
Abstract
Respiratory syncytial virus (RSV) is a major cause of severe lower respiratory tract infections in infants and the elderly worldwide. The significant morbidity and mortality associated with this infection underscores the urgent need for development of RSV vaccine. In this study, we first show that intranasal administration of RSV glycoprotein core fragment (Gcf) to neonatal mice can induce systemic humoral immune responses and protective immunity against RSV without causing lung eosinophilia, although antibody response was shifted to a Th2 response. Next, we examined whether the presence of maternal anti-RSV antibodies would affect the responsiveness and protection efficacy of Gcf in newborn mice, since infants can possess RSV-specific maternal antibodies due to frequent RSV re-infections to adults. Intranasal administration of Gcf induced antibody response and increased IFNγ secretion and protected mice against RSV challenge without severe lung eosinophilia, even in the presence of high levels of RSV-specific maternal antibodies. Thus, our findings suggest that Gcf may be an effective and safe RSV vaccine during the neonatal period.
Introduction
Since the target population for RSV vaccine includes infants, relevant neonatal animal studies of current RSV vaccine candidates must precede human clinical trials. Although mice are immunologically less mature at birth and their innate immune system is more divergent than humans, mouse neonates can be a useful tool for comparative immunological research, since mice, like humans, also rely on placental transfer, with lesser dependency on colostral transfer, of maternal immunoglobulins for the establishment of passive immunity early in their life (1,11). In humans, evidence indicates that Th2-skewed immunity prevails in newborns (1). In neonatal mice, immature myeloid dendritic cells are incapable of driving the immune responses to Th1 against RSV (37). RSV infection in the neonatal period leads to substantial production of Th2 cytokines (9,26) and subsequently results in lung eosinophil infiltration, the major characteristic of airway hypersensitive responses (13). Th2-biased immune responses recur when mice are reinfected with RSV (7). Because RSV infection does not assure full protective immunity in humans, repetitive infections occur (46,47). Of importance, maternal antibodies generated by RSV infections before or during pregnancy are transmitted via placenta or breast milk, and these passively transmitted antibodies may influence the RSV vaccine-related immune responses in infants (17,25). The presence of high levels of maternal antibodies hinders the immunogenicity of vaccines for other diseases including measles, poliomyelitis, tetanus, diphtheria, and pertussis (24).
RSV F and G glycoproteins, as well as peptides derived from them, have been tested as vaccine candidates against RSV infection, as both F and G proteins induce neutralizing antibodies and, thereby, correlate with protection against RSV challenge in animal models (12,15,33). While F protein is highly conserved, G protein shows only ∼50% amino acid sequence identity between prototype RSV subgroups A and B (21). Moreover, it has been reported that there are up to 20% sequence diversity in the G protein within the subgroups (5,42). Such sequence difference within the G protein has been hypothesized to serve a crucial function in causing repeated RSV infections owing to the immunodominance of the hypervariable C-terminus region of the G protein (6,38). While a full length of G protein confers only subgroup-specific protection, a recombinant fragment containing G protein amino acid residues 130 to 230 (G2Na) has been shown to induce protective immune responses against both RSV A and B in mouse model in the context of BBG2Na fusion protein, where G2Na was fused in frame to the albumin binding domain of Streptococcal protein G (BB) (35). Importantly, the murine B-cell protective epitopes (152 to 163, 165 to 172, 171 to 187, and 196 to 204) within G2Na were also recognized in RSV-seropositive humans (32). BBG2Na has moved into a phase III clinical trial (34,35). However, there has been a report on adverse events following BBG2Na immunizations, which has led to halting that phase III trial (29). In addition, Libon et al. have reported a post-vaccine type III hypersensitivity reaction in a rabbit model, mainly due to the BB component, rather than G2Na (29). Based on these findings, further studies are required to understand the immunogenicity and protective efficacy generated by the core fragment of the RSV G protein alone.
Since RSV replication and disease are restricted to the respiratory tract, priming of local mucosal immunity may be sufficient to confer protection against RSV challenge (15). Furthermore, mucosal vaccination conveniently eliminates the need for the use of needles. We, therefore, delivered our candidate vaccine intranasally with a mucosal adjuvant, cholera toxin (CT), which has been shown to induce potent mucosal immunity when administered via nasal route together with protein vaccines and, thus, has been frequently used in various animal studies (10,19,39). Recently, Kim et al. (23) have generated an RSV G attachment glycoprotein core fragment (Gcf, a.a 131 to 230) without any fusion component. Gcf has substantial prophylactic antiviral and chemotactic effects in an adult mouse model. In this study, we extend such observations by evaluating the safety of Gcf in neonatal mice and by assessing its protective abilities when administered during the neonatal period. Here, we found that our Gcf vaccination could induce protective immunity even in the presence of maternal antibodies without causing any immunopathologic symptoms after RSV challenge.
Materials and Methods
Mice
To test for the effects of maternal RSV antibodies, we used 6- to 8-week-old female BALB/c mice purchased from Charles River Laboratories (Orient Bio, Seongnam, Korea) or Samtako Bio Korea (Osan, Korea). For immunization studies, we used neonatal BALB/c mice (2–5 days old) obtained from Samtako Bio Korea or bred under specific pathogen-free conditions at the International Vaccine Institute (IVI). All studies involving animals adhered to the guidelines of the IVI Institutional Animal Care and Use Committee (2011-014).
Preparation of antigen and RSV stocks
The recombinant RSV (RSV A2 strain) G protein fragment from amino acids 131 to 230 (Gcf) was expressed in Escherichia coli and purified as described (23). A stock of RSV A2 strain (American Type Culture Collection, Manassas, VA) was propagated in HEp-2 cells (American Type Culture Collection) in 150-mm cell culture dishes. Four days after inoculation, virus was harvested and titer was determined by plaque assay. HEp-2 cells were maintained in minimum essential media (MEM) containing Earle's salts, L-glutamine, 10% fetal bovine serum (FBS) (Hyclone, South Logan, UT), and 1% penicillin-streptomycin (Gibco, Grand Island, NY).
Immunization and challenge
One-week-old neonatal BALB/c mice were immunized with five different substances via intranasal (i.n.) or footpad routes: live RSV A2 (5×104 PFU), Gcf (20 μg), Gcf (20 μg) with cholera toxin (CT; List Biological Laboratories, Campbell, CA) adjuvant (1 μg), and PBS were administered via the i.n. route whereas FI RSV A2 (1×105 PFU) was injected into mouse footpads. Two weeks later, mice were primed with the same antigen and adjuvant. At 4 to 5 weeks after the last immunization, all mice were challenged with live RSV A2 (1x106 PFU).
To test the maternal effect of antigen exposure before pregnancy, female BALB/c mice were i.n. infected twice with RSV A2 (1×105 PFU) or PBS at 2-week intervals, and mated 4 weeks after the last infection to obtain offspring. When their neonates were 1 week old, the mice were divided into two subgroups and immunized: one group was immunized i.n. with Gcf (20 μg) plus CT adjuvant (1 μg) twice at 2-week intervals and the other group (the control) was immunized with PBS alone. Five weeks after the final immunization, mice were challenged with live RSV A2 (1×106 PFU). At 4 days after challenge, the eosinophil population in bronchoalveolar lavage (BAL) fluid was analyzed by flow cytometry, and a plaque assay was performed using lung homogenates.
Analysis of cells in BAL fluid
Four days after RSV A2 challenge, mice from each group were sacrificed and their tracheas were washed twice with 700 μL of PBS. The collected BAL fluid was centrifuged and BAL cells were resuspended in 1 mL of PBS. The cells were incubated with violet fluorescent reactive dye (Invitrogen, Eugene, OR) for 10 min at room temperature and then washed with 1 mL of PBS and blocked with purified CD16/CD32 Fc (clone 2.4G2; BD Pharmingen, San Jose, CA) for 5 min. After blocking, 50 μL of antibody cocktail containing anti-CD45-APC (clone 30-F11; BD Pharmingen), CD11c-FITC (clone HL3; BD Pharmingen, San Jose, CA), and Siglec-F PE (clone E50-2440; BD Pharmingen, San Jose, CA) were added to cells and incubated at 4°C for 30 min. Cells were subsequently washed two times with PBS (2% FBS) and fixed with 200 μL of paraformaldehyde. Cells were analyzed using BD FACS LSR flow cytometer and data were analyzed with FlowJo software (version 7.2.5; Tree Star, Ashland, OR).
Lung RSV detection and cytokine analysis
After we collected mouse BAL fluid, lungs were perfused with PBS containing heparin (10 U/mL) until all blood was removed from the lungs. After perfusion, lung tissues were washed with PBS and then homogenized by passing through a 70-μm cell strainer (BD Labware, Franklin Lakes, NJ) with 2.5 mL of MEM (10% FBS). Lung homogenates were centrifuged at 300 rcf for 5 min and 100 μL of supernatants was inoculated into 90% confluent HEp-2 cells in 6-well plates. After incubation for 4 days at 37°C, each well was stained with neutral red and then incubated an additional 24 h at 37°C before PFU was determined by plaque counting. The remaining supernatant was stored at −80°C until cytokine analysis. We used the BD Cytometric Bead Assay kit (BD Biosciences, San Jose, CA) to compare cytokine levels in supernatant with mouse Th1/Th2/Th17 cytokine standards per the manufacturer's recommendations.
ELISA
RSV- or Gcf-specific antibody levels in blood sera were measured by ELISA (23). After mice were anesthetized intraperitoneally with a solution of ketamine hydrochloride (Yuhan, Seoul, Korea) and xylazine hydrochloride (Rompun®; Bayer Korea, Seoul), blood was collected from tails or orbital sinuses. 96-well ELISA plates were coated with 100 μL of RSV A2 (5×103 PFU/well) or Gcf (200 ng/well) diluted in PBS overnight at 4°C. After blocking with 5% skim milk and washing, we added serially diluted sera in blocking buffer to each well. After a wash, 1:3,000 dilution of horseradish peroxidase-conjugated goat anti-mouse IgG, IgG1, IgG2a, or IgA (Southern Biotech, Birmingham, AL) were added. Peroxidase substrate (TMB; Moss, Pasadena, MD) was added after a wash and the reaction was stopped by the addition of 0.5 N HCl. The plates were analyzed at 450 nm by ELISA reader (Molecular Devices, Sunnyvale, CA). Antibody titer was expressed as reciprocal log2 titer of serum dilution showing 0.2 of absorbance at 450 nm.
Lung histology
Five days after RSV A2 challenge, lungs were isolated and fixed with 4% formalin. Formalin-preserved lungs were embedded in paraffin, sectioned into 5-μm thicknesses, and stained with hematoxylin and eosin or periodic acid Schiff (PAS).
Statistical analysis
Data were analyzed using Prism software (version 5; GraphPad, La Jolla, CA) and expressed as mean±SEM. Statistical significance was determined by using an unpaired, two-tailed Student t test. P values less than 0.05 were considered statistically significant.
Results
Gcf vaccination in neonatal mice induces humoral immunity
A previous study reported that mucosal vaccination with either Gcf or Gcf plus CT adjuvant (Gcf-CT) could induce Ag-specific B cell immune responses in adult BALB/c mice (23). To determine whether Gcf vaccination is capable of effectively inducing humoral immune responses in neonatal mice, 7-day-old mice were immunized intranasally and boosted 14 days later (Fig. 1A). On day 35, we evaluated RSV- or Gcf-specific antibody responses using sera harvested from immunized mice. Of note, the RSV-specific serum IgG responses in the Gcf-CT immunized group were comparable to those of either the FI RSV or live RSV groups. In addition, immunization with Gcf alone also induced considerable RSV-specific serum IgG responses (Fig. 1B). In terms of IgG2a and IgG1, representing Th1 and Th2 CD4+ T cell responses, respectively, Gcf-CT–immunized mice showed dominant IgG1 responses and low levels of IgG2a responses to RSV. RSV-specific IgG2a was not detected in either the groups immunized with FI RSV or with Gcf alone (Fig. 1B). In terms of Gcf-specific antibody levels in sera, all of the isotypes in the assay (IgG1, IgG2a, and IgA) were detected in the Gcf-CT–immunized group. IgA in response to recombinant Gcf was not detected in other groups. Gcf-specific IgAs in BAL fluid, obtained 4 days after the RSV challenge, was detected in Gcf and Gcf-CT immune mice (Supplementary Fig. S1; Supplementary material is available online at
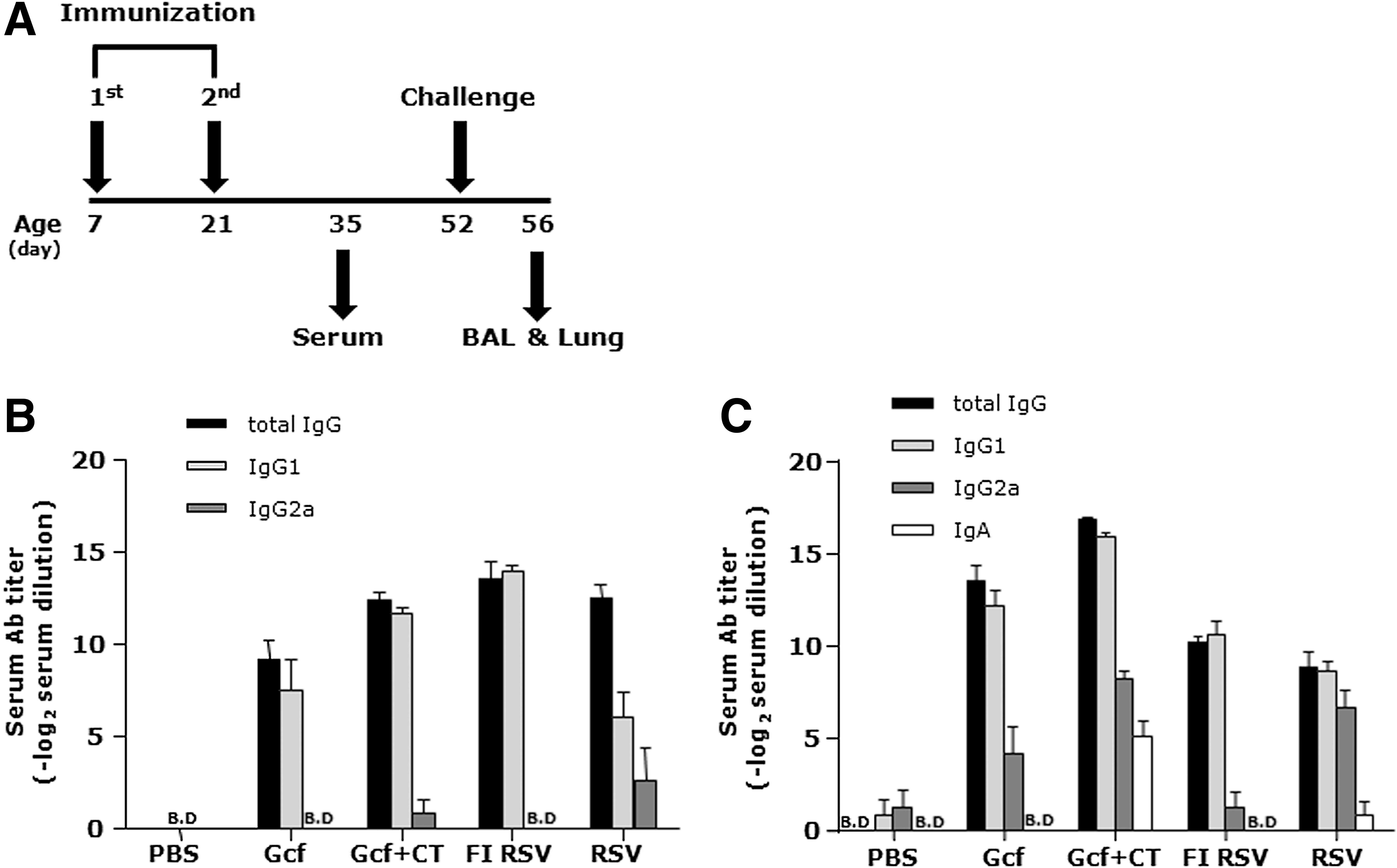
Systemic RSV and Gcf-specific antibody responses after Gcf immunization in neonatal mice.
Effect of Gcf immunization on lung eosinophilia and protection against RSV
Next, we investigated whether vaccination with Gcf promotes lung eosinophilia upon live RSV challenge, since exposure to live RSV after FI RSV vaccination can lead to airway obstruction and excessive eosinophilia (15) and, in a mouse model, live RSV infection after immunization with FI RSV resulted in Th2-skewed immune responses (45). As illustrated in Figure 1A, mice were challenged with live RSV A2 and BAL fluids were obtained after 4 days. While FI RSV-immunized mice had substantial eosinophil increases in BAL fluid, the Gcf-CT- and the live RSV-immunized groups had low levels of eosinophils (Fig. 2A). Lung histology data also showed low levels of eosinophilic infiltrations in Gcf alone, Gcf-CT, and live RSV-immune groups. Eosinophils were not detected from BAL fluid or lung tissues of PBS control group. According to our PAS staining results, bronchiolar obstruction by mucus could be detected in FI RSV immune mice (data not shown). In the contrary, mucus production could rarely be observed in the Gcf-CT group, and the levels of mucus production was comparable to that of PBS or Gcf alone group (Fig. 2D). These data demonstrate that mice immunized with Gcf or Gcf-CT may not develop vaccine-induced immunopathology after RSV infection.
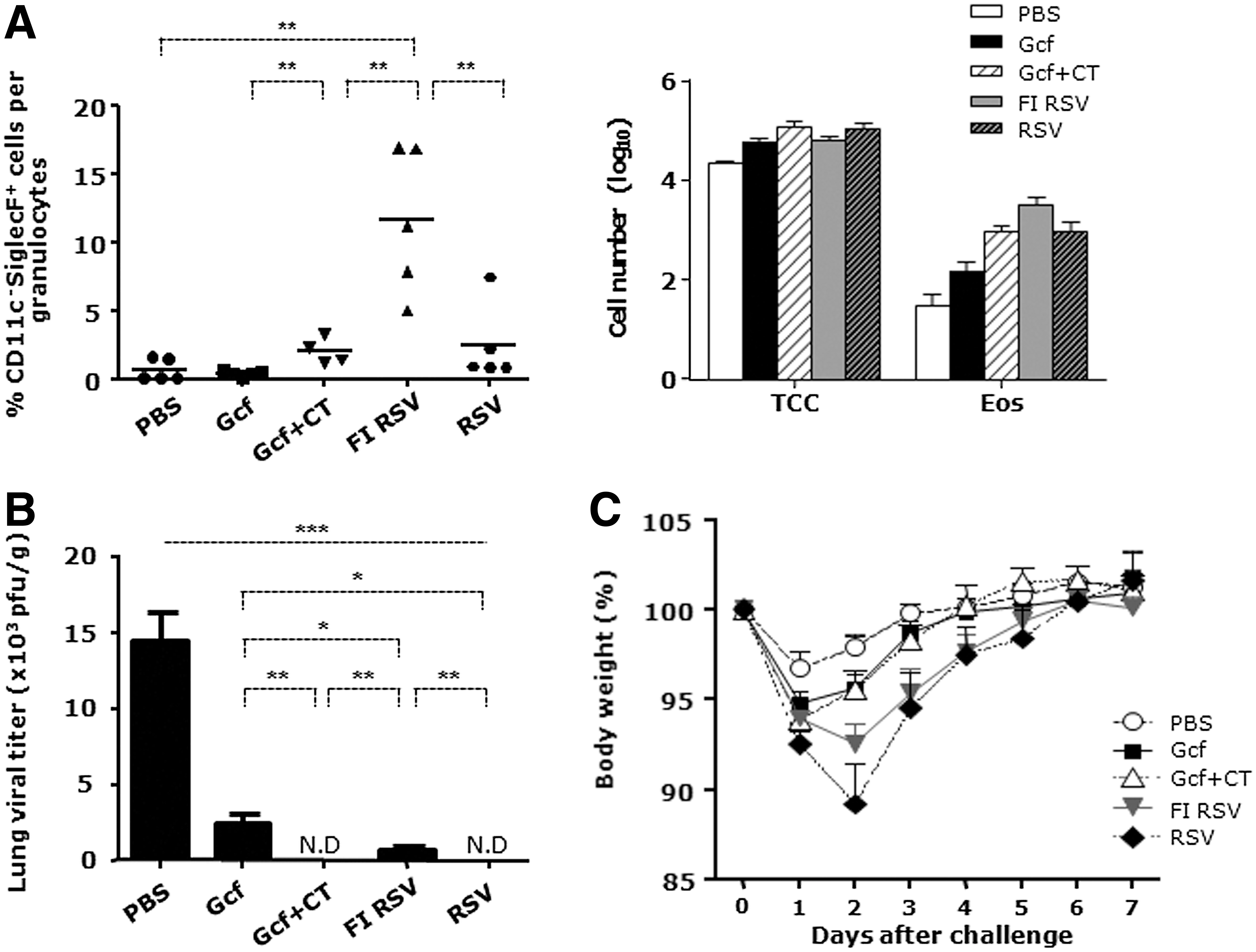
Low level of BAL eosinophilia and protective efficacy against RSV challenge following intranasal Gcf immunization of neonates.

We also analyzed whether Gcf immunization would ameliorate viral load in the lung after challenge with RSV. Lung homogenates were prepared for viral titration from lungs of mice sacrificed on day 56. At day 4 post-infection to HEp-2 cell, viral loads in lung homogenates were assessed by plaque counting. No virus was detected in lung homogenates of Gcf-CT-immune mice, whereas the PBS control group showed significant lung viral titers (Fig. 2B). In addition, Gcf alone also provided potent protection as previously reported in adult mice (23). Furthermore, Gcf or Gcf-CT-immune mice not only showed reduced weight loss upon RSV challenge but also accelerated weight recovery compared to FI RSV or RSV-immune mice (Fig. 2C). Taken together, these results indicate that i.n. immunization with Gcf greatly improves the lung viral clearance following RSV challenge without causing subsequent vaccine-mediated enhanced disease.
Gcf-CT vaccination induces immune response regardless of maternal antibody
We further examined whether pre-existing RSV-specific maternal antibodies affect the immunogenicity of Gcf in neonates. The presence of CT adjuvant has shown better protection against RSV and so Gcf alone group has not been included in the following experiments. Female mice were infected i.n. with RSV twice at 2-week intervals and mated to produce pups possessing maternally-transferred RSV-specific antibodies. The pups were immunized with either Gcf-CT or PBS at ages 1 and 3 weeks, weaned at age 3 weeks, and challenged with RSV at age 8 weeks (Fig. 3A).
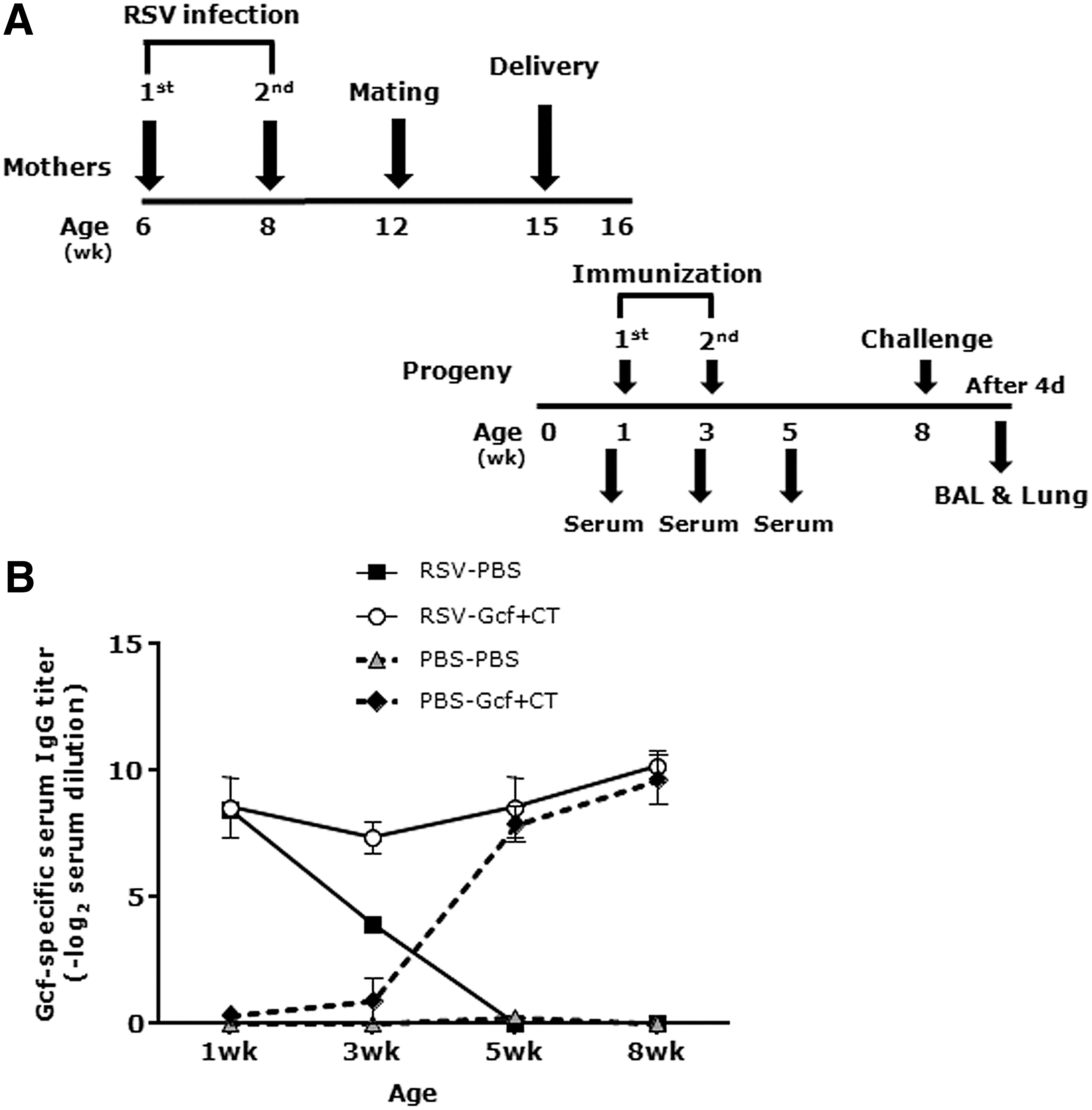
Anti-Gcf IgG changes in sera of immunized offspring of RSV-immunized mothers.
Slight increase in Gcf-specific IgG titers following the primary immunization, precipitous increase in Gcf-specific IgG titers following the booster immunizations (measured on the weaning date), and further gradual increase in Gcf-specific IgG titers until age 8 weeks were observed in Gcf-CT–immunized offspring of PBS control mothers (Fig. 3B). In Gcf-CT–immunized offspring of RSV-infected mothers, IgG titers specific to Gcf declined slightly between weeks 1 and 3 and then began to increase after booster immunization at age 3 weeks, while Gcf-specific maternal IgG titers declined steeply in PBS control offspring of RSV-infected mothers and were not detected at age 5 weeks. Taken together, these findings indicate that Gcf-CT is sufficiently immunogenic, even in the presence of pre-existing RSV-specific maternal antibodies.
Maternal antibodies have little effect on eosinophil recruitment and protective efficacy against RSV challenge
We next compared immune responses in the lung and protection efficacy in pups possessing pre-existing maternal antibodies. As described in Figure 3A, the progeny of RSV-infected and naïve mothers were immunized with Gcf-CT at ages 1 and 3 weeks and i.n. challenged with RSV A2 at age 8 weeks. BAL fluids and lungs were obtained 4 days after challenge.
The pups immunized with Gcf-CT showed significantly increased total cells in BAL fluid regardless of the presence of maternal antibodies. Likewise, although a limited number of eosinophils were detected in BAL harvested from pups that received Gcf-CT immunization, the presence of maternal antibodies against RSV A2 did not have a negative effect on the number of lung eosinophils in their RSV-challenged offspring (Fig. 4A). Next, Th2 type cytokines, represented by IL-4 and IL-6 were measured in lung supernatants by cytometric bead assay. There was no difference in IL-4 production in any group. Although the Gcf-CT–immune offspring of RSV-immunized and naïve mothers had significantly more IL-6 than non-immunized neonates, there was no statistically significant difference between these two Gcf-CT–immunized groups.
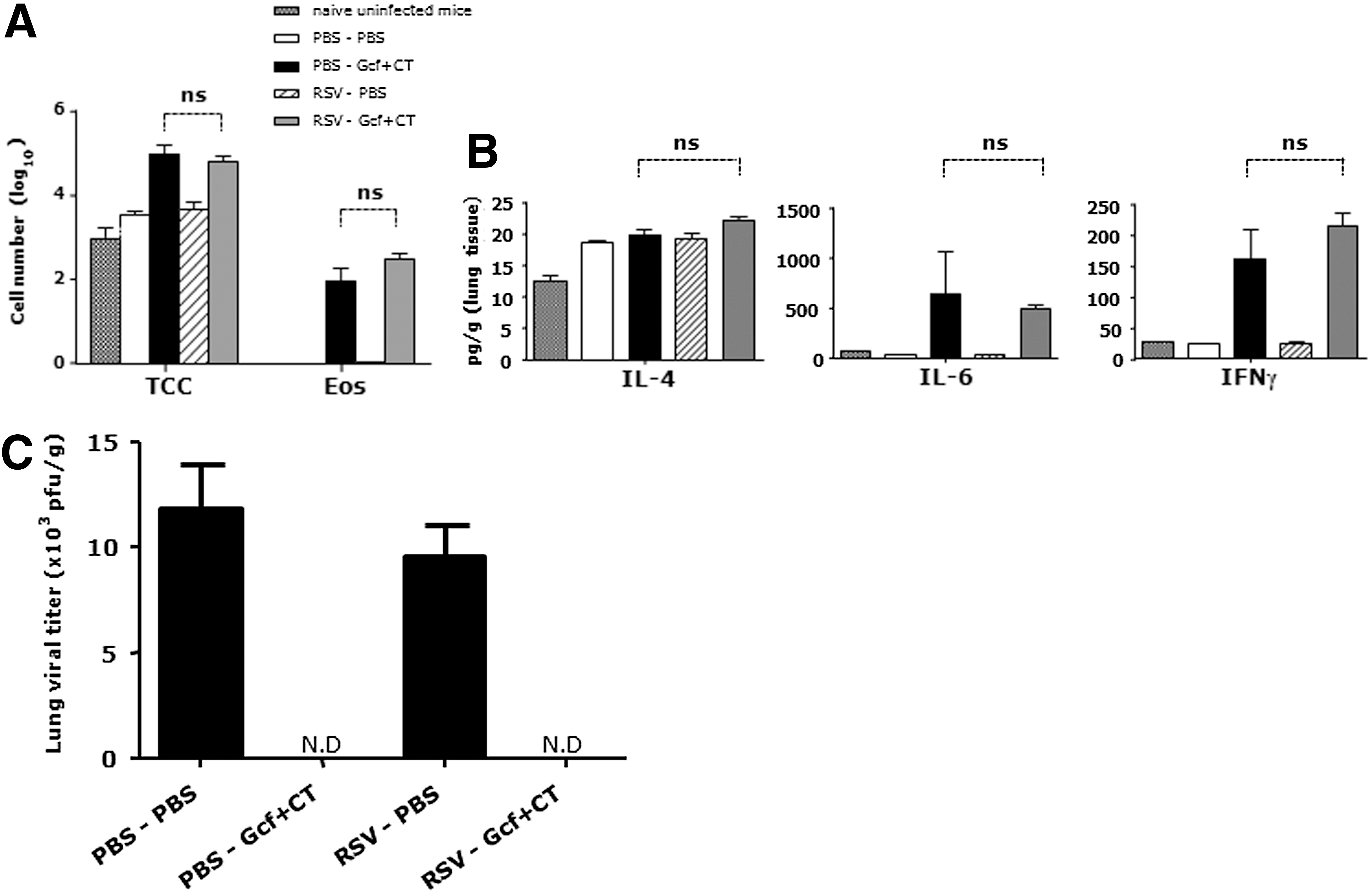
Effect of maternal immune response on neonatal Gcf immunization by lung immune response.
High levels of IFNγ were detected in the neonatal groups immunized with Gcf-CT, but there were no statistically significant differences in immunized offspring of RSV-immunized or naïve mothers (Fig. 4B). Moreover, regardless of the presence of RSV-specific maternal antibodies, Gcf-CT immunizations lead to full protection against RSV lung infections (Fig. 4C).
Collectively, these results suggest that i.n. inoculation of neonatal mice with Gcf-CT elicits protective immunity without severe vaccine-enhanced lung eosinophilia, even when total cells increase in BAL fluid, regardless of the presence of maternal anti-RSV antibodies.
Discussion
Our results clearly show that i.n. administrations of Gcf to neonatal mice induce not only prominent humoral immune responses but also protect against RSV infection without enhanced illness, regardless of the presence of passively transmitted RSV-specific maternal antibodies.
In humans, exacerbated morbidity and mortality that are related to RSV infections peak 2 to 4 months after birth. Thus, the most suitable RSV vaccine would be administered soon after birth in order to elicit effective immune responses (16,36). Although RSV vaccine development has been actively investigated for years using several animal models and numerous vaccine candidates, most studies used adult animal models (7). Because adults and neonates differ in immune system maturity, Th2 immune responses, and response to maternal antibodies (25), the neonatal animal model is useful for investigating RSV vaccine efficacy and safety. Our data indicate that most neonatal mouse immune responses are Th2-biased, as the IgG1 is predominant over the IgG2a even in response to live RSV inoculation (Fig. 1B), which is contrary to the ratio detected in adult mice (data not shown). Although antibody responses to Gcf are Th2 dominant, eosinophils infiltration into the lung after vaccination with Gcf, followed by RSV challenge is negligible compared to that after FI RSV vaccination.
In infants, vaccine-related antibody responses to rabies virus, canine parvovirus, pseudorabies virus, and feline rhinotracheitis virus are suppressed by maternally acquired antibodies (3,18,20,43,48). Several studies in adult mice suggest that passively transferred RSV antibodies suppress the immune responses against RSV F and G protein immunizations (27,28). Also, Crowe et al. (8) reported that while primary systemic and mucosal antibody responses induced by their live RSV vaccine candidate were inhibited by passive transfer of RSV-specific serum antibodies, cell-mediated immune responses were not inhibited. Because RSV reinfections occur throughout life, RSV-specific antibodies of maternal origin can play an important role for a few weeks or months in providing passive immunity to offspring (14,31,41), but unfortunately they may interrupt vaccine-associated immune responses in human infants. However, in our study, the maternal history of RSV infection of mice did not affect neonatal vaccine efficacy of Gcf with CT adjuvant. It is possible that the immunogenicity of nonglycosylated Gcf purified from E. coli differs from glycosylated G protein from either live RSV or recombinant vaccinia virus (44). If so, humoral immune responses to Gcf in offspring may not be affected by pre-existing maternal RSV-specific antibodies. Indeed, Gcf-specific immunoglobulin titer increased after booster immunization. Similar results occurred after neonatal mouse immunization with recombinant G fusion protein BBG2Na. In that study, pups developed antibody immune responses even in the presence of RSV maternal antibodies (4). Nonetheless, the clinical trial of BBG2NA was halted at phase III due to side effects. After the failure of BBG2Na as a vaccine, there have been attempts to re-evaluate the immunogenicity and protective efficacy of G2Na against RSV infection. It has been shown that two injections of G2Na with adjuvant were sufficient to induce protective immune responses in adult mice, preventing RSV infections (29).
Our Gcf (a.a. 131 to 230) lacks one amino acid compared to G2Na (a.a. 130-230). Gcf also conferred protection against RSV challenge when administered mucosally into adult mice in combination with CT. Our present study demonstrated that Gcf-CT immunization during the neonatal period was safe and efficacious in generating protective immune responses following RSV challenge, with minimal lung eosinophilia and less body weight loss compared to FI RSV immunization. In addition, the protection and Th1 cytokine productions induced by Gcf-CT immunization was not influenced by pre-existing maternal antibodies. Future studies are needed to determine more safe administration routes and adjuvant for use in humans.
Footnotes
Acknowledgment
This work was supported by grants from National Research Foundation of Korea (No. 2010-00240) and from the governments of the Republic of Korea and Sweden (SIDA).
Author Disclosure Statement
The authors have no financial conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.