S2 antigen, resulting in the generation of proVAX/S2-HK-1 and proVAX/S2-3HK-1, respectively.
Transfection of the NIH/3T3 cell lines
The EGFP coding region was inserted downstream into proVAX/S2, proVAX/S2-HK-1, or proVAX/S2-3HK-1, in-frame with the S2, S2-HK-1, or S2-3HK-1 antigen, resulting in the generation of proVAX/S2-EGFP, proVAX/S2-HK-1-EGFP, and proVAX/S2-3HK-1-EGFP, respectively. The purified plasmids were transfected into NIH/3T3 cells with lipofectamine 2000 (Invitrogen, Carlsbad, CA), according to the manufacturer's instructions, and their expressions of EGFP were detected by fluorescence microscopy 48 h after transfection.
Immunization
The plasmids were prepared using an alkaline method and purified by PEG8000. The endotoxin was removed using the EndoFree Plasmid Giga kit (Qiagen, Hilden, Germany), following the manufacturer's recommendations. For vaccination, the mice were randomly divided into six groups (n=15 per group), as listed in Table 1, and immunized intramuscularly on days 0, 14, and 28, with either 100 μg plasmid in 100 μL saline solution, or the vehicle only.
The mice were randomly divided into 6 groups, 15 animals per group. The mice were immunized intramuscularly with either 100 μg plasmid in 100 μL saline solution or 100 μL saline solution only, on days 0, 14, and 28.
Detection of anti-HBsAg-specific antibodies and antigen-antibody affinity
The mice were bled on day 7 after the third immunization. The International Units of total IgG were measured with an enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer's instructions (SIIC Kinghaw Biotech Co. Ltd., Beijing, China). The 96-well microtiter plates in the kit were coated with 2 μg/mL rHBsAg antigen. The amount of total anti-HBsAg antibody was calculated according to the standard curve generated using an anti-HBsAg standard monoclonal Ab from the kit, and the mean number of units was determined from at least three points on the linear portion in the titration curve.
The mixture of HBsAg and the immunized serum was incubated at 37°C for 1 h, and then at 4°C overnight. The supernatants were harvested by centrifugation at 10,000 g for 30 min at 4°C. The antigen-antibody affinity was measured with a diagnostic kit for hepatitis B surface antigen according to the manufacturer's instructions (Kehua Bio-Engineering Co. Ltd., Shanghai, China).
T-cell proliferation assay
Mice immunized with DNA vaccine were sacrificed on day 7 after the third immunization, and lymphocytes in the spleen were prepared. Single-lymphocyte suspensions were incubated in triplicate in 96-well plates at 5×104 cells/well, in RPMI-1640 plus 5% FCS at 37°C in a 5% CO2 incubator for 48 h. They were stimulated during that time by the presence of 2 μg/ml HBsAg as specific antigen; 2 μg/mL bovine serum albumin (BSA) as irrelevant antigen; 0.1 μg/mL of phorbol myristate acetate (PMA) plus 1 μg/mL ionomycin (Ion) as positive control, or no addition as unstimulated control. T-cell proliferation was evaluated using the MTT method, and the OD values were read at 570 nm by a plate reader (Magellan; Tecan Austria GmbH, Grödig, Austria) after 4 h of incubation with MTT (12). Data were expressed as the stimulation index (SI), calculated as the mean reading of triplicate wells stimulated with antigen, divided by the mean reading of triplicate wells with medium only.
In-vivo cytotoxic assay
Splenocytes from naïve BALB/c mice were pulsed with 10−6 M HBsAg CTL peptide S208-215, and served as target cells after labeling with a high concentration of CFSE (2.5 mM, CFSEhigh) purchased from Molecular Probes (Eugene, OR). A portion of the same cells was alternatively labeled with a low concentration of CFSE (0.25 mM, CFSElow) without a peptide pulse, and served as the non-target control. The target and control cells were mixed in a 1:1 ratio and injected into immunized mice at 2×107 total cells per mouse via the tail vein on day 7 after the third immunization. The labeled cells were recovered from the spleens of mice 4 h later and examined for their differential CFSE fluorescence intensities using the FACS Calibur (BD Biosciences, San Jose, CA). Specific lysis was calculated using the following formula: percentage of specific lysis=[1 − (ratio unprimed/ratio primed)×100], where the ratio is defined as the percentage of CFSElow cells divided by the percentage of CFSEhigh cells in immunized or non-immunized mice.
Real-time PCR
Splenic B cells were isolated from immunized mice on day 15 after the first immunization using anti-B220-coated magnetic beads (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer's instructions. The purity was routinely >90% as indicated by flow cytometric analysis. Total mRNA was isolated and cDNA was synthesized by reverse-transcription PCR. SYBR Green incorporation quantitative real-time PCR was performed using the Fast Start SYBR Green mix (Roche Diagnostics, Indianapolis, IN), in the ABI7400 Sequence Detection System (Applied Biosystems, Foster City, CA). The primers used are listed in Table 2.
On day 15 after the first immunization, total mRNA was isolated from splenic B cells, and cDNA was synthesized by reverse-transcription PCR.
Flow cytometric analysis
Single-cell suspensions were prepared from mouse spleens on day 7 after the third immunization, and Fc receptors were blocked with Fcγ mAb. Cells were stained with anti-B220-APC, anti-CD27-PE, or anti-CD138-PE, for 30 min at 4°C, and then washed with ice-cold PBS. For the intracellular cytokine assay, lymphocytes were isolated on day 7 after the third immunization. T cells at 0.5×106 cells/20 μL were stimulated in 96-well plates with HBsAg (5 μg/mL), or S208-215 (ILSPFLPL; H-2Kb-restricted, 10 μg/mL), and anti-CD28 (5 μg/mL) mAb for 6 h at 37°C and 5% CO2. Monensin (2 μg/mL) was added for the last 4 h, and the cells were washed three times with PBS/10% FCS. The cells were blocked with 1 μL of Fcγ mAb (0.5 μg/mL) for 30 min at 4°C, and fixed with 4% paraformaldehyde at 4°C for 15 min. The cells were permeabilized with 0.1% saponin at 4°C for 10 min, then immunostained for 30 min at 4°C with pairs of labeled antibodies: anti-CD4-APC and anti-IL-4-PE; anti-CD8-FITC and anti-IFN-γ-PE; anti-CD4-APC and anti-IFN-γ-PE; anti-CD4-APC and anti-IL-9-PE; anti-CD4-APC and anti-IL-17A-PE; anti-CD4-APC and anti-IL-21-PE; or with isotype controls. Data were acquired and analyzed using a FACS Calibur with Cell Quest software (BD Biosciences). Dead cells were excluded on the basis of low forward-light scatter (FSC) and propidium staining. All fluorescent-conjugated anti-mouse monoclonal antibodies were purchased from BD PharMingen (San Diego, CA).
Statistical analysis
Data were analyzed using the one-sided Student's t-test. Differences were considered statistically significant at p<0.05.
Results
Expression of DNA constructs in NIH/3T3 cells
A single copy of the HK-1 coding sequence, or a series of three copies with five glycine residues between each, was cloned downstream of proVAX/S2 in-frame with S2 antigen. The correct insertion of proVAX/S2, proVAX/S2-HK-1, and proVAX/S2-3HK-1, was indicated by restriction enzyme digestion and further confirmed by sequence analysis (data not shown).
In order to confirm the expression of plasmids in a eukaryotic system, pEGFP-N3 (positive control), proVAX/S2-EGFP, proVAX/S2-HK-1-EGFP, and proVAX/S2-3HK-1-EGFP were transfected into NIH/3T3 cells. The expression of EGFP was detected under a fluorescence microscopy 48 h after transfection. Positive green fluorescent signals were clearly seen in the cells transfected by pEGFP-N3, proVAX/S2-EGFP, proVAX/S2-HK-1-EGFP, and proVAX/S2-3HK-1-EGFP (Supplementary Fig. S1; see online supplementary material at
Effects of HK-1 on IgG antibody production
To evaluate whether HK-1 has the ability to enhance a humoral response to HBsAg DNA vaccines and thus is suitable for use as a molecular adjuvant in mice, mice received intramuscular injections of the proVAX/S2, proVAX/S2-HK-1, proVAX/S2-3HK-1, proVAX vector, or saline solution at 2-wk intervals for a total of three times. On days 14, 28, and 42 after the initial immunization, serum IgG antibodies against HBsAg were analyzed by quantitative ELISA. Compared to the group immunized with proVAX/S2, a significantly elevated level of HBsAg-specific antibodies was seen in the groups immunized with plasmids incorporating a single copy of HK-1, and an even higher level was observed in the presence of triple copies of HK-1 (Fig. 1a). These data show that use of HK-1 as an adjuvant in combination with HBsAg DNA vaccine can enhance the humoral response in mice. The amount of HBsAg-specific antisera in immunized mice on day 7 after the final boost was further quantified by comparison against an International Reference Unit (IU) for HBsAg. As depicted in Fig. 1b, the amounts of antigen binding to polyclonal antibodies were significantly higher in the proVAX/S2-HK-1 and proVAX/S2-3HK-1 groups. These results suggested that these two DNA vaccines elicited good anti-HBsAg antibody responses.
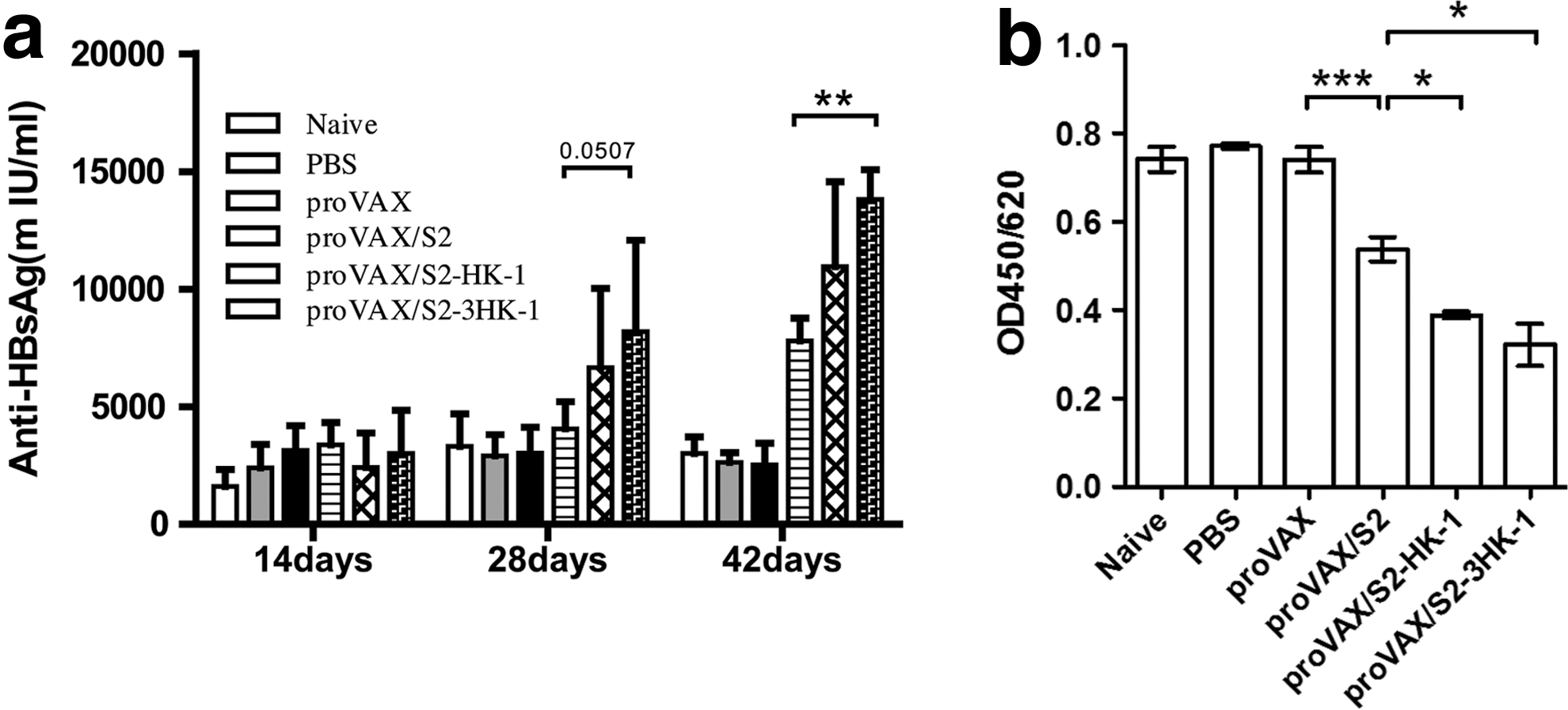
Analysis of the humoral response and antigen-antibody affinity. (
HK-1 enhances plasma cell differentiation
Since HK-1 was reported to play an important role in the terminal differentiation of activated B cells into Ab-secreting plasma cells (9), it was reasonable to test if using HK-1 as an adjuvant increased the humoral response by enhancing B-cell differentiation. Splenocytes isolated from the mice on day 15 after the initial immunization were double-stained with anti-B220 and anti-CD138. B220−CD138+ cells, defined as Ab-secreting plasma cells, were analyzed by FACS. As shown in Fig. 2a, there was a limited number of B220−CD138+ cells in non-immunized mice. While these cells remained low in mice immunized with proVAX/S2, vaccination with proVAX/S2-HK-1 and proVAX/S2-3HK-1 resulted in two- and fourfold increases, respectively (Fig. 2a and b). These data suggest that HK-1 as an adjuvant in combination with HBsAg DNA vaccine could influence plasma-cell differentiation.
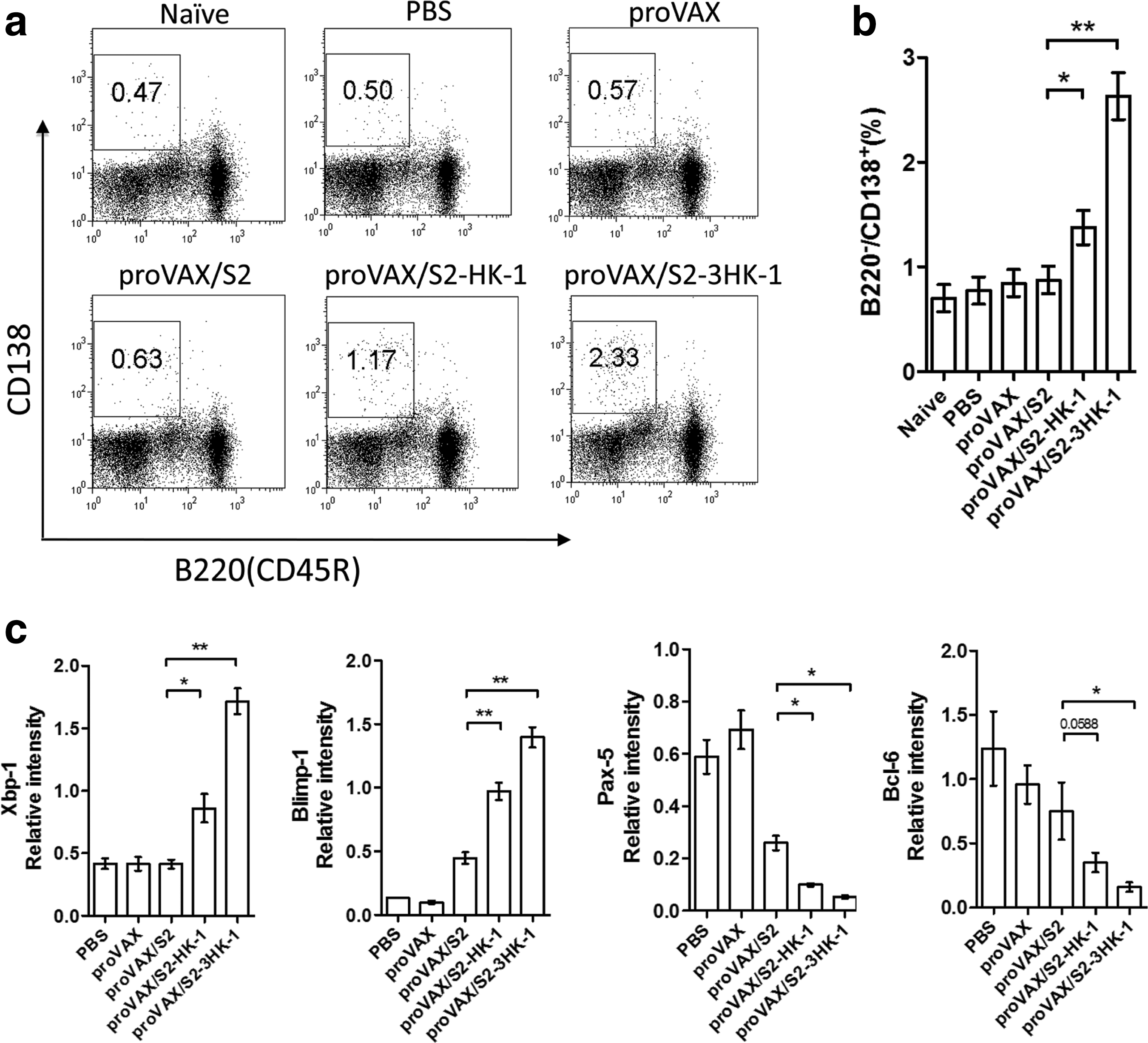
Effect of HK-1 on B-cell activation and plasma cell differentiation. Splenocytes were isolated from the mice on day 15 after the initial immunization, and double stained with anti-B220 and anti-CD138. The percentages of Ab-secreting plasma cells are shown in each dot-plot, and the gate was set on B220−CD138+ cells or events (
Differentiation of plasma cells depends on the coordinated expression of several transcription factors, including the positive regulators Blimp-1 and Xbp-1, and the negative regulators Bcl-6 and Pax-5. The expression of these transcription factors was analyzed by real-time PCR using RNA extracted from isolated B cells on day 15 after the initial immunization. As depicted in Fig. 2c, the expression of Blimp-1 and Xbp-1 were markedly upregulated, whereas those of Bcl-6 and Pax-5 were found to be decreased in B cells prepared from mice immunized with proVAX/S2-HK-1 or proVAX/S2-3HK-1 in comparison to proVAX/S2. Hence the use of HK-1 as a molecular adjuvant could modulate expression of several key transcription factors, thereby enhancing plasma-cell differentiation for Ab production.
Effects of HK-1 on immunological memory
Since stimulation of CD27 generates a signal that helps B cells to commit to the memory lineage in the mouse (13), the B220+CD27+ subset of the B-cell population was analyzed by flow cytometry after immunization. As shown in Fig. 3a and b, a small number of this subset was observed in the group immunized with proVAX/S2, whereas a higher number was seen in the proVAX/S2-HK-1 group, and an even higher number in the proVAX/S2-3HK-1 group.

Effects of HK-1 on immunological memory. (
Memory B cells have the ability to rapidly produce Ab upon secondary Ag exposure (14). Mice previously immunized by the DNA vaccines were challenged with HBsAg on day 60 after the third immunization, and the serum samples were collected on days 60 and 70. IgG antibodies against HBsAg were analyzed by quantitative ELISA. Compared to the proVAX/S2-immunized group, a significantly higher level of HBsAg-specific IgG was obtained in the mice immunized with proVAX/S2-HK-1, with the highest level seen in the proVAX/S2-3HK-1 group (Fig. 3c). This result suggests that use of HK-1 as an adjuvant could facilitate the memory response.
Modulation of cytokine expression by HK-1
Cytokines play a central role in the modulation of immune responses upon infection or immunization. We analyzed whether administration of the various DNA constructs affected the profile of cytokine production by T cells. Intracellular cytokine staining showed no significant difference among the various groups in the percentage of IL-4- and IFN-γ-producing CD4+ T cells (Fig. 4a and b), and IFN-γ-producing CD8+ T cells (Fig. 4c). Low but similar levels of IL-9 and IL-17 production were also observed for CD4+ T cells (data not shown). We did see a slightly increased number of CD4+IL-21+ T cells in the proVAX/S2-HK-1 group, which was further elevated to reach statistical significance in the proVAX/S2-3HK-1 group (Fig. 4d).
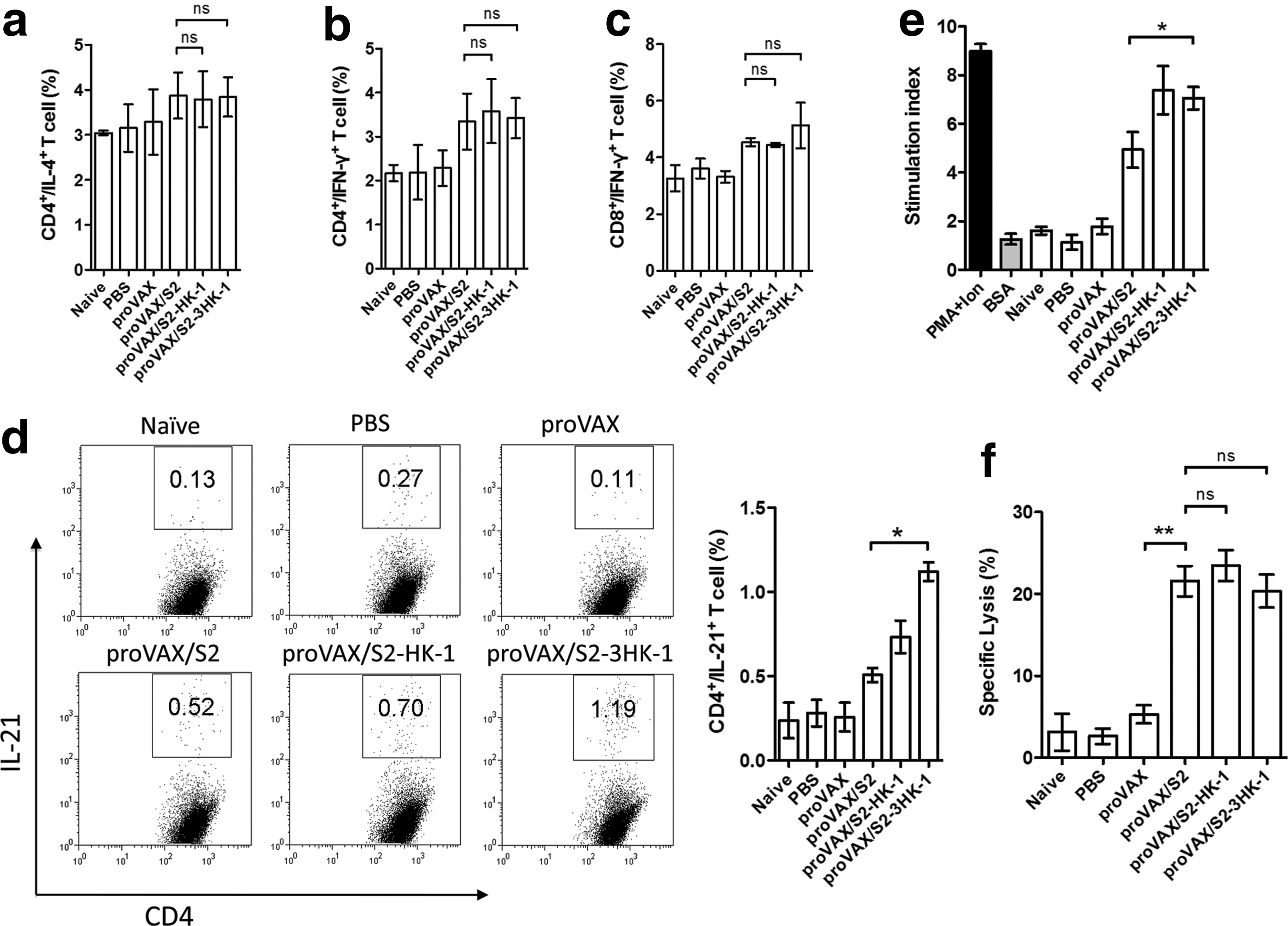
Effects of HK-1 on cytokine production, T-cell proliferation response, and in vivo CTL responses. (
HK-1 as adjuvant enhances the T-cell proliferative response
To further evaluate whether the use of HK-1 as an adjuvant could influence antigen-specific lymphocyte proliferation, splenocytes were isolated on day 7 after the last immunization, and re-stimulated in culture with HBsAg, or BSA (as irrelevant antigen), or PMA plus ionomycin (as a positive control). The proliferative response seen in the group immunized with proVAX/S2 was significantly lower than in the groups immunized with either proVAX/S2-HK-1 or proVAX/S2-3HK-1 (Fig. 4e), indicating that the use of HK-1 as an adjuvant can enhance antigen-specific T-cell responses.
The in-vivo cytotoxic response
To examine whether the use of HK-1 as an adjuvant can influence an antigen-specific cytotoxic response, we transferred CFSE-stained, antigen peptide-labeled and non-labeled splenocytes into mice that had been immunized three times. As shown in Fig. 4f, immunization with proVAX/S2 induced antigen-specific cytotoxic activity, leading to the lysis of splenocytes pulsed with HBsAg CTL peptide S208-215. In contrast to most other aspects of the HBsAg-triggered immune response, the addition of HK-1 showed no impact on the induction of cytotoxic T cells.
Discussion
In this study, we have demonstrated that HK-1 has the potential as a molecular adjuvant to enhance humoral responses obtained with an HBsAg DNA vaccine. Compared to the immune responses induced with proVAX/S2, immunization with plasmids containing preS2 plus HK-1 induced a higher level of IgG production (Fig. 1a), and more vigorous T-cell proliferation (Fig. 4e). Most importantly, from a prophylactic of view, our vaccine also induced a higher level of Ab production upon secondary Ag exposure (Fig. 3c), indicating that using HK-1 as an adjuvant might promote immunological memory. The Th-1 indicator IFN-γ, and the Th-2 indicator IL-4, remained the same among experimental groups (Fig. 4a, b, and c), as did IL-9 and IL-17. However, both of the vaccines could induce higher antigen-specific CTL activities and T-cell responses than the control groups, indicating that the DNA vaccine could stimulate cellular immune responses effectively. Compared to mice immunized with proVAX/S2 or proVAX/S2-HK-1, proVAX/S2-3HK-1 induced a higher level of IgG production, a higher percentage of differentiated antibody-secreting plasma cells, and a higher level of T-cell proliferation. Cusi et al. demonstrated that DNA-elicited immune responses could be enhanced and modulated by fusion antigens to the ligands for APC receptors (15). We hypothesize that the effect of HK-1 occurs via a novel signal pathway on B cells, and that the affinity between three copies of HK-1 and its unambiguous receptor is greater than that of one copy, resulting in its more pronounced effects. The mechanisms of this interaction should be further investigated.
The level of IL-21 was significantly increased in the groups immunized with plasmids containing preS2 plus HK-1 (Fig. 4d). Follicular helper T (Tfh) cells are CD4+ T cells that help B cells, and are important in humoral immunity (16). IL-21 produced by Tfh cells is an important regulator of Tfh cell development and B-cell responses (17). Hence our results suggest that HK-1 might have enhanced humoral immunity through an influence on Tfh cells.
An ideal vaccine should be capable of eliciting strong responses in both humoral and cellular immunity. DNA vaccines can induce cytotoxic T lymphocyte (CTL) responses that do not usually occur after immunization with protein subunit vaccines (18). However, when compared with conventional vaccines, DNA vaccines are often found to be less potent for the induction of antibody responses. Wang et al. showed that B cells cultured with HK-1 peptide showed enhanced proliferation, survival, differentiation into plasma cells, and Ab production (9). In the present study, we have demonstrated for the first time that when the HK-1 coding sequence is subcloned in-frame and downstream of S2 HBsAg in the proVAX-S2 construct in single or triple copies, the resulting vaccines induce increased amounts of IgG (Fig. 1a). Furthermore, the induction of long-lasting memory responses against HBsAg-specific antigen was greatly enhanced (Fig. 3c).
Memory is an adaptive property of the immune system that derives from a variety of mechanisms, and is characterized by recall responses that are faster and more intense than and qualitatively different from primary responses (19). In part these effects are caused by increased precursor frequency of antigen-specific T and B cells (20,21). However, the study of murine memory B cells has been limited by small cell numbers and the lack of a definitive marker (22). Unambiguous identification of memory B cells has been a major obstacle to further investigation. CD27 is a lymphoid cell-specific tumor necrosis factor receptor (TNFR) that is expressed on a small proportion of murine B cells. Hence signaling through this receptor can help to commit these B cells into the memory lineage (13). Our results showed that a vaccine expressing HK-1 in addition to preS2 antigen induced increased numbers of the B220+CD27+ subset of cells compared with the vaccine expressing the antigen alone. The inclusion of HK-1 also resulted in a more rapidly rising IgG titer upon secondary Ag exposure. The effects were possibly mediated through stimulation of signaling through CD27 with consequential enhancement of memory B-cell differentiation, but further investigations should be carried out to clarity this. Wang et al. showed that HK-1 could influence B220+IgM−IgD− memory B-cell proliferation (9). However, another possibility is that memory B cells were stimulated directly by HK-1. This supposition remains to be further investigated.
Conclusion
To summarize, this study describes for the first time the use of HK-1 as an adjuvant molecule incorporated into HBsAg DNA vaccines. It resulted in stronger humoral and memory responses. Thus, HK-1 can reasonably be predicted to be a molecular adjuvant used to enhance the humoral response of HBsAg DNA vaccine against hepatitis B infection.
Footnotes
Acknowledgments
This work was partly supported by the National High-Tech 863 Project of China and the National Nature Science Foundation (30771602). We wish to thank Dr. Douglas Lowrie for his critical review of the manuscript. and Dr. Jane Q.L. Yu and Mr. Zhonghuai He for their assistance in this work.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.