
Abstract
The antibody and cellular immune responses against infectious bronchitis virus (IBV) were evaluated at mucosal sites of chickens after immunization with various doses of an attenuated vaccine at 1 day of age. The correlation of these immune responses with protection of tracheal tissues was evaluated after experimental infection of these birds. Significantly reduced tracheal pathologic effects, measured according to ciliostasis and histology lesions, and reduced viral load were observed only in the full-dose vaccinated group at 5 days post-infection (dpi), while incomplete protection was observed for the subdose vaccinated groups. Moreover, birds of vaccinated groups, especially with full dose, developed higher levels of lachrymal IBV-specific IgG and IgA and increased the expression of cell-mediated immunity (CMI) genes, such as gamma interferon (IFNγ), CD8+ T cell marker, and granzyme homolog A more rapidly. In addition, these humoral and cellular immune responses evaluated at mucosal sites correlated significantly with tracheal protection against homologous IBV challenge in a vaccine dose-dependent manner. The results indicate that IgG, IgA and CD8+ T cell responses developed at mucosal sites after IBV vaccination of day-old chicks, could be taken as good correlates of protection against this virus.
Introduction
A number of studies have reported the development of humoral immune responses after vaccination against IBV, but most of these studies were limited to the measurement of serum antibodies by ELISA, virus neutralization (VN), or inhibition of hemagglutination (HI). However, the systemic anti-IBV antibody responses have shown low correlation with protection against IBV infection (3,4), though IgA anti-IBV antibodies in mucosal secretion have been found to exert a relevant role in the protection against re-infection with this virus. A marked increase in the expression of Ig γ chain gene was found in the trachea of chickens secondarily immunized with IBV, suggesting that the IgG locally produced after secondary immunization provided effective protection against IBV by neutralizing this virus (10). In addition, the few studies that analyzed the levels of anti-IBV antibodies in the respiratory mucosal surface, including the lachrymal and tracheo-bronchial secretions, did not find a significant correlation, nor clearly determine whether a correlation exists between the levels of local antibodies and the protection status against IBV (3,22,24).
A close association was also reported between the development of systemic CMI and viral clearance and the reduction of clinical signs and tissue lesions in chickens experimentally challenged with IBV (5). Additionally, the transfer of memory cytotoxic CD8+ T cells (CTLs) from the spleens of immune birds convalescing from IBV infection to naive inbred chicks provided the recipients protection against homologous viral infection (5).
Another study assessed the immunophenotyping of mononuclear cell infiltrating the tracheal mucosa of naive chickens experimentally infected with IBV. The CD8+ T cells were found significantly increased in the trachea after 3 or 4 days post-infection (dpi), and peaked at 5 dpi. These cells decreased to baseline levels by 14 dpi. In view of this, the infiltrating CD8+ T cells at trachea mucosa were supposed to be involved in the clearance of IBV at this site, in an early phase of infection (14). However, no immunophenotyping was addressed in birds previously vaccinated against IBV and challenged.
During the course of viral infections, a number of mediators produced by CD8+ effector T cells (CTLs), such as IFNγ, granzymes, and perforins, have been associated to the destruction of virally infected cells (9,13,16,18). Additionally, the transcriptome analysis have been developed to measure the expression of genes associated with immune responses at tracheal sites after infection and/or re-infection with attenuated Massachusetts IBV strains (10,24).
Despite all these investigations, the effector immune mechanisms induced by IBV vaccines and which are effectively involved to confer protection against this virus, remain to be better understood (4).
Thus, the goal of this study was to measure the tracheal CMI and lachrymal antibody responses induced by different doses of attenuated IBV vaccine administered to day-old chicks, and to estimate correlation between these immune responses with tracheal protection against IBV infection with virulent IBV strain, and evaluated in this study as ciliostasis, histopathology, and viral load in the tracheal samples of experimentally infected chickens.
Materials and Methods
Virus
The IBV M41 strain was obtained from Empresa Brasileira de Pesquisa Agropecuária–Embrapa (Concórdia, SC, Brazil). This virus was propagated and titrated in 10-day-old specific pathogen-free (SPF) embryonated chicken eggs. The eggs were inoculated via the allantoic sac route, as described previously (17). The commercial H120 attenuated live strain was used for the immunization.
Both the M41 and vaccine strains were titrated in embryonated chicken eggs according to standard procedures (6), and the 50% embryo-infective doses (EID50) were determined (19).
Experimental design
Five groups of 14 SPF (specific pathogen-free) chicks (White Leghorn lineage) were used. The birds were housed in positive-pressure isolators. Groups I, II, and III received, respectively, one full dose (104.3EID50/bird, as recommended by the manufacturer), half dose (103.99EID50/bird), or quarter dose (103.68EID50/bird) of the attenuated H120 vaccine strain of IBV at the first day of age via the ocular-nasal route. Groups IV and V received only the vaccine diluent. At 28 days of age, groups I, II, III, and IV were experimentally infected via the intra-ocular and intranasal routes with the M41 strain of IBV (104.0 EID50/bird). A negative control group (Group V) was mock infected with SPF allantoic fluid and maintained under the same conditions. Samples of tears were collected at 1, 2, 3, and 4 weeks post-vaccination (pv) and at 24 hours (h) and 5 days (d) post-infection (pi) and stored at −20°C. Seven chickens from each group per interval were sacrificed at 24 hpi and 5 dpi. Tracheal samples were collected from each group; a portion was immediately frozen and kept at −70°C until processing, and the remaining portion was subjected to histopathological and ciliostasis analysis.
Measurement of local antibodies against IBV
The levels of local IgG, IgA, and IgM were measured from tear samples using the sandwich-ELISA-concanavalin A (S-ELISA-ConA) technique, as described (2), with the exception that anti-chicken IgG, IgA, and IgM peroxidase conjugates (Bethyl Laboratories, Montgomery, TX) were used.
The tear samples were diluted in PBS (1:25) and the reagents used for each well were 2.5 μg of Concanavalin A, viral antigen (50 μL of LCA H120 strain), peroxidase conjugates IgA (1:4000), IgM (1:800), and IgG (1:6250). All procedures, including wash steps, temperatures, time of incubation, and determination of S/P values, were performed as reported previously (2).
Scoring of tracheal lesions
The tracheal samples were technically processed for ciliary kinetic analysis and histopathology, and the scores were determined according to recommendations described previously (1,7,26).
RNA extraction and reverse transcription
The RNA extractions from the tracheal samples of experimentally infected chickens were performed using the Trizol reagent (Invitrogen). The extracted RNA was resuspended in 10 μL of RNAse-free water treated with diethylpyrocarbonate (DEPC). The residual genomic DNA was digested by DNase treatment of RNA samples using a RNase-free DNase I kit (Fermentas). The RNA quality was analyzed on a 1% formaldehyde gel and quantified by ultraviolet (UV) absorbance at 260 nm (A260). The cDNAs were synthesised according to instructions provided with Reverse Transcriptase (Fermentas) and using OligodT (IDT), with the exception that dithiothreitol (DTT) was omitted from the reaction, accordingly to recommendations (23).
SYBR Green I real-time PCR
Real-time PCR using the SYBR Green I marker was used for the relative and absolute quantification of mRNA and viral load, respectively. The reaction mixture contained 2.0 μL of cDNA, 10.0 μL of Maxima SYBR Green qPCR Master Mix (Fermentas), and 10 pmol of each primer (Table 1) in a final volume of 20 μL. The amplification reaction included a pre-incubation step at 95°C for 10 min to activate the HotStart Taq DNA polymerase. This reaction was followed by 40 cycles of amplification (30 cycles for the 18S), including denaturation at 95°C for 30 sec, annealing at 49.2°C to 65°C (Table 1) for 30 sec, and extension at 72°C for sec. Following amplification, a melting curve analysis was performed by raising the incubation temperature from 65°C to 95°C in 0.2°C increments with a hold of 1 sec at each increment. The oligonucleotides used were designed according to sequences deposited in GenBank and using Primer3 software (
a, 5′to 3′sense; Bp, base pairs; F, forward; R, reverse.
The relative expression of the CD8+, Granzyme A, and IFNγ genes in the tracheal samples was quantified as the fold change between samples relative to the nonvaccinated and noninfected group (Group V) at the same time point, and the gene expression from each sample were standardized using the Ct value of the 18S rRNA for the same sample, according to the recommendations (15).
Standard curve construction and absolute quantification of viral replication
Total RNA was extracted from the allantoic fluid of eggs infected with the M41 strain using the Trizol LS reagent (Invitrogen) and was then submitted to RT using a SuperScript kit (Invitrogen) and random primers (Invitrogen). The cDNA obtained was processed by PCR for the amplification of the whole open reading frame of the S1 gene (approximately 1620 base pairs) using a High Fidelity kit (Invitrogen) and the forward (5′-CACGTGATGTTGGTAACACCTCTTTT-3′) and reverse (5′-CCGCGGAGAACGTCTAAAACGACG-3′) oligonucleotides. The amplified product was purified using the GFX PCR DNA and Gel Band Purification kit (Amersham) and cloned using the pGEM T Easy vector (Promega) according to the manufacturer's recommendations. The recombinant plasmid was used for the transformation of Escherichia coli (DH10b lineage). The plasmid DNA was extracted using the Wizard Plus SV Minipreps DNA Purification System (Promega), quantified and converted to the number of moles using the
All tracheal samples were tested for viral replication using the HV+ and HV- primers, and the CT results were used to calculate the Log of the number of moles using the linear equation.
Statistical analysis
The comparisons of the mean antibody levels, CMI gene expression, and tracheal pathological changes (viral replication, histopathology, and ciliostasis) between the experimental groups were performed using the Kruskal-Wallis test, followed by Dunn's test. Correlations between the immune response parameters and the tracheal pathology parameters were estimated using the Spearman method. All analyses were conducted using the Graph Pad software, and the probability level for significance was set as p≤0.05 (11).
Results
Pathological alterations of the trachea induced by IBV challenge
Tracheal ciliostasis and histopathology induced by IBV challenge in immune and non-immune birds
None of the experimental groups exhibited significant alterations of ciliostasis at 24 hpi, although the histopathology scores increased slightly, from score 2 to 9, in the vaccinated groups at this time point. The most striking pathological changes occurred in the nonvaccinated birds (Group IV) at 5 dpi, which showed marked ciliostasis and the histological lesions in the trachea (Fig. 1A, B). In contrast, the birds from all of the vaccinated groups showed less tracheal damage. The mean of tracheal lesion scores of the vaccinated groups were significantly different (p≤0.05) from those of the control group. A vaccine dose-dependent protection against pathological changes in the trachea was observed in the vaccinated groups. The lower mean scores of tracheal lesions were recorded for the full-dose vaccinated birds, and they were significantly (p≤0.05) from the half- and quarter-dose vaccinated groups (Fig. 1A, B).
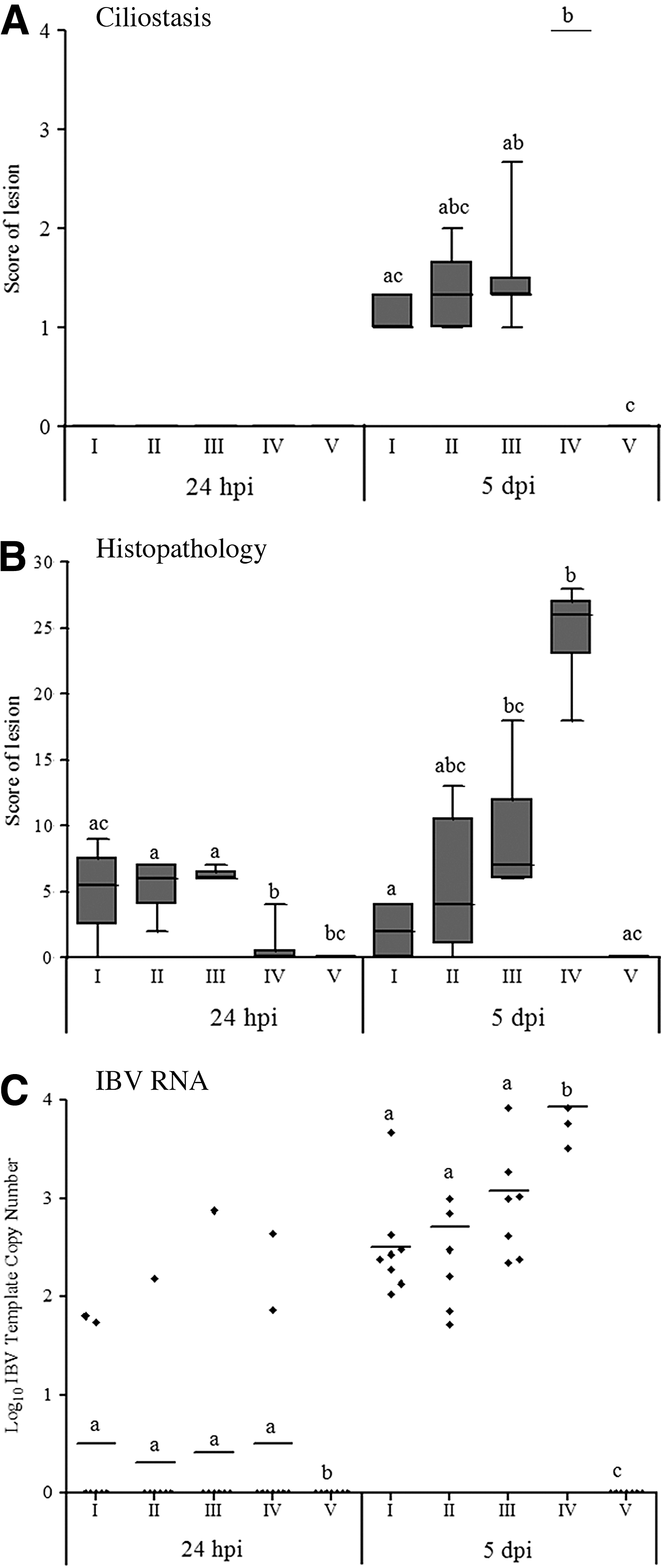
Pathological alterations of the trachea induced by IBV challenge in samples collected at 24 hours and 5 days post-infection from chickens experimentally challenged at 28 days of age with the M41 strain of infectious bronchitis virus. The chickens were previously immunized with different doses of the H120 strain at one day of age (I=one full dose, II=half dose, and III=quarter dose) or were left non-immunized (IV). Group V was mock vaccinated and infected. Significant differences between groups for each interval were detected by the Kruskal-Walls test, followed by Dunn's mean test and are represented by different letters, with p≤0.05.
Quantification of viral load in the tracheas of challenged birds
At 24 hpi, the means of number of copies of S1 gene were below the detection limit in the groups infected with IBV. At 5 dpi, however, all of the tracheal samples collected from the vaccinated and nonvaccinated groups infected with IBV were positive for S1 gene, although the viral load was significantly higher (p≤0.05) (≅ 104 copies of the S1 gene) in group IV (nonvaccinated and infected) than in the other vaccinated groups, as measured by real-time RT-PCR. The vaccinated groups showed mean of number of copies of S1 gene varying from 102 (the limit of detection for real time RT-PCR) to 103, and there were no significant differences between these groups (Fig. 1C). Additionally, significant positive correlations were observed between viral load and the tracheal histopathology, and also with ciliostasis scores (Table 2).
a, Relative quantification by Real Time RT-PCR; b, Measurement by ELISA; c, Absolute Quantification by Real Time RT-PCR; d, Interval post-vaccination. *** P≤0.001; ** P≤0.005; * P≤0.01; N, nonsignificant.
Measurement of lachrymal antibodies against IBV
Gradual and slight increases of anti-IBV IgA, IgM, and IgG levels were observed in tear samples from 1 to 4 weeks after vaccination in day-old chicks. The levels of all antibody isotypes increased further after challenge in the vaccinated groups (Fig. 2).
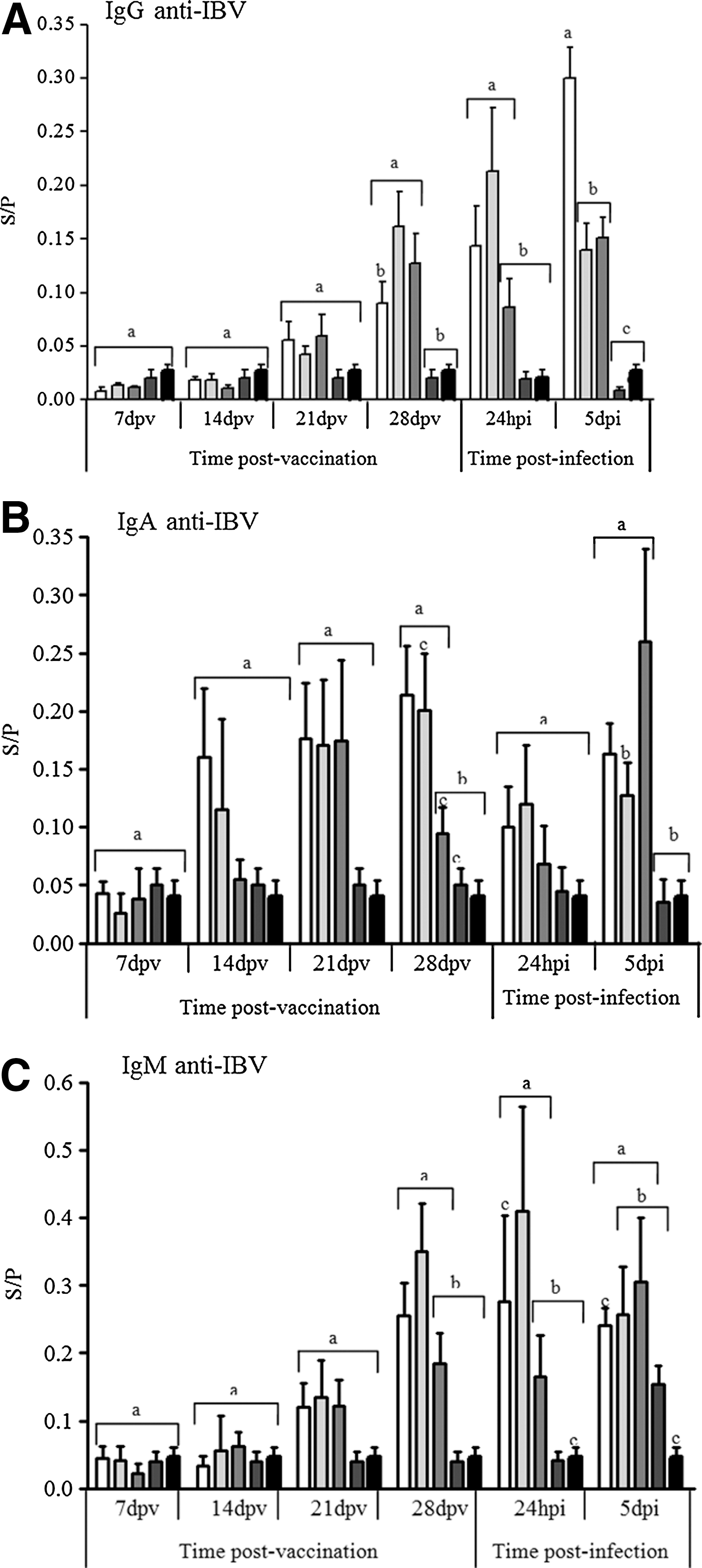
Mean Sample/Positive (S/P) values of optical density (OD) for the levels of anti-IBV IgG
Only the full-dose vaccinated group (Group I) demonstrated anti-IBV IgA levels that were significantly different from those of the unvaccinated group at challenge day (28 dpv) (Fig. 2B). In addition, the local IgG levels peaked at 5 dpi, and the levels of this isotype were also significantly higher (p≤0.05) in the full-dose vaccinated group (I) than in the remaining groups, including those that received vaccine subdoses (groups II and III) (Fig. 2A). The levels of IgM in the lachrymal secretions were not significantly different between the vaccinated groups (Fig. 2C).
The levels of tear IgG showed the strongest negative correlation (p≤0.05) with the three parameters of tracheal pathological alterations induced by IBV at 5 dpi (ciliostasis, histopathology, and viral replication). Similar negative correlations were also found for IgA, but with lower correlation coefficient (Table 2).
Expression of CMI genes in tracheal tissues after IBV challenge in immune and non-immune birds
At 24 hpi, only the full-dose vaccinated group demonstrated moderate and significant increases in CD8 and IFNγ transcripts compared with the unvaccinated and infected group (group IV). Moreover, there was granzyme-A upregulation in all of the vaccinated groups compared with the nonvaccinated chickens from group IV, and the transcript levels of this gene were higher in the full-dose vaccinated group (p≤0.05). In contrast, CD8 gene expression was downregulated at 5 dpi in the vaccinated groups, resulting in lower expression levels, while there was a significant (p≤0.05) upregulation of this gene in the nonvaccinated group compared with the full- and half-dose vaccinated groups. At this time point, the nonvaccinated infected birds (group IV) showed a significant increase (p≤0.05) in IFNγ transcripts only in comparison to the full-dose vaccinated birds (group I), whereas the granzyme-A gene expression of these birds was increased significantly (p≤0.05) compared with both the full- and half-dose (group II) vaccinated birds (Fig. 3).
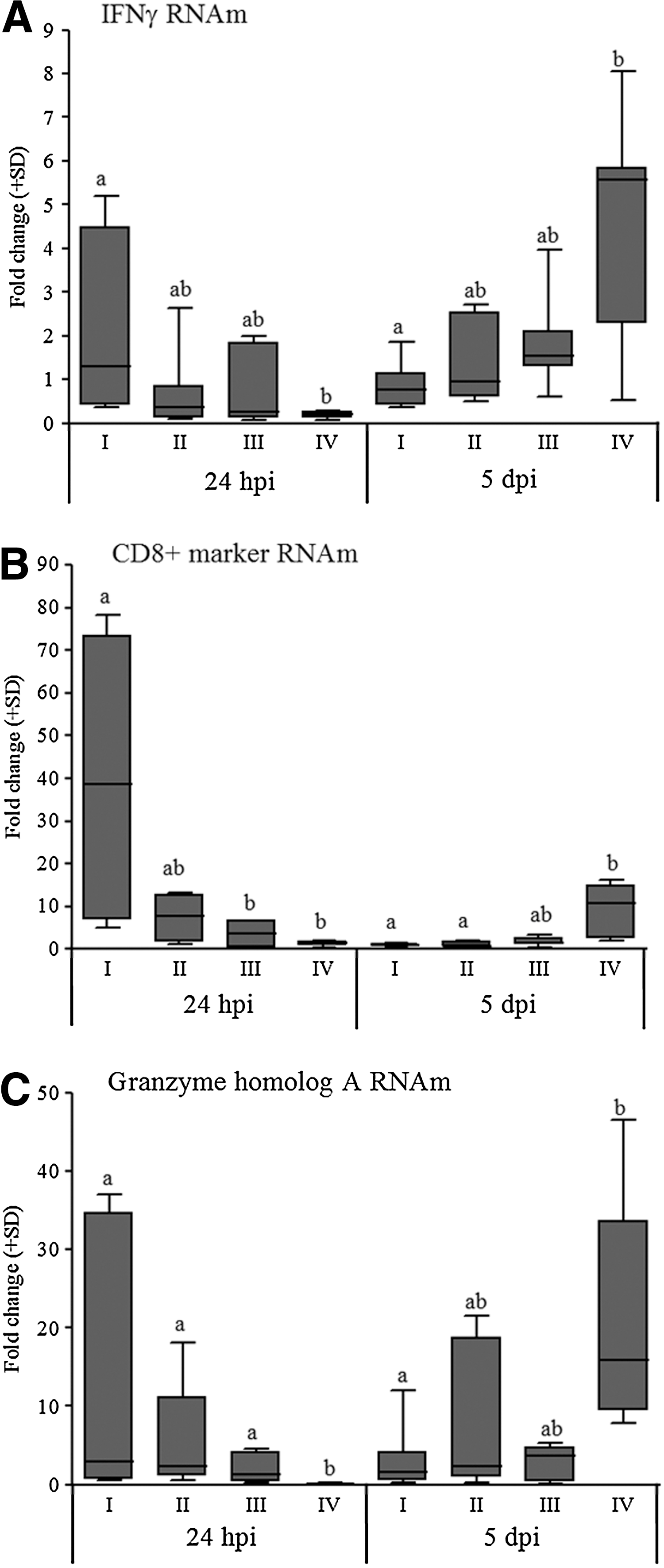
Mean fold changes in the mRNA expression of the IFNγ
The levels of granzyme-A gene expression at 24 hpi demonstrated a negative and significant correlation (p≤0.05) with the three parameters of tracheal pathologic changes induced by IBV experimental infection (viral load, histopathology and ciliostasis) at 5 dpi (Table 2). The levels of expression of IFNγ and CD8 genes demonstrated negative correlations (p≤0.05) only with the histopathology scores (Table 2).
Discussion
The results of this study showed that only the full-dose vaccinated birds were effectively protected against experimental IBV infection and that the immune mechanisms induced in day-old chicks by the attenuated vaccines against IBV depend on the development of local antibodies and CMI responses. Thus, only the full-dose vaccinated group maintained a high integrity of ciliary movement at 5 dpi, developed less tracheal histopathology, and had lower viral loads, whereas the subdose vaccinated groups displayed higher levels of tracheal pathological changes according to at least one of these three parameters. However, at 24 hpi, all of the vaccinated groups demonstrated increased tracheal histopathology scores, due the presence of intense infiltrates of mononuclear cells, when compared with the unvaccinated group. This inflammatory response was likely due to the transient and early migration of mononuclear cells to the primary site of IBV replication, as a possible consequence of participation of chemokines derived from T cell mediated-immune responses. This response was previously described for tissues from respiratory tract infected with IBV, or avian influenza virus (14,10,20).
The local memory antibody response was induced by the administration of an attenuated IBV vaccine to day-old chicks, as shown by the increase in all anti-IBV isotypes at 24 hpi and 5 dpi. In addition, the production of IBV-specific IgG in the lachrymal secretions at 5 dpi was dose dependent of the attenuated vaccine administered to the day-old chicks. At 5 dpi, the full-dose vaccinated group showed significant increases in the anti-IBV IgG levels in tear samples compared with the groups receiving vaccine subdoses. Conversely, no significant differences were detected in the anti-IBV IgA and IgM levels in the lachrymal secretions between the vaccinated groups at this time point. Furthermore, the early increase in the anti-IBV IgG levels detected at 24 hpi and 5 dpi in this study agree with the upregulation in the expression of IgG gene detected in a transcriptome analysis of tracheal samples collected from birds re-exposed to an attenuated vaccine strain of IBV and re-exposed after 21 days to the same viral strain (10). Thus, this increase in lachrymal anti-IBV IgG corroborates the hypothesis of the former authors (10) regarding the important role played by this antibody isotype against IBV infection of upper respiratory tract.
Additionally, moderate to high negative correlations were observed between anti-IBV IgG and IgA levels in lachrymal secretion, at 24 hpi, and the tracheal pathological effects caused by this virus, at 5 dpi, evaluated here as scores of ciliostasis, histopathology, and viral load. This supports the idea to take these lachrymal antibodies as good correlates of protection, or indicative of effective immunization, induced by attenuated IBV vaccines.
The expression levels of the CD8, granzyme-A, and IFNγ genes in tracheal mucosa of the full-dose vaccinated birds were significantly increased at 24 hpi (Group I), demonstrating the development of a CMI memory response. In addition, the vaccine doses affected the expression of the IFNγ and CD8 genes, as the expression levels of these genes were significantly higher than those of the non-immune birds only in the full-dose vaccinated group. The granzyme A gene was also highly expressed by all vaccinated groups at 24 hpi, and the levels of expression of these groups differed from those of the non-immune birds.
The important role of CMI induced by IBV vaccination of day-old chicks in the tracheal protection against this virus was indicated in this study by the consistent negative correlations found between the expression of CMI-genes at 24 hpi and tracheal pathologic changes at 5 dpi. The expression of granzyme-A gene was negatively correlated with all the tracheal pathological parameters evaluated, which make this gene a good correlate of immune-protection against IBV infection of respiratory tract.
This early upregulation of mucosal CMI-related genes in vaccinated birds nearly coincided with the kinetics of systemic CTL activation and the memory responses previously reported following infection with the IBV Gray strain (5). IFNγ and granzyme-A are produced mainly by CD4 and CD8 T lymphocytes, although these CMI mediators can also be produced by other cells of the immune system, especially NK cells (12). However, the observed kinetic profile of the CMI-related genes detected here in the IBV-vaccinated birds more closely resembled the evolution of T cell activation against viral infection, which is followed by extensive transcriptional and functional changes in CD8 memory T cells and the acquisition of CTL activity (5,21,25). Similar early increases in CMI-related gene expression were also reported in lung samples collected from chickens that had been or not vaccinated with an inactivated oil-adjuvanted vaccine and infected with avian influenza virus (8).
In contrast to former findings (10), an upregulated expression of CMI-genes was detected in the present study. This discrepancy may be due to the fact that in this previous study the expression of CMI-genes was measured after a secondary administration of attenuated IBV vaccine strain (H120), whereas in the current study the expression of these genes was measured after challenging vaccinated chickens with a virulent IBV strain (M41). It is possible to hypothesize that the virulent IBV strain was not totally blocked by innate immune mechanisms and mucosal anti-IBV antibodies, and after infecting susceptible cells from the host respiratory system, the viral antigens were recognized by memory T cells, activating CMI secondary responses, which were detected here as an early and up-regulated expression of CD8, IFN-γ, and granzyme A genes in the tracheal samples of vaccinated birds.
Additionally, it appears that in non-immune birds, the late increase in the expression of CMI genes, at 5 dpi, may contribute to pathological tissue changes, as indicated by the fact that the increased expression of CD8, granzyme A and IFN-γ at 5 dpi correlated with viral replication in the trachea. There was also a positive correlation between IFN-γ gene expression at this time point and high tracheal histopathology scores, which were likely caused by delayed and unbalanced immune responses in nonvaccinated birds, as detected for this and other inflammatory cytokine genes in the experimental infection of non-immune birds with avian influenza virus (20). However, these results should be interpreted with caution and further investigations are necessary to advance our understanding of the mechanisms involved in the pathogenesis of IBV infection.
A conceivable constraint of our study is related to the fact that it evaluated the expression of CMI genes in tracheal tissues, but it did not measure directly the activity of CMI, such as the cytotoxic mechanisms of CTLs. Despite this, the upregulated expression of CMI genes such as perforin, granzymes, and IFN-γ has been associated with activation markers of CD8 T cells derived from tissues infected with influenza virus or infectious bursal disease virus (13,16,18). Thus, these findings support our approach for using the expression of these CMI-genes as markers of effector activity of CD8 T cells induced by IBV vaccination in tracheal tissues.
In conclusion, our results indicate that measuring anti-IBV IgG and IgA antibodies in the lachrymal secretion and the expression of granzyme-A, CD8, or IFN-γ genes in the tracheal tissues after IBV infection provides a reliable approach to evaluate the immune protection status induced by vaccination with this virus. Furthermore, the intensity of these humoral and cellular immune responses and their ability to confer effective protection were dependent on the dose of vaccine administered.
Footnotes
Acknowledgments
We would like to thank: financial support of FAPESP (07/55070-2) and CNPq (476120/2007-1); Dr. Gener Tadeu Pereira from UNESP—Jaboticabal for contribution with statistical analysis; Dra Liana Brentano from Embrapa Swine and Poultry for providing the virus; Pfizer and Merial for supplied SPF eggs and SPF chicks.
Author Disclosure Statement
No competing financial interests exist.