
Abstract
Noroviruses (NoVs) are one of the leading causes of acute nonbacterial gastroenteritis in humans of all ages. In the 1990s, NoV genotype GII-4 became responsible for the majority of NoV sporadic gastroenteritis cases and outbreaks worldwide. Vaccine development against NoV GII-4 is underway. At the same time, there is concern of new emerging NoV genotypes, such as GII-12, which has been recently associated with increasing numbers of NoV outbreaks worldwide. The specific question is whether type-specific pre-existing immunity to NoV GII-4 might impair cognate immune response induced by new viral infections or vaccines. Using GII-4 and GII-12 virus-like particles, we tested the impact of the immunity generated against NoV GII-4 on de novo antibody responses to GII-12 in mice. We found that pre-existing immunity to NoV GII-4 did not impair de novo immune response to the novel antigen, therefore suggesting lack of original antigenic sin (OAS).
Introduction
The NoV genome encodes a capsid protein (VP1) that self-assembles into virus-like particles (VLPs) when expressed in a recombinant baculovirus-insect cell system (10). These VLPs are morphologically and antigenically similar to the native virus (10). Considerable effort has been put into the development of NoV vaccines based on VLP's (1,2,25). However, challenges to the vaccine development are high genetic diversity and the antigenic variation of NoV strains (6) and high level of NoV pre-existing antibodies (11,15), which might impair the immune response to the vaccine. We determined the impact of pre-existing immunity to GII-4 on the generation of de novo immune response to a novel NoV genotype GII-12 in a mouse model.
Materials and Methods
VLP production
The RNA (ORF-2 gene) of GII-4-1999 and GII-12-1998, extracted from NoV infected human stool samples were amplified by RT-PCR (2) and cloned in baculovirus vector (Invitrogen, Carlsbad, CA) for the production of NoV VLPs in a SF9-insect cell system (Invitrogen) (12). Purification of the VLPs was done by sucrose gradients as described previously (9). Protein purity, morphology, in vitro antigenicity, and functionality are described elsewhere (9,15).
Mice immunization
Seven-week-old female BALB/c mice (Harlan Laboratories, Horst, the Netherlands) were immunized twice (at study weeks 0 and 3) with 1 μg of GII-4 or GII-12 VLPs by intradermal (ID) route; mice were sacrificed at week 5. In other sets of experiments, groups of mice received two doses of 10 μg GII-4 by intramuscular (IM) route at the above schedule. Half of the mice receiving 10 μg GII-4 doses were boosted with 10 μg of GII-12 VLPs at study week 18, and all mice were sacrificed at week 27. No adjuvants were incorporated in immunizations. Blood samples were collected from each mouse at study weeks 0 (pre-bleed), 4, 5, 6, 8, 10, 12, 14, 16, 18, 19, 23, 25, and 27. All procedures were authorized and performed according to the guidelines of the Finnish National Animal Experiment Board.
Enzyme-linked immunosorbent assay (ELISA) and avidity assay
ELISA was used to measure antibody levels in mouse serum as described earlier (21). Briefly, GII-4 or GII-12 VLPs were coated (0.2–1 μg/mL) on a 96-well plate (Nunc-Immuno Maxisorp, Thermo Fisher Scientific Inc., Waltham, MA) prior to addition of serum dilutions. Antibodies were detected by goat anti-mouse IgG HRP and OPD-substrate (both from Sigma-Aldrich, St. Louis, MO). Termination sera were serially diluted two-fold and tested for GII-4- and GII-12-specific IgG serum titers. Serum dilution with an OD greater than or equal to 0.1 and three times the mean OD of the negative control serum was regarded as positive. Serum from mice receiving carrier only (PBS) was used as a negative control.
Antibody avidity was determined by testing individual mouse serum samples (diluted 1:200) in a modified ELISA in which 8 M urea was included in two washing steps after incubation of the serum samples in order to elute low-avidity antibodies (15,18). The avidity index was calculated as (OD with urea/OD without urea) × 100% and avidity index ≥50% was considered as high avidity.
Saliva phenotyping and HBGA blocking assays
Saliva ABO phenotyping was conducted using monoclonal antibodies (Immucor, Houston, TX) against A, B, and H antigens, as described by others (8). Saliva from secretor-positive individuals used in the blocking assays were selected by predetermined saliva-VLP binding profiles (data not shown). A saliva blocking assay (7) was conducted as described earlier (2). Briefly, GII-4 and GII-12 VLPs (0.2 μg/mL and 0.5 μg/mL, respectively) were pre-incubated with serially diluted mice antisera (week 27) and added to saliva type A (for GII-4 binding) or type B (for GII-12 binding) coated plates. The bound VLPs were detected using human NoV antiserum (15) and anti-human-HRP conjugate (Invitrogen) reacting with OPD-substrate (Sigma Aldrich).
Results
BALB/c mice were immunized twice with NoV GII-4 or GII-12 VLPs alone with a short-term immunization schedule and low dose of antigen (1 μg per dose). Immunizations with both VLPs induced strong antigen-specific IgG response with mean serum reciprocal titers 5 logs10 each (data not shown). After confirming good immunogenicity of the VLPs, mice were immunized twice with 10 μg dose of GII-4 VLPs, and the immune response was followed up until week 27 either with or without GII-12 VLP boost at study week 18. After the two doses of GII-4 VLPs, the homologous and cross-reactive IgG responses to GII-12 were stable until study week 27 (Fig. 1A and B, dashed lines). The boost with GII-12 VLPs did not impair the immune response to the primary antigen as seen from the unaltered level of anti-GII-4 IgG during the study (Fig. 1A, solid line). In addition, the final antibody titer of GII-4-specific antibodies was identical between the groups receiving the GII-12 VLP boost or not (1:51200 titer for both). Rather, GII-12-specific IgG level rose two-fold from the mean prior-boost level (OD=1.065±0.140) to the post-boost level (OD=2.180±0.151) and remained steady for the rest of the study (Fig. 1B, solid line). Anti-GII-12 mean IgG titer increased four-fold after the GII-12 VLP boost (mean antibody titer 1:51200) compared to the group receiving no boost (mean antibody titer 1:12800).
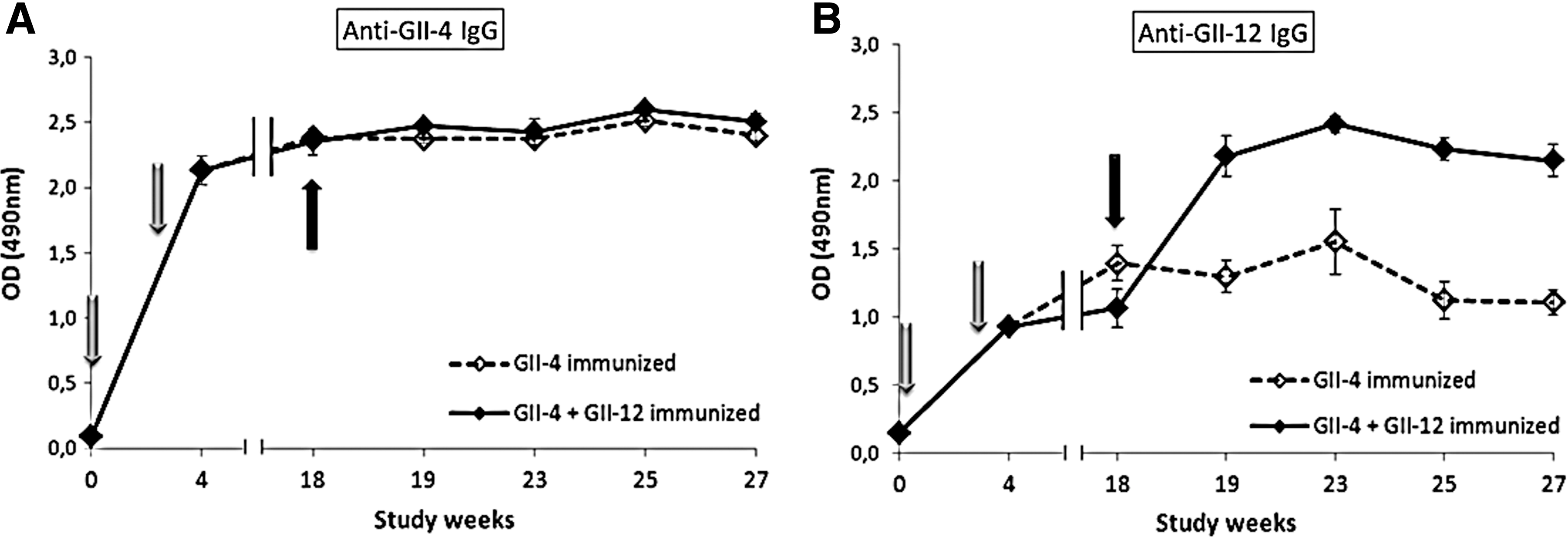
Antibody-mediated immune response induced by NoV VLPs. BALB/c mice were immunized twice with 10 μg GII-4 VLPs (light gray arrows) and terminated at study week 27. A group of mice received a 10 μg boost with GII-12 VLPs (dark gray arrows) at week 18. Dashed lines represent the mean IgG antibody levels of solely GII-4 immunized group, while solid lines represent the mean IgG levels of the GII-12-boosted group. GII-4-specific IgG
Antibody avidity of both experimental groups was high to the primary antigen GII-4 (mean avidity index 69.6±3% for GII-4 immunized group and 64.8±7.9% for the GII-12 boosted group), but only the GII-12 boosted group had anti-GII-12 IgGs with high avidity (mean avidity index 61.5±6.9%).
To show the potential neutralization of the serum antibodies, blocking assays were used in which the antisera from mice terminated at week 27 were used to block the binding of NoV VLPs to HBGA carbohydrates present in human saliva (14). Blocking of GII-4 VLP binding to HBGAs was achieved with the sera of GII-4 VLP-primed mice with or without GII-12 VLP boost at a similar level (Fig. 2A). However, GII-12 VLP binding to HBGAs was completely blocked only by antisera from the GII-12-boosted mice (Fig. 2B).
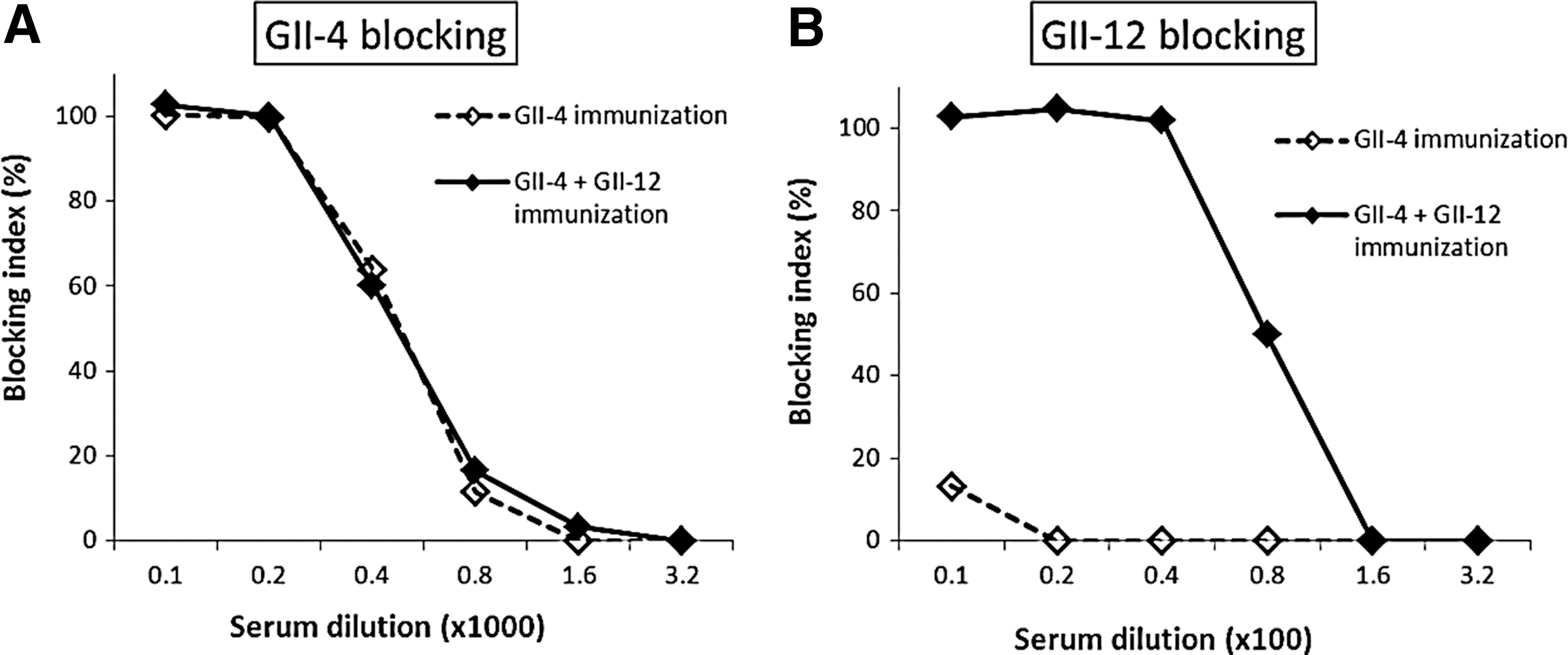
Blockage of VLPs binding to human saliva. Sera of mice immunized twice with 10 μg GII-4 VLPs without (dashed lines) or with (solid lines) GII-12 boost (10 μg) were used to block GII-4
Discussion
The NoV genotype GII-4 has been circulating and evolving for at least 4 decades, as the earliest finding of the ancestral NoV GII-4 dates back to 1974 (3). Since the early 1990s, it has become the most abundant NoV genotype, accounting for 80% of current NoV outbreaks worldwide (19). In 2009, a novel GII-12 NoV strain emerged in the United States, causing 16% of NoV outbreaks (24), and its emergence has been reported worldwide (5,16,20). Nearly all adults and the majority of children over 5 years of age are seropositive to NoV GII-4 (11,15). There is a concern (13) that pre-existing immunity may impair immune response to new emerging strains or to vaccination with NoV VLPs in similar fashion as with influenza virus and HIV infections (23). This phenomenon is known as “original antigenic sin” (OAS) in which the immune response to new infections or vaccination is impaired by the host's infection history (22). Immune responses generated upon NoV challenge were skewed to strains other than the infecting strain, giving reason to suspect OAS involvement in the NoV immunity (13). Studying OAS in NoV infections is extremely challenging because of the multiple strains of NoVs a volunteer is exposed to during the lifetime, and the variable genetic susceptibility of individuals to NoV infection (4). The mouse immunogenicity model is a useful tool to study OAS in NoV-induced immunity because the “exposure history” can be generated and controlled by the researchers.
We used two genetically distant NoV capsid-derived VLPs, GII-4 and GII-12 genotypes, to immunize naïve BALB/c mice. Both VLPs were able to induce a similar level of antibodies at a very low antigen dose (1 μg/dose). Two immunizations with 10 μg GII-4 VLPs were further used to generate long lasting immunity to primary antigen (2) prior to boosting with GII-12 VLPs. In this way we hoped to induce immunological memory specific to the primary immunogen. Our results show that a strong (reciprocal IgG titer 5 log10) GII-4-specific antibody response lasting up to 27 weeks was induced. A heterologous boost with GII-12 VLPs at 18 weeks did not reduce the IgG antibody level or the HBGA blocking activity of GII-4 antisera. Furthermore, de novo immune response to the new secondary antigen was induced as confirmed by the following: 1) GII-12-specific serum IgG content after the boost reached the same level as for the primary antigen GII-4; 2) a four-fold rise observed in the GII-12 specific geometric mean serum titer compared to the nonboosted group sera; and 3) blocking potential (100%) of GII-12 antisera to block GII-12 VLPs' binding to HBGA compared to the group not receiving the boost (<20% blocking). To further confirm that the rise in anti-GII-12 antibody response did not merely reflect the cross-reactive antibodies induced by the GII-4 immunization (2), we used an avidity assay to show the increase in the affinity of GII-12-specific antibodies. The results show that GII-4 immunization alone was not able to induce high avidity anti-GII-12 IgGs, whereas GII-12 VLP boost did increase the avidity of GII-12 specific antibodies, indicating formation of specific antibody response and functional antibody maturation to de novo antigen.
Our study in a mouse immunogenicity model shows that immunity to NoV induced by VLPs is long-lasting and protective as measured by blocking activity of the antisera. Furthermore, the pre-existing immunity to GII-4 did not impair de novo immune response generation to the novel antigen GII-12. Therefore, our results encourage the use of VLPs in NoV vaccine development as no evidence of OAS was detected; however, further studies are needed to confirm these results. Vaccination with VLPs could boost the pre-existing immunity to extend to a wide range of genotypes and likewise the circulating strains could boost the vaccine-induced immunity.
Footnotes
Acknowledgments
We thank Suvi Lappalainen for the sequencing of recombinant baculovirus stocks. We are also grateful to Eeva Jokela, Marianne Karlsberg, and Sanna Kaven for the valuable technical assistance during the study.
Author Disclosure Statement
No competing financial interests exist.