
Abstract
Besides being an important target in the antiretroviral therapy against the human immunodeficiency virus type 1 (HIV-1), the HIV-1 reverse transcriptase (RT) enzyme has potential as a vaccine antigen. In this study, we explored the ability of plasmid-encoded RT to induce cell-mediated immune responses. The strategy for increasing the immunogenicity of the protein was to delete non- or low-immunogenic parts in order to focus the immune responses to known immunogenic regions. Expression and immunogenicity of the truncated RT was compared to a clinically evaluated full-length RT construct, and the truncated RT displayed enhanced in vitro expression and cell-mediated immune responses in BALB/c and HLA-A0201 transgenic C57BL/6 mice. The strong immune responses were retained also when the truncated RT was delivered as a part of a multigene HIV-1 vaccine. Linking the RT gene to a highly expressed HIV-1 protease gene did not increase the immunogenicity of RT. This optimization strategy could be used to enhance the immunogenicity of other RT-encoding DNA vaccines.
Introduction
Here we aimed to design a highly immunogenic RT DNA vaccine using an alternative approach. Regions in the protein that have no or low numbers of known cytotoxic T lymphocyte- (CTL) and T helper epitopes were deleted from the RT gene, generating a construct encoding the most immunogenic parts of the antigen (dRT). Moreover, the use of long amino acid stretches, rather than selecting the most immunogenic epitopes alone (12), does not restrict the antigenicity to a certain human leukocyte antigen (HLA) background. We also studied whether coupling the truncated RT gene to a gene encoding a highly expressed enzymatically inactive HIV-1 PR (PRi) (13) could enhance the expression and subsequent immunogenicity of RT. The study showed that dRT was well expressed and stimulated significantly higher magnitudes of cell-mediated immune responses compared to enzymatically inactive full-length RT. The coupling of PRi to dRT did not enhance RT-specific immune responses.
Material and Methods
The dRT gene (HXB2) is devoid of nucleotide stretches encoding no or low numbers of known CTL and T helper epitopes and was manually assembled using information from the Los Alamos HIV immunology database (
Protein expression in transfected Vero E6 cells was examined by Western blot (13). Briefly, polyclonal rabbit sera to RT (16) and a monoclonal mouse antibody to PR (Clone 1696, Exbio, Praha, Czech Republic) were used in a 1:2000 dilution to detect RT and PR, respectively. Secondary horseradish peroxidase (HRP)-conjugated anti-rabbit and anti-mouse detection antibodies (Dako, Glostrup, Denmark) were subsequently used in a 1:3000 dilution. Molecular weights were calculated using the web-based Protein Molecular Weight software (
Groups of 4 female BALB/c mice (Charles River Laboratories, Sulzfeld, Germany), 5–9 weeks old, were used. Mice were immunized twice (weeks 0 and 5) intradermally at two separate injection sites with 10 μg DNA/plasmid/injection diluted in 20 μL saline (20 μg DNA in total). Directly after DNA injection, the sites were electroporated using the Derma Vax™ electroporation device (Cellectis, Romainville, France) (17). Some groups of mice were immunized with plasmids from a multigene vaccine encoding Env (subtype A, B, and C) and Gag (subtype A and B) (7), with or without the RT- and PR-encoding vaccine plasmids. Plasmids encoding Env, RT, and PR were mixed and injected at two sites on one side of the back of the mouse, and plasmids encoding Gag, RT, and PR were mixed and injected in the same manner on the other side of the back of the mouse. For the Env and Gag plasmids, 10 μg of each plasmid were used for immunizations. Groups of 5 HLA-A0201 transgenic C57Bl/6 mice (18) were immunized once with RT-encoding plasmids as described above. Mice were sacrificed 2 weeks post the last immunization and spleens and sera were collected.
Interferon-γ (IFN-γ) and interleukin-2 (IL-2) FluoroSpot (Mabtech, Nacka Strand, Sweden) assays were used as described (19). Purified splenocytes were stimulated with pools of peptides representing HIV-1 RT, PR, Gag, and Env (15 mers with ten amino acids overlap). The final concentration of peptides was 5 μg/mL/peptide. Antibody responses to PR and RT were assessed with enzyme-linked immunosorbent assay (ELISA) as previously described (13). Plates were coated with 100 μL/well of 1 μg/mL recombinant HIV-1 RT or PR (NIBSC/CFAR, Potters Bar, UK).
Amino acid sequence similarities between vaccine constructs were determined using the ALIGN software (
Results and Discussion
The aim of this study was to construct a HIV-1 RT-encoding vaccine plasmid able to induce strong cell-mediated immune responses. An RT subtype B (HXB2) gene was chosen and regions encoding known CTL and T helper epitopes were identified. Regions with no or low numbers of such epitopes, constituting 26% of the wild-type protein, were removed, resulting in a gene encoding a 413 amino acid long dRT protein (Fig 1). In an additional fusion gene, dRT was coupled to the highly expressed and immunogenic PRi (13) to form the PRidRT gene. Both genes were codon-optimized to enhance expression (20) and successfully cloned into the pKCMV expression vector. Expression levels and immunogenicity were compared with an enzymatically inactivated full length RT gene (termed RTmut) (8) that has been evaluated in clinical trials (6,7).

Alignment of RT (HXB2) amino acid sequences. Wild-type RT (RT) was compared with RT carrying mutations that inactivates the enzymatic activity (RTmut), and RT where regions with no or few CTL and T helper epitopes were deleted (dRT).
Expression of the various proteins, all being encoded by the same expression vector, showed that dRT was considerably better expressed than RTmut (Fig. 2A), this despite deleting large parts of the protein and thus potential binding sites for the RT-specific antibodies in the polyclonal sera. The pronounced difference in expression levels could be due to the deletions in RT as well as the codon optimization. Transfections with a mixture of dRT and PRi reduced the expression of both antigens, in particular RT, and this is probably due to competition between the two plasmids for the transcription/translation machinery. Similar observations have been reported for in vitro expression of HIV-1 regulatory proteins (21) and might correlate with reduced expression and immunogenicity in vivo. The PRidRT chimera was however highly expressed, suggesting that the fusion of PRi to dRT can indeed augment the expression of dRT with retained high levels of PR being detected in the Western blot assay.
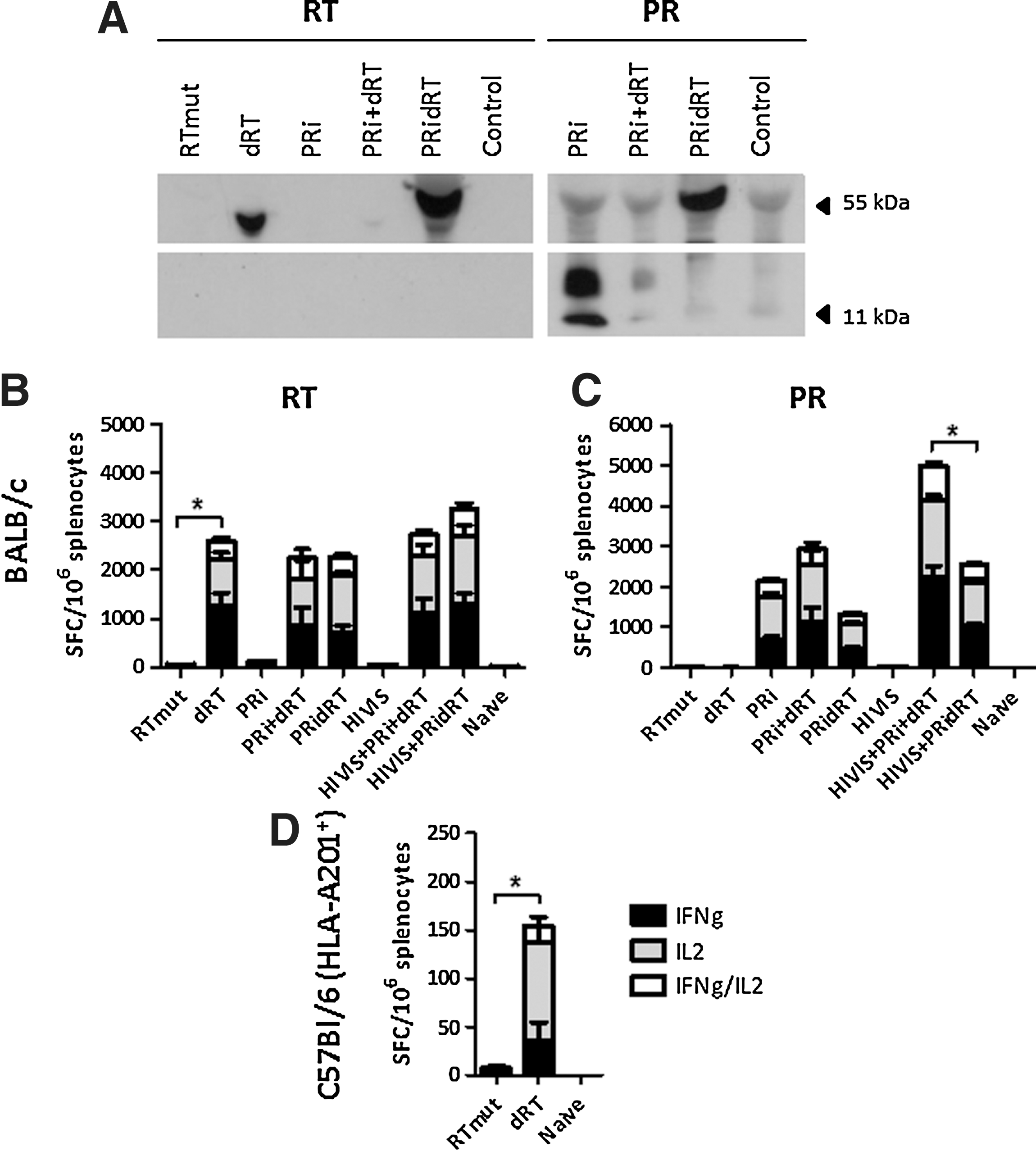
Expression and cell-mediated immune responses of RT and PR.
Intradermal immunization augmented by electroporation of BALB/c and HLA-A0201 transgenic C57Bl/6 mice with the different vaccine plasmids showed that the high level of expression of dRT correlated with significantly higher levels of IFN-γ and IL-2 secretion than RTmut (Fig. 2B–D). For BALB/c mice, the reduction of RT and PR expression seen after the co-delivery of both plasmids did not correlate with reduced RT- and PR-specific immunogenicity (Fig. 2B and C). Neither did immunizations with the highly expressed PRidRT induce enhanced immunogenicity compared to immunizations with dRT alone. This suggests that only more pronounced differences in expression levels, such as the one seen between dRT and RTmut, have an impact on the immunogenicity. The genetic linking of the two antigens did however lead to reduced PR-specific IFN-γ and IL-2 responses, especially when the plasmids were administered together with plasmids from the multigene HIVIS vaccine. We hypothesize that this is because the large dRT part (413 amino acids) of the PRidRT chimera outcompetes the small PRi part (99 amino acids) for degradation and presentation via the MHC pathways, and the fact that RT contains more CTL and T helper epitopes on the BALB/c (H-2kd) and HLA-A0201 background than PR (
We have previously observed interference between the Env and Gag antigens that are encoded by a multigene vaccine. Therefore, this vaccine is being delivered as two entities at separate injection sites in clinical trials (22). Co-administration of RT- and PR-encoding plasmids with Env- and Gag-encoding plasmids did however not impact the immunogenicity of any of the antigens (Fig. 2B, 2C, and data not shown), holding promise for the potential inclusion of the RT and PR immunogens in the multigene vaccine.
No antibodies to RT were induced in neither BALB/c nor HLA-A0201 transgenic C57Bl/6 mice. Antibodies to PR were detected in sera from BALB/c mice (data not shown). The linking of PRi to RT, but not the co-delivery of RT- and PR-encoding plasmids, hampered the induction of PR-specific antibodies. This is probably due to steric hindrance of the small PR by the larger RT antigen.
Taken together, we report a means of genetically optimizing an RT gene to enhance expression and RT-specific immune responses. This optimization strategy can most likely be applied also to other multigene HIV-1 vaccine candidates where RT is represented.
Author Disclosure Statement
No competing financial interests exist.