
Abstract
Marek's disease (MD) is caused by Marek's disease virus (MDV). Various vaccines including herpesvirus of turkeys (HVT) have been used to control this disease. However, HVT is not able to completely protect against very virulent strains of MDV. The objective of this study was to determine whether a vaccination protocol consisting of HVT and a Toll-like receptor (TLR) ligand could enhance protective efficacy of vaccination against MD. Hence, chickens were immunized with HVT and subsequently treated with synthetic double-stranded RNA polyriboinosinic polyribocytidylic [poly(I:C)], a TLR3 ligand, before or after being infected with a very virulent strain of MDV. Among the groups that were HVT-vaccinated and challenged with MDV, the lowest incidence of tumors was observed in the group that received poly(I:C) before and after MDV infection. Moreover, the groups that received a single poly(I:C) treatment either before or after MDV infection were better protected against MD tumors compared to the group that only received HVT. No association was observed between viral load, as determined by MDV genome copy number, and the reduction in tumor formation. Overall, the results presented here indicate that poly(I:C) treatment, especially when it is administered prior to and after HVT vaccination, enhances the efficacy of HVT vaccine and improves protection against MDV.
Introduction
Herpesvirus of turkeys (HVT) vaccine is the first successful vaccine that was developed against MD, and is currently used worldwide for vaccination of commercial flocks (11,47). In addition, other vaccines such as SB-1 and Rispens, as well as combinations of the above, are available and widely used for conferring protection. Despite the significant success of MD vaccines, MD outbreaks still take place in various countries (7). These outbreaks are probably due to different factors such as stress, suppression of the immune system by other pathogens, and evolution of the virus (43). It is also important to note that although vaccines are highly effective at preventing clinical signs of disease, they are less capable of controlling virus replication and shedding, especially in feathers (3,7).
Several vaccination protocols have recently been developed that incorporate chicken cytokines in order to alleviate some of the shortcomings of the currently available vaccines, especially in terms of their efficacy against highly virulent pathotypes of MDV. For example, the efficacy of a recombinant HVT vaccine expressing chicken interleukin (IL)-2 was evaluated. However, expression of IL-2 failed to improve the protective efficacy of HVT vaccine (46). In contrast, the use of chicken myelomonocytic growth factor improved the efficacy of Marek's disease vaccines (19). In addition, co-treatment of chickens with HVT and a plasmid construct expressing chicken IFN-γ resulted in enhancement of protection conferred by the vaccine (27).
Toll-like receptors (TLRs) are innate receptors that recognize pathogen-associated molecular patterns (PAMPs) (10). TLRs have been characterized in chickens (10,31), and TLR3 is speculated to be involved in antiviral immune responses via its activation by viral double-stranded RNA or its analog [such as polyriboinosinic polyribocytidylic [poly(I:C)]] (34). TLR ligands have been used successfully as vaccine adjuvants in various species, including chickens. For instance, TLR ligands improved immunity against influenza viruses in mice and monkeys (29,30), as well as metapneumovirus subtype C in turkeys (15). In addition, administration of a TLR ligand incorporated into a virosome-based influenza vaccine in chickens improved immunogenicity and efficacy of the vaccine (38). To date, the use of adjuvants such as TLR ligands in conjunction with MDV vaccines has not been investigated. The objective of the present study was to determine whether immunization of chickens with HVT, along with treatment with poly(I:C), could enhance protective efficacy of the vaccine. Here we have demonstrated that administration of poly(I:C) with HVT does enhance the protective efficacy of this vaccine against challenge with a very virulent strain of MDV.
Materials and Methods
Experimental animals
Specific-pathogen free (SPF) eggs were received from the Canadian Food Inspection Agency (CFIA, Ottawa, Canada). The eggs were hatched in Arkell Research Station of the University of Guelph. Day-old chicks were placed in the Animal Isolation Unit, University of Guelph. Experimental protocols were approved by the Animal Care Committee, University of Guelph.
Experimental design
Seventy-two SPF birds were randomly divided into 6 groups. Cell-free HVT vaccine (MD-Vac-CFL; Fort Dodge Animal Health, Fort Dodge, IA) was administered at one-quarter of the recommended dose via the aerosol route using a method described previously (5) on the day of hatch. This is the only cell-free Marek's disease vaccine that is commercially available, and therefore can be potentially used via the respiratory route. We reasoned that administration of a vaccine via the respiratory route might elicit mucosal immune responses leading to immunity against initial phases of virus entry, because MDV enters the host through the respiratory route. A reduced dose of the vaccine was used in the present study, because we wanted to observe the full potential of poly(I:C) when used as an adjuvant. Furthermore, although this vaccine is not commonly used in Canadian hatcheries, other Marek's disease vaccines are normally used at reduced doses. On day 5 post-hatch, 5 groups of chicks were infected with 250 plaque-forming units of MDV-RB1B, and one group received sham treatment (vaccine diluent+uninfected skin) via the respiratory route. On day 4 post-hatch and 14 dpi, birds were treated with 200 μg/bird of poly(I:C) (P0913; Sigma-Aldrich, Toronto, Ontario, Canada) via the aerosol and intramuscular routes (IM), respectively. We reasoned that administration of poly(I:C) by 4 d post-hatch may potentiate the immune response generated by HVT, and may even steer the response towards a T-helper (Th-1) response, as demonstrated in other species (35). Also, poly(I:C) administered 1 d prior to virus challenge on day 5 post-hatch may exert direct antiviral activity against the challenge virus. We selected 14 dpi because this is presumably the time of reactivation of MDV from its latency. The dosage of poly(I:C) was chosen based on pilot experiments (unpublished data). In the negative control group, chickens were treated with HVT diluent and uninfected skin. In summary, the groups in the in vivo trial were: group 1: vaccinated with one-quarter dose of HVT (day of hatch) and treated with poly(I:C) (4 d post-hatch) and infected with RB1B (5 d post-hatch); group 2: vaccinated with one-quarter dose of HVT (day of hatch) and treated with poly(I:C) (4 d post-hatch) and infected with RB1B (5 d post- hatch), and additionally treated with poly(I:C) (14 d post-infection via IM injection); group 3: vaccinated with one-quarter dose of HVT (day of hatch) and infected with RB1B (5 d post-hatch), and treated with poly(I:C) (14 d post-infection via IM injection); group 4: vaccinated with one-quarter dose of HVT (day of hatch) and infected with RB1B (5 d post-hatch); group 5: infected with RB1B (5 d post-hatch); and finally group 6: treated with HVT diluent and uninfected skin. Feather tips were collected at 5, 10, and 21 d post-infection and kept in RNAlater® (Invitrogen, Toronto, Ontario, Canada). Birds from each group were euthanized according to the University of Guelph Animal Care Committee guidelines. The spleen of each bird was collected at 21 dpi and kept in RNAlater solution until further use.
To gain a better understanding of the responses of chickens to HVT, we conducted an in vitro experiment, which involved stimulation of spleen cells from chickens belonging to various treatment groups with inactivated HVT. Briefly, 60 SPF birds were randomly divided into three groups. Cell-free HVT vaccine (MD-Vac-CFL) was administered at one-quarter of the recommended dose via the aerosol route using a method described previously (5), on the day of hatch. At 4 d post-hatch, a group of birds was treated with 200 μg/bird of poly(I:C) (P0913; Sigma-Aldrich) via the aerosol route. In the negative control group, chickens were treated with HVT diluent. Birds were euthanized following the same protocol as that mentioned above at 4, 10, and 21 dpi. Spleens were collected and mononuclear cells were isolated and cultured in plates using the methods previously described (40). Then mononuclear cells (5×106 cells/treatment) from each group were treated in vitro with two doses of irradiated HVT (the dosage was previously determined for the in vitro treatments, unpublished data), cell culture medium (negative control), or concanavalin A (Con A; 20 μg/treatment; positive control). Cells were collected and resuspended in Trizol (Invitrogen) at 24 h post-treatment. In summary, the groups were as follows: birds that were vaccinated with one-quarter dose of HVT and splenic mononuclear cells of these birds were treated with medium (HVT+M), irradiated HVT vaccine (HVT+HVT), or ConA (HVT+ConA). Another group of birds was vaccinated with one-quarter dose of HVT and treated with poly(I:C) on day 4 post-hatch. Splenic mononuclear cells of these birds were treated with medium [(HVT+poly(I:C)+M)], irradiated HVT vaccine [(HVT+poly(I:C)+HVT], or ConA [(HVT+poly(I:C)+ConA]. Finally, the control birds that were treated with HVT vaccine diluent isolated splenic mononuclear cells of these birds were treated with medium (control+M), irradiated HVT vaccine (control+HVT), or ConA (control+ConA). Although the mononuclear cells of all three groups were treated with ConA, only the data for ConA-treated cells from the HVT+ConA group are shown in Fig. 4.
DNA and RNA extraction and reverse transcription
DNA was extracted from spleens using a method described previously (1). RNA was extracted from spleens and splenic mononuclear cells using Trizol following the manufacturer's instructions. The quality as well as the quantity of RNA was estimated using NanoDrop® ND-1000 spectrophotometry (NanoDrop Technologies, Wilmington, DE). Reverse transcription of total RNA (500 ng for the cells to 1 μg for the tissues) was carried out using Oligo(dT)12–18 primers (SuperScript™ First-Strand Synthesis System; Invitrogen Life Technologies, Carlsbad, CA) following the manufacturer's instructions.
Design of primers for innate defense genes
Three genes, interferon (IFN)-γ, interleukin (IL)-10, and viral gene meq, were selected as the target genes for relative quantification in relation to β-actin in the spleen. MDV genome copy number was quantified in 100 ng of DNA in spleen and feather tips. IFN-γ, IL-2, and granzyme A expression were relatively quantified in relation to β-actin in mononuclear cells in vitro as well. The primer design and relative quantification were done as previously described (2,4).
Real-time PCR
Each real-time PCR assay was carried out with a dilution of the standard that was utilized as calibrator. All the real-time PCR assays were conducted in 384-well plates (Roche Diagnostics GmbH, Mannheim, Germany) in a final volume of 20 μL of LightCycler® 480 SYBR Green I Master (Roche Diagnostics GmbH) that contains FastStart Taq Polymerase, reaction buffer, dNTP mix, SYBR Green I dye, and MgCl2 in a LightCycler 480 instrument (Roche Diagnostics). In addition, the reaction contained 0.25 μM of each gene-specific primer, 5 μL of a 1:10 dilution of cDNA, and 3 μL of PCR-grade water.
The optimum thermal cycling parameters varied according to the gene, and consisted of denaturation at 95°C for 10 min; 40 cycles (β-actin, IL-10, and IFN-γ), 45 cycles (meq, IL-2, and granzyme A) of amplification at 95°C for 10 sec, 64°C for 5 sec (55°C/5 sec for IL-10), 72°C for 10 sec (72°C/7 sec for meq and 72°C/5 sec for IL-10); melting curve analysis at 95°C/1 sec (95°C/5 sec for meq) (segment 1), 65°C/15 sec (65°C/1 s for meq) (segment 2), and 95°C/1 sec, except for meq, which needed 97°C/1 sec (segment 3), and cooling at 40°C/30 sec.
Data analysis
The efficiency of real-time PCR and relative quantification of target genes was calculated based on methods previously described. Briefly, the expression of genes was calculated relative to the expression of β-actin based on Pfaffl's formula (41). The expression ratios of target genes were subjected to analysis of variance (ANOVA), two-tailed Student's t-test, and Fisher's exact test, by Minitab software as well as SAS 9.1 (SAS Institute Inc., Cary, NC). These data are presented as comparisons between treated and control groups, and were considered significant at p≤0.05.
Results
MD tumor incidence in poly(I:C)-treated and HVT-vaccinated chickens
To monitor the presence of MD lymphoma in internal organs, the birds were euthanized and necropsied at 21 dpi. Ninety-one percent of birds challenged with the virus that were not vaccinated developed MD tumors (group 5), while no MD tumor was observed in the control group (group 6; no challenge). The incidence of MD tumors in the birds that received HVT and additional poly(I:C) treatments (group 2) was the lowest (41%) among the challenged groups (and was significantly lower compared with group 5; p=0.02). MD tumor incidence in the birds vaccinated with HVT only (group 4) was 80%. The groups of birds that received poly(I:C) (before or after vaccination; groups 1 and 3) plus HVT demonstrated lower tumor incidence (64–66%) than birds receiving vaccine only (80%). There was no significant difference in MD tumor incidence in the birds receiving poly(I:C) with the vaccine (group 1), or after vaccination (group 3; Fig. 1).

Tumor incidence in the study groups at 21 days post-infection (dpi). Tumor incidence in the different groups was calculated by observing gross tumors in visceral organs of birds at 21 dpi. The groups were as follows: group 1: ¼ HVT+poly(I:C)+challenged; group 2: ¼ HVT+poly(I:C)+challenged+poly(I:C); group 3: ¼ HVT+challenged+poly(I:C); group 4: ¼ HVT+challenged; group 5: challenged; group 6: HVT diluent+uninfected skin. The data are the percentages of tumor incidence in 12 biological replicates in each group at 21 dpi. “a” denotes a significant difference (p≤0.05).
MDV genome copy number in feather follicles
Since MDV replicates in feather follicles and generates infective cell-free virus in this tissue, MDV genome copy number was quantified by measuring the amount of meq gene in DNA extracted from feather tips. There was a gradual increase in the MDV genome copy number in feather tips of all infected chickens throughout the course of the experiment. There was no significant difference among groups within each time point. However, the MDV genome copy number was significantly higher at 21 dpi compared to 10 dpi (p<0.001; the MDV genome copy number was lower than the sensitivity of our test at 4 dpi; the lowest copy number that could be detected by the assay was 340 copies; therefore, an arbitrary value [100 copies] that was lower than the test's cut-off point was used; Fig. 2).
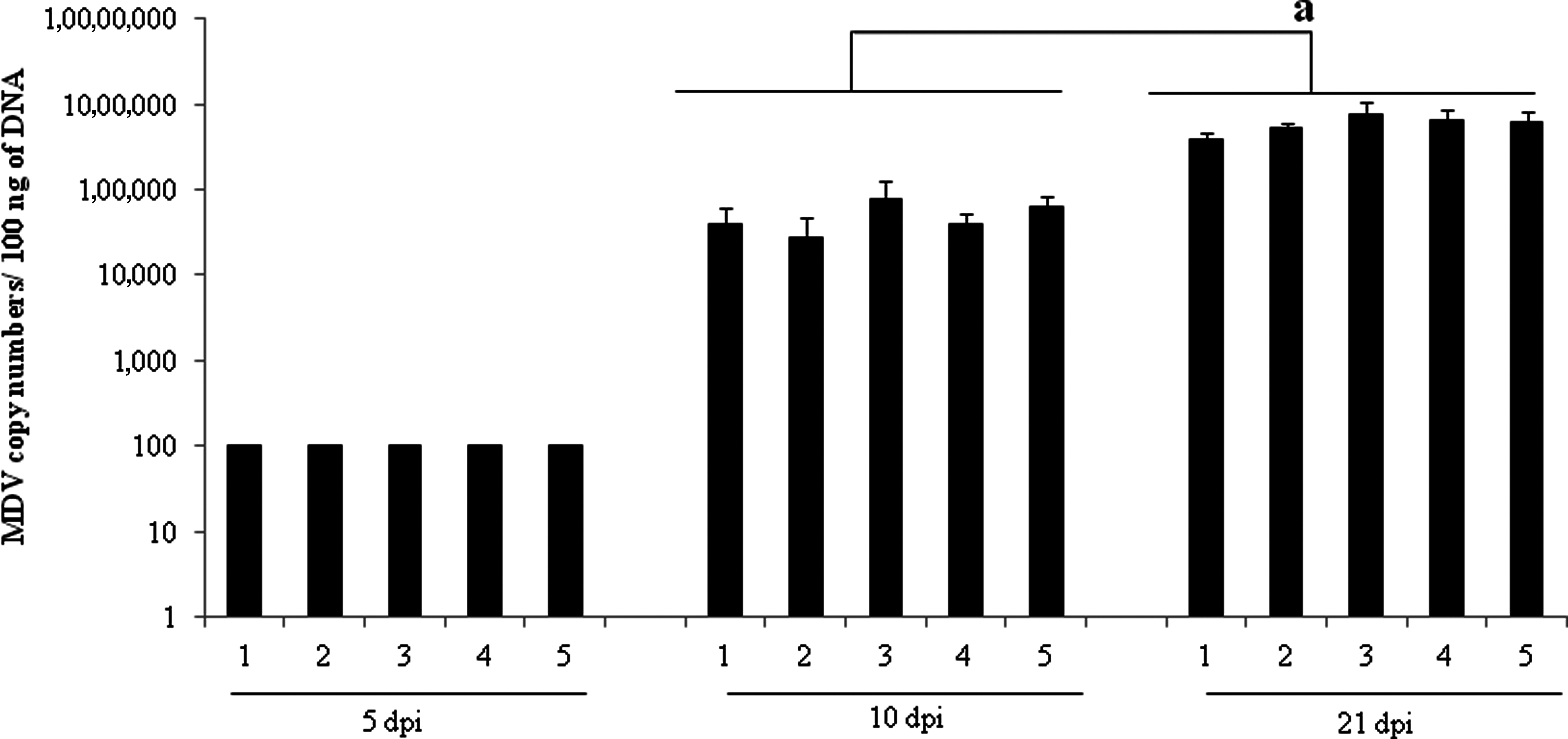
MDV copy number in feather follicles at 5, 10, and 21 dpi. The meq gene is the major oncogene of MDV that was used to calculate the virus copy number. The MDV copy number was calculated in 100 ng of DNA. The data represent the mean±standard error for 12 biological replicates at each time point. “a” denotes a significant difference (p≤0.05).
Relative expression of meq and meq genome copy numbers in spleen
The meq gene is the major oncogene of MDV, and it plays a key role in the transformation of T cells. There was no statistically significant difference among the infected groups in terms of MDV genome copy numbers and relative expression of meq (Fig. 3a and 3b). However, the expression of meq and MDV genome copy number was lower in the vaccinated group that received poly(I:C) twice (group 2).

(
IFN-γ and IL-10 relative expression in spleen
IFN-γ and IL-10 were used as indicators of the host response. IFN-γ indicates the activation of CD4+ and CD8+ T cells, whereas IL-10 may indicate the presence and activation of regulatory T cells. On the other hand, the expression of IL-10 was significantly higher in groups 1, 3, 4, and 5, compared to groups 2 and 6 (p<0.001) at 21 dpi (Fig. 3c and 3d).
IFN-γ, IL-2, and granzyme A expression in splenic mononuclear cells
IFN-γ and IL-2 expression as an indicator for the reactivation of primed CD4+ and CD8+ T cells was assessed. There was no statistically significant difference between the different groups for expression of IFN-γ. However, a trend was observed in cells from HVT-vaccinated groups that were re-stimulated in vitro with irradiated HVT. These cells tended to express more IFN-γ than cells from control birds that were stimulated with HVT, or cells from HVT-vaccinated birds that were treated with medium only. The abundance of IL-2 transcripts was increased in the HVT+HVT group at 4, 10, and 21 days post-treatment (dpt). The upregulation in expression of IL-2 in the HVT+HVT group was significantly higher at 4 dpt (p=0.02). On the other hand, IL-2 expression was increased significantly in the [HVT+poly(I:C)]+HVT group at 21 dpt compared to the [HVT+poly(I:C)]+M group (p=0.03; Fig. 4b). Moreover, the expression of granzyme A, an indicator of reactivation of primed CD8+ T cells, was increased in the [HVT+poly(I:C)]+HVT group at 21 dpt, which was significantly higher compared to the [HVT+poly(I:C)]+M group (p=0.04; Fig. 4c). In brief, the difference between the [HVT+poly(I:C)]+HVT and HVT+HVT groups in terms of the expression of IFN-γ, IL-2, and granzyme A was not statistically significant.
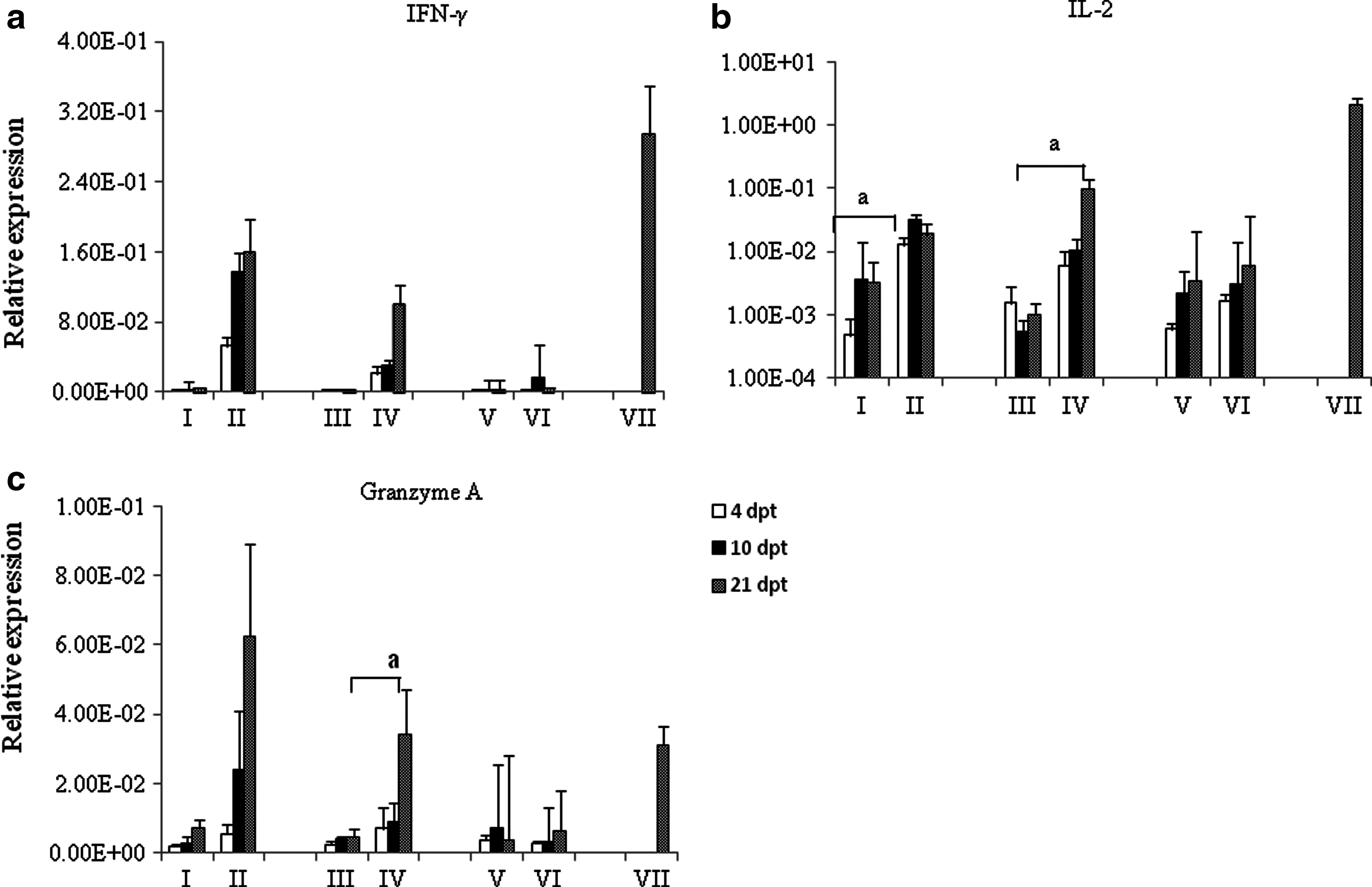
(
Discussion
TLR ligands are promising candidates for the development of novel vaccine adjuvants. Several TLRs, including TLR3, are known to interact with nucleic acids of pathogens. In the case of TLR3, interactions between this receptor and double-stranded RNA or its analog, poly(I:C), results in induction of type I interferons in both mammalian and avian species (34,44). It has also been shown that TLR3 ligands such as poly(I:C) induce dendritic cell maturation, and stimulate the release of proinflammatory cytokines, a prerequisite for the initiation of adaptive immune responses (8,36). In the present study, we explored the use of poly(I:C) as an adjuvant to enhance the efficacy of a vaccine against MDV. The results demonstrate that the administration of poly(I:C) in combination with HVT improves protection against the incidence of MD tumors in vaccinated chickens. Two different mechanisms may be involved in the inhibitory effects of poly(I:C) on MD tumor development: (1) poly(I:C) may enhance the quality or/and quantity of antigen-specific immune responses induced by the vaccine; or (2) antiviral or anti-tumor activity of poly(I:C) may inhibit viral infection/replication or tumor growth in the treated chicken. Regardless of the mechanisms involved in protection, poly(I:C) treatment did not inhibit viral shedding from the infected chickens. Moreover, the protection conferred by poly(I:C)/HVT vaccine treatment was not associated with an upregulation of IFN-γ expression in the spleen of the treated chicken.
The expression of IL-10 was significantly lower in the group that received HVT vaccine and additional poly(I:C) treatments. The results demonstrate that a decrease in IL-10 expression is associated with lower tumor incidence. IL-10 is a regulatory cytokine that inhibits the expression of inflammatory cytokines such as IFN-γ, which is involved in conferring protection against viruses or tumors (22). It is well known that IL-10 promotes the development of Th-2-type responses both in mammals (37) and chickens (42), a type of response shown to suppress effective antiviral/tumor immunity. In addition, it has been reported that MDV induces the expression of IL-10 both in the lytic and latent phases of infection (28,45). Furthermore, Buza and Burgess hypothesized and provided some evidence that the elevation in IL-10 expression in the course of MDV infection is probably a strategy used by the virus to elude and hamper the Th-1 immune response (12). Therefore, lower expression of IL-10 in our study may be an indicator of a protective immune response against MDV in these chickens. In addition, a decrease in IL-10 expression may simply be related to the fact that there is less tumor formation in poly(I:C)-treated birds.
Although MDV copy number and replication were numerically lower in group 2 (the group with the lowest tumor incidence), the difference with other groups was not statistically significant. One possible explanation for this is the possibility that the effects of poly(I:C) might have been transient. Based on the available evidence, poly(I:C) has the ability to inhibit herspesvirus replication (23). Therefore it is conceivable that poly(I:C) has similar effects on MDV. Another alternative, but not mutually exclusive scenario, is the leukopenic effects of poly(I:C). It has been demonstrated that administration of poly(I:C) could lead to depletion of T cells (32). T cells are the targets for virus-induced transformation. Therefore, it is possible that at least one of the effects of poly(I:C) administration was reducing the number of T cells, hence reducing T-cell transformation and formation of lymphomas without influencing virus load and replication. On the other hand, MDV vaccines cannot prevent the transmission of the virus via feather dander, despite the presence of an immune response elicited by different vaccines in this tissue (3,4). We also observed that there was no difference in terms of MDV copy number in the feather follicles among the various groups. Therefore, administration of poly(I:C) with the vaccine was not able to prevent the generation of cell-free MDV that leads to spread of the virus. However, further work needs to be done to confirm this observation.
In general, Marek's disease vaccines can confer partial protection against very virulent strains of MDV, or nearly complete protection against virulent strains of MDV, without curtailing virus replication and shedding from feathers of infected chickens (7). Our results support these findings and demonstrate that poly(I:C) treatment reduces tumor development without reducing viral replication and load. It has been shown that some TLR ligands can directly exert anti-tumor effects. For example, it has been demonstrated that poly(I:C) treatment reduces tumor formation in a mouse prostate tumor model (17). It is possible that poly(I:C) directly inhibits MD tumor growth, and this effect contributes to the anti-tumor effects of adaptive immunity conferred by the HVT vaccine. The mechanisms involved in the anti-tumor effects of poly(I:C) are not fully understood. However, it has been reported that poly(I:C) may induce autophagy in tumor cells by expression of inflammatory cytokines, or apoptosis through a RIP/FADD/caspase-8–dependent pathway (33). Further studies are required to demonstrate anti-tumor effects of poly(I:C) on tumor growth in MDV-infected chickens.
To further unravel the mechanisms involved in protective immunity elicited by poly(I:C) in vaccinated chickens, splenic mononuclear cell responses to in vitro re-stimulation with HVT was examined. We used IFN-γ, IL-2, and granzyme A expression as indicators of re-activation of antigen-specific primed T cells (18,25). Poly(I:C) can act directly via TLR3 (8,24), or indirectly through induction of apoptosis in tumor cells (36), to promote dendritic cell (DC) maturation in mammals. Activation of DCs leads to the release of cytokines such as IFN-γ and IL-12, which in turn results in activation of antigen-specific CD4+ Th-1 cells in mammals (25). Activated CD4+ T cells can also produce IFN-γ in response to viral infections under the influence of mature DCs and macrophages. We noted an upregulation of IL-2 transcripts in the HVT+HVT group at 4 and 10 dpt. The increase in IL-2 expression was also noted in the [HVT+poly(I:C)]+HVT group at 21 dpt, that was higher compared to the [HVT+poly(I:C)]+M group (26). IL-2 is a key player in the expansion of both CD4+ (9) and CD8+(16) T cells. IL-2 is also essential in the formation of memory CD4+ (21) and CD8+ (20) T cells. Taken together, we show here that in vitro re-stimulation of cells from HVT-vaccinated birds with inactivated HVT results in cellular activation marked by expression of cytokines, and to some extent, granzyme. However, poly(I:C) did not appear to significantly enhance cell-mediated immune responses in vitro. Although cytokine or granzyme expression may be regarded as surrogate markers of cell-mediated immune responses, it would have been ideal to investigate whether poly(I:C) in conjunction with vaccination can enhance cytotoxicity of effector T cells against virus-infected or tumor cells. However, there are no reliable or reproducible assays currently available for chicken T cells.
In conclusion, we have demonstrated for the first time an enhancement in protection conferred by HVT vaccine when a TLR3 ligand is concomitantly administered in chickens. However, understanding the underlying mechanisms by which TLR ligands exert their effects requires further experimentation. Finally, it is essential to learn the optimal vaccine regimen in terms of time and method of delivery of TLR ligands with MDV vaccines.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Acknowledgments
Payvand Parvizi was a recipient of an Alexander Graham Bell Canada Graduate Scholarship (CGS). This study was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC)-CRD Partnerships Program, the Poultry Industry Council, and the Saskatchewan Chicken Industry Research and Development Fund. We would like to thank Dr. Douglas Hodgins for his help with the statistical analysis of the data. We would also like to acknowledge the staff of the animal isolation facility of the University of Guelph.