
Abstract
Pigs were immunized with DNA plasmids containing different open reading frames (ORFs) of a porcine reproductive and respiratory syndrome virus (PRRSV) genotype I strain. One group was injected with three inoculations of ORF7, a second group was immunized with three inoculations of plasmids containing ORF5 and ORF6, and a third group was kept as controls. Later, +21 days after the last inoculation, animals were challenged with the homologous strain. After the challenge, PRRSV-specific interferon (IFN)-γ-secreting cells and anti-PRRSV IgG antibodies developed faster in DNA vaccinated pigs (p<0.05). However, DNA-immunized pigs showed an exacerbation of the disease compared to the unvaccinated challenged pigs. The data suggest that previous immunization with DNA vaccines against glycoprotein 5 and/or matrix protein of PRRSV, as well as nucleoprotein but to a lesser degree, could result in an exacerbation of the clinical course in terms of fever upon challenge.
Introduction
Currently, two types of commercial vaccines against PRRSV are available, modified live-virus vaccines (MLVs) and inactivated vaccines (IV). To date, MLVs seem to offer the strongest immunity (23,46), although, even in this case, protection is often partial (25). Other approaches such as subunit vaccines based upon plasmid DNA or bacteria (18) have not been commercialized yet.
Taken into account the feasibility of DNA immunization and its protective capacity against viral infection in domestic pigs (11,35), we determined to use this methodology as a tool to analyze the potential of different ORFs for inducing an effective immune responses associated with protection. Therefore, the main objective of the present study was to evaluate DNA constructs in plasmids containing ORF5 plus ORF6 or ORF7 of genotype I PRRSV as potential vaccines. After the immunization period, pigs were challenged with the homologous virus. The clinical course, lesions, and the humoral and cellular immune responses of pigs were determined.
Material and Methods
Construction of the plasmid vectors expressing ORF5, ORF6, and ORF7
The L-450 (VP21) strain used for the challenge and for DNA constructions was grown in porcine alveolar macrophages. The strain has been characterized previously (6,7); sequences of ORFs 2–7 can be found in GenBank (accession numbers DQ009647, DQ009648, DQ009653, DQ009656, DQ009657, and DQ064785). Primers used for plasmid constructions enabling the amplification of PRRSV ORFs 5 and 7 with restriction sites have been described previously (8); whereas ORF6 primers were designed for the purposes of the present work (Table 1). Expression of the viral proteins coded by the plasmids was verified by means of an immunoperoxidase monolayer assay (IPMA) (40) in BHK-21 and in MARC-145 cells using monoclonal antibodies for GP5 or N protein (Inmunología y Genética Aplicada S.A., Madrid, Spain; Ingenasa, 1.1 PR5. K3), or a polyclonal serum for M. All DNA preparations used in the experiments were endotoxin free, as determined using a commercial kit (Qiagen).
Experimental design
Twelve healthy 4-week-old Landrace PRRSV and porcine circovirus type 2 (PCV2)-free pigs (determined by serology and reverse transcription-polymerase chain reaction (RT-PCR) for PRRSV and by PCR for PCV-2) were located in the experimental biosafety level 3 facilities of Centre de Recerca en Sanitat Animal (CreSA) and randomly allocated to three groups (A, B, and C). Group A was inoculated with the plasmid encoding ORF7 (400 μg in 1.2 mL sterile phosphate buffered saline, PBS) and B with the plasmid encoding ORF5 plus the plasmid encoding ORF6 (400 μg of each plasmid in 1.2 mL sterile PBS). Group C animals were kept as controls, with each pig receiving 1.2 mL of the unaltered plasmid pcDNA3.1+. Pigs were immunized again at 21 day intervals for a total of three inoculations. In each administration, two-thirds of the total quantity of DNA was injected intramuscularly (IM), and one-third of the DNA was injected subcutaneously (SC), as reported before (12). Twenty-one days after the third immunization (day 0 post-infection (PI); namely +63 days since the first inoculation), pigs were challenged intranasally with 2 mL containing 106 TCID50/ml strain L-450 PRRSV.
Sample taking and clinical follow-up
Immediately before the third inoculation and during the challenge period, blood samples were collected in siliconized and heparinized tubes (days -21, 0, 3, 7, 10, 14, and 21 PI and days -21, 0, 7, 14, and 21 PI, respectively). After the challenge, pigs were examined clinically and body temperatures were recorded daily from day 0 to 21 PI. Pigs were also weighed weekly. At day 21 PI, pigs were euthanized and a post-mortem examination was performed. All experimental procedures were approved by the Animal Ethics Committee of the Universitat Autònoma de Barcelona (n° 3987).
Detection of viremia
Viremia was assessed by viral isolation in PAM and by nested RT-PCR (Nrt-PCR). Viral isolation and titration was done as reported elsewhere (9). Briefly, serum samples were serially diluted from 100 until 10−4, inoculated in PAM cultures (50 microliters in quadruplicates) and incubated at 37°C in 5% CO2 for 1.5 h. Then, sera were removed and culture medium was added. Isolation of the virus was verified at day 3 PI in PAM by means of IPMA (40). PRRSV was detected in sera by Nrt-PCR as previously reported (6). The first-round PCR has a sensitivity of about 10–50 TCID50/mL serum, whereas Nrt-PCR had a sensitivity of 1 TCID50.
Humoral immune responses
A commercially available ELISA (HerdChek PRRS 2XR, Idexx Laboratories) was used for measuring anti-PRRSV IgG antibodies; sample to positive (S/P) ratios>0.4 were considered to be positive. Antibodies against GP5 were measured using a competitive home-made ELISA kindly provided by Ingenasa, that uses recombinant genotype I GP5 as antigen for coating plates and a specific-GP5 AcM labeled with peroxidase. Percentage of competition was calculated using the formula: ((Odneg – Odsample)/(Odneg – Odpos))×100. Positive and negative controls were provided by the manufacturer. Samples were considered as positive when percentage of competition was>21%. Neutralizing antibodies (NA) were measured as described before (22,44) with minor modifications (7). Infection of cell cultures was revealed by IPMA (40).
Cell-mediated immunity measured by interferon-γ ELISPOT
Peripheral blood mononuclear cells (PBMC) were separated from blood by density-gradient centrifugation with Histopaque 1.077 (Sigma). The interferon (IFN)-γ ELISPOT was performed as described previously (5,45) using commercial mAbs (Porcine IFN-γ P2G10 and biotin P2C11; BD Biosciences Pharmingen). PBMC were stimulated with strain L-450 at a multiplicity of infection (MOI) of 0.01 as a recall antigen. Unstimulated cells and phytohemagglutinin-stimulated controls (10 μg/mL) were included in the assay. To calculate the antigen-specific IFN-γ-secreting cells (IFN-γ-SC), average counts of spots in unstimulated wells were subtracted from average counts in antigen-stimulated wells. Results were expressed as the number of PRRSV-specific IFN-γ-SC per 106 PBMC.
Cytokine ELISAs for interleukin-1, interleukin-8, and tumor necrosis factor-α
PBMC were seeded at a density of 2×106 cells/well (250 μL) in 96-well plates and were mock-stimulated or stimulated with strain L-450 (MOI 0.01) for 24 h. Cytokine concentrations of interleukin (IL)-1β and tumor necrosis factor (TNF)-α from culture supernatants of PBMC, and sera were measured using commercial pairs of antibodies (porcine IL-1β and IL-8, BD Biosciences Pharmingen; TNF-α, RD Systems) and expressed as pg/mL.
Histopathology and immunohistochemistry
Samples from pigs were collected during postmortem examinations and were processed histopathologically and for PRRSV antigen detection by immunohistochemistry (IHC). Severity of interstitial pneumonia and follicular lymphoid hyperplasia (submandibular, tracheobronchial, mesenteric, and superficial inguinal lymph nodes) were semi-quantified according to previously described procedures (28). Detection of PRRSV by IHC was done using monoclonal antibody against N protein (Ingenasa) and semi-quantified (16). In order to avoid biases, these examinations were done in a blinded fashion by two pathologists.
Statistical analysis
The regression line of cytokine standards in ELISA was calculated by using FC Junior software (Biotek Instruments). Statistical comparisons between groups were done by using Statsdirect v2.4.1: Mann-Whitney/Kruskal-Wallis for means of temperature, anti-PRRSV IgG antibodies, GP5 antibodies, cytokines and IFN-γ-SC; χ2 tests for sum of febrile days; and Fischer's exact test for proportion of positive pigs by ELISA (IgG antibodies).
Results
Clinical course
After challenge, all pigs developed mild signs of disease consisting of dullness and fever; a few of them—from all groups—developed slight dyspnea. Regarding temperatures, all pigs had fever (>40°C) at least once during the first 2 weeks PI. The highest mean of temperatures corresponded to pigs in group B at day 9 PI. Also, significantly higher temperatures compared to controls (group C) were recorded in group B at days 4, 6, 9, and 10 PI (p<0.05) (Fig. 1A). In order to have a further picture for body temperatures, proportions of daily observations in which pigs were feverish were compared among groups (Fig. 1B). Thus, group C accounted for six positive records (11%) out of a total of 56 potential observations (4 pigs×14 days), whereas A and B summed 9 (16%) and 16 (29%), respectively. Thus, the number of positive observations/total number of observations in B was also significantly higher (p<0.05).
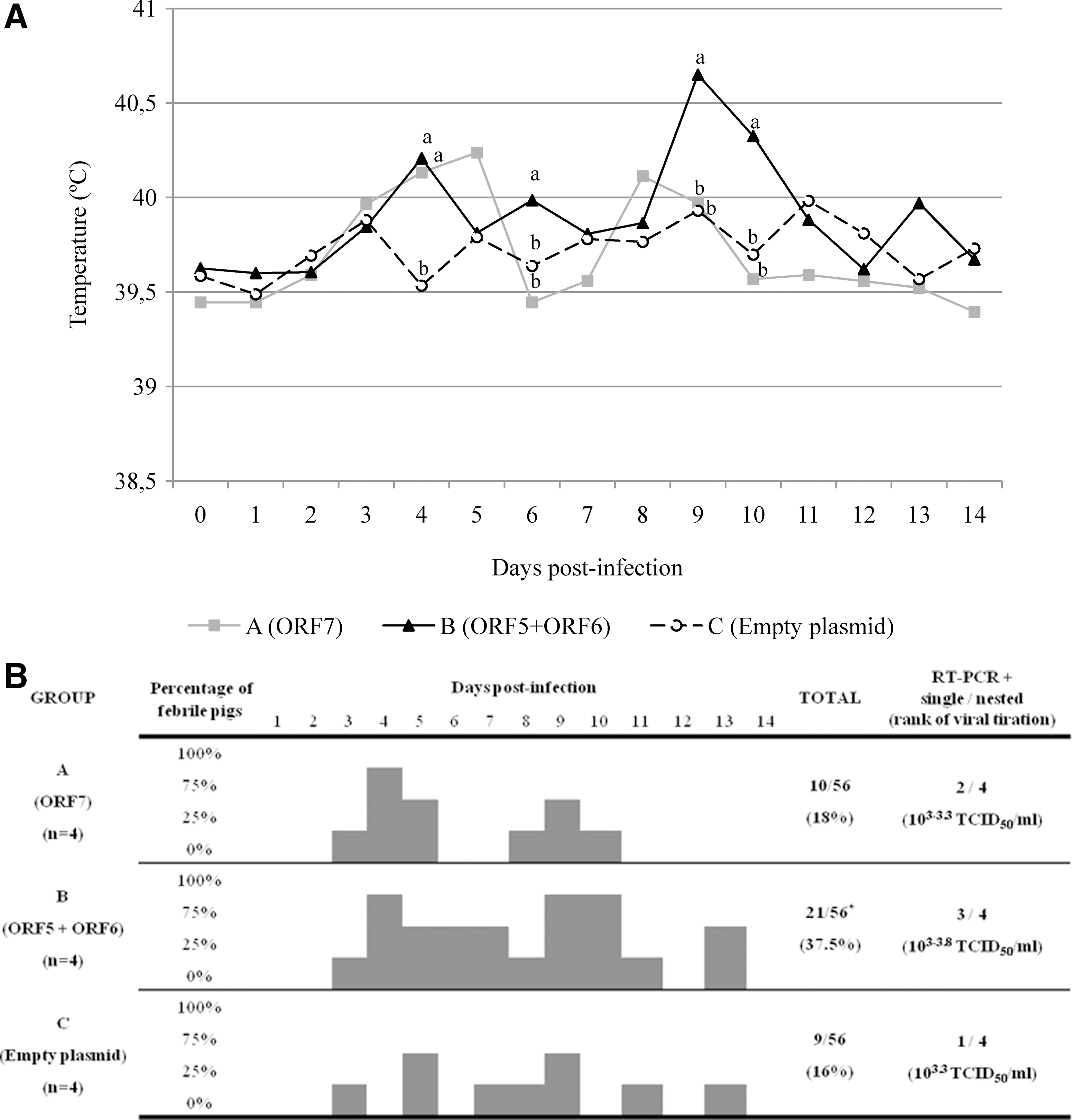
Evolution of body temperatures and viremias after inoculation with PRRSV. Body temperatures were measured at 24 h intervals during the first 14 days post-inoculation (PI). All pigs had fever (>40°C) at least once during the first 2 weeks PI.
Regarding weight gains, although significant different weight gains were observed in a given group when comparing intervals of time (Fig. 2), there were no significant differences among groups (p>0.05).
Individual means and differences of daily weight gain of pigs from 0 to 7 PI and from 7 to 14 days PI in groups A (ORF7), B (ORF5+ORF6), and C (empty plasmid). *Differences between intervals in all the groups (p<0.05). Means and standard deviations for body weights were: A=33.75±3.23, B=37.02±2.00, and C=35.47±4.83 for day 0 PI (p>0.05); A=40.45±3.40, B=43.32±2.13, and C=41.30±4.60 for day 7 PI (p>0.05); A=43.87±4.09, B=45.52±2.52, and C=44.90±3.77 for day 14 PI (p>0.05).
Viremia
After challenge, some animals were detected as positive using viral isolation, with titers from 103 to 103.8 TCID50/mL. These same pigs were also positive by a single-round of RT-PCR: one pig in group C, two in group A, and three in group B (not significant; p>0.05) (Fig. 1B). All pigs were shown to be viremic by Nrt-PCR.
Humoral response
According to the ELISA PRRS 2XR results, all animals were negative to PRRSV before the challenge except for two pigs in group A (day 0 PI; mean S/P ratio=0.45±0.05). At day 7, PI all pigs belonging to group A had already seroconverted, while pigs in groups B and C were still negative. Later, all pigs from all groups seroconverted. Until the end of the experiment, average S/P ratios of group A were nearly always significantly higher than those of groups B and C (p<0.05) (Table 2). Regarding antibodies against GP5 (Table 3), all pigs were negative until 14 PI. At 21 PI, all pigs in group B were positive (mean of percentage of competition=54.7±6.9), whereas in group A one pig still remained negative (mean for positive pigs=37.3±10.8). In group C, only two pigs were positive by 21 PI (mean for positive pigs B>A and C at day 21 PI; p<0.05). None of the animals included in the study developed NA before or after the challenge (day 0, 7, and 21 PI).
Development of the anti-PRRSV IgG antibodies (HerdChek PRRS 2XR; Idexx Laboratories) expressed as number of positive pigs, the mean of S/P ratio±SD and range for positive pigs.
Different superscript letters indicated statistically significant differences among groups for a given date (P<0.05).
Development of antibodies against GP5 (1.1 PR5. K3; Ingenasa) expressed as number of positive pigs, the mean of percentage of competition±SD and range for positive pigs.
Different superscript letters indicated statistically significant differences among groups for a given date (p<0.05).
Interferon-γ ELISPOT
Before the challenge, only a few vaccinated pigs showed low IFN-γ responses in ELISPOT. However, after the challenge PRRSV-specific IFN-γ-SC rapidly rose in DNA vaccinated pigs (average IFN-γ-SC frequencies at 7 PI: A=330±175; B=230±31, and C=84±70; A=B>C; p<0.05). This trend towards greater responses in previously vaccinated pigs was maintained until the end of the experiment (A=B>C from day 7 to day 21 PI; p<0.05) (Fig. 3).
Frequencies of PRRSV-specific IFN-γ-SC in 106 PBMCs±SD measured by ELISPOT. Different superscript letters indicated statistically significant differences among groups for a given date (p<0.05).
Cytokine ELISAs
In vitro stimulation of PBMC with strain L-450 only produced significant differences for IL-1β at day 21 PI. At that time, pigs belonging to group C were negative for IL-1β, whereas three-quarters of pigs in both DNA vaccinated groups were positive (Group A: 207±212 pg/mL; Group B: 323±298 pg/mL) (p<0.05).
Regarding serum samples, sporadic IL-1β responses were observed in group C (one-quarter of pigs at 3, 10, and 14 PI). In vaccinated pigs, group B reached significantly higher responses than A at 3 and 7 PI (A=172±233 pg/mL versus B=1051±1072 pg/mL at day +3 PI; A=162±135 pg/ml versus B=715±615 pg/mL at day +7 PI) (B>A; p<0.05). Later on, only sporadic responses were observed in those groups. When serum levels of IL-8 were measured, all pigs were positive without differences among groups. For TNF-α, positive results were sporadic.
Gross and microscopic lesions
Gross interstitial pneumonia was observed in one pig from group C, in all pigs belonging to group B, and in two pigs from group A (not significant; p>0.05). Microscopically, group C pigs showed less severe lesions of interstitial pneumonia than DNA vaccinated pigs (not significant; p>0.05) (Table 4). Mild to moderate diffuse follicular lymphoid hyperplasia was observed in all pigs (data not shown, not significant; p>0.05).
Pathological and intensity was scored as normal (-), mild (+), moderate (++ ) and intense (+++).
Proportion of pigs in each group, sum, sum of individual scores, and sum of scores by group.
Proportion of pigs in each group, sum, sum of individual scores, and sum of scores by group.
ACC, accessory pulmonary lobe; API, apical pulmonary lobe; DIA, diaphragmatic pulmonary lobe; MED, medium pulmonary lobe; pneumonia, degree of interstitial pneumonia.
Detection of PRRSV by immunohistochemistry
PRRSV was detected by IHC in lungs of two pigs from group C, in one in B, and three from A (not significant; p>0.05) (Table 5). Viral antigen was also detected in most lymph node samples regardless of the group (Table 5).
PRRSV antigen detection was scored in order to measure the staining in tissues: no positive cells (−), single positive cells (+), scattered groups of positive cells (++), widespread positivity (+++).
ACC, accessory pulmonary lobe; API, apical pulmonary lobe; DIA, diaphragmatic pulmonary lobe; MED, medium pulmonary lobe; Mesenteric (MES), submandibular (SUB), superficial inguinal (ING), and tracheobronchial (TR) lymph nodes.
Discussion
Proteins encoded by ORFs 5–7 of PRRSV—namely GP5, M, and N are the major structural proteins of the virus and are reported to elicit cell-mediated responses although of different magnitude (2, 8, 20, 38). In addition, GP5 is thought to contain a major neutralization epitope (30). For these reasons, those genes and gene products have been the most commonly used in studies reporting immunization of pigs with subunit or DNA-based PRRSV vaccines (18). In the present study, pigs were vaccinated with either ORFs 5 and 6 or ORF 7 and then were subjected to a homologous challenge in order to evaluate the efficacy of this strategy of immunization.
The results obtained in this work showed that a barely detectable humoral response devoid of NA were only detected in group A (ORF 7) before the challenge, whereas the response during this period was null in those pigs immunized with ORF 5 plus ORF6. Regarding cell-mediated immunity before the challenge, it was low in both vaccinated groups. After the challenge, all ORF 7 vaccinated pigs showed anamnestic responses in terms of humoral immunity, whereas only a priming might have occurred in some animals from group B; regarding cell-mediated immunity, all pigs showed an anamnestic response. These phenomena of barely or null responses before and anamnestic responses after the challenges, has been also reported in other studies using DNA vaccines (1,12,24,32,43). Despite the priming induced by the DNA vaccination, in the present case no significant protection was observed upon challenge, indicating that the immunity induced was not relevant enough in terms of protection. Other experiments using DNA vaccination for PRRSV showed similar results (18). However, unexpectedly in our case, pigs vaccinated with ORF5 and ORF6 showed the highest temperatures and the highest number of observations with fever, even compared to unvaccinated pigs (p<0.05). Also, pigs in group B had the lowest weight gains from 7 to 14 PI (not significant; p>0.05) and both vaccinated groups (A and B) had the highest pathological score in the lungs (not significant; p>0.05). Interestingly, in the case of group B, the higher pathological scores in the lungs did not correspond to a higher frequency of detection of PRRSV by IHC. It has been reported that in PRRSV neither the macrophage recruitment nor the degree of histopathological lesion in lungs are correlated with the presence of viral antigen (15,42) and, therefore, lung lesions may well also be the result of not only infection of macrophages but also the consequence of an immune-mediated process.
Adverse effects have occasionally been found following the use of other genetically engineered viral vaccines, including those against equine infectious anemia (39), herpes simplex-1 (14), foot-and-mouth disease (13), and influenza (17) viruses. Particularly in the case of PRRSV, two studies have reported an adverse effect in pigs previously immunized with a GP5 subunit PRRSV vaccine (Rgp5) produced in Escherichia coli (31) or with an E. coli GST-ORF5 recombinant fusion protein (29). In the present study, results could not be conclusive at all because there were no significant differences for some of the parameters analyzed and we could not definitively explain the reason behind the exacerbation. However, similarly to us, Prieto et al. (2011) have also observed that vaccinated pigs showed a trend for higher fever and more severe clinical signs after challenge.
Several reasons could explain the findings of the present report, but for other pathogens the exacerbation of different diseases in previously vaccinated animals has been related to 1) the production of non-neutralizing antibodies associated to the antibody-dependent enhancement (ADE) and/or 2) inflammatory responses (19). In the case of PRRSV, ADE is believed to be mediated by antibodies against envelope or membrane-associated proteins, particularly against GP5 and N (3). If it occurred in the present study, then it should probably have been more accentuated in group A where anti-PRRSV IgG antibodies could already be detected in some animals before the challenge, whereas antibodies against GP5 appeared later. Nevertheless, when the parameters related to the exacerbation of the disease were individually analyzed and compared to the development of antibodies against the different proteins, there was a lack of correlation (data not shown). In whatever case, the role of ADE in the exacerbation of the disease is controversial; Pirzadeh and Dea (1998) observed more severe lesions in vaccinated pigs that developed non-neutralizing antibodies, but Prieto et al. (31) did not find this correlation. Regarding inflammatory responses, Prieto et al. (31) have suggested a role for pro-inflammatory cytokines, among them IL-1β or TNF-α. In our study, clear differences were only seen for IL-1β, which appears at higher levels in PBMC cultures and serum from vaccinated animals, particularly in group B (p<0.05). IL-1 promotes inflammation by itself and also by inducing the expression of genes of pro-inflammatory molecules (10). In PRRS and other pig infections, IL-1β has also been correlated with the development of lung lesions, anorexia, weight loss, and fever (10,26,27,36,37). Therefore, it is tempting to believe that this is the mechanism underlying the enhancement of the lesions and the clinical course observed in vaccinated animals. Also, vaccinated animals developed stronger IFN-γ responses after inoculation. To be active, IL-1β needs to be cleaved by caspase-1 whose expression is, in turn, upregulated by IFN-γ (4). As it has been suggested by Lawson et al. (26), IL-1β expression could lead to a strong Th1 cell response increasing IFN-γ.
Taken together, our data confirm previous studies where DNA immunizations primed the immune response (18), but also suggest that immunity against GP5, M, and/or N could be associated with exacerbated clinical signs, implying that this immunity might be a double-edged sword. It is uncertain what immune mechanism underlies the observed exacerbation, although cell-mediated responses, particularly IL-1β, might play a role. The present study does not exclude the use of those viral proteins in genetically engineered vaccines, but emphasizes the need for a better understanding of the immunopathogenesis of PRRS.
Footnotes
Acknowledgments
We would like to thank Núria Navarro and Esmeralda Cano for their excellent technical assistance. Competitive home-made ELISA was kindly provided by Inmunología y Genética Aplicada S.A. (Ingenasa). Dr. Ivan Díaz has been financially supported by project ‘Porcivir’ CDS2006-00007 of the Spanish Ministry of Science and Innovation. This study has been funded by the Spanish Ministry of Science and Innovation project AGL2005-07073/GAN.
Author Disclosure Statement
The authors declare that they have no competing financial interests.