
Abstract
Th17 cells and the secreting cytokines play an important role in the immune response and inflammation that is induced by hepatitis B virus (HBV). However, it remains not fully elucidated how the antiviral agents affect Th17 cytokines and signal pathway. Telbivudine therapy has been proved to inhibit HBV replication effectively and to improve clinical outcome of chronic hepatitis B (CHB). Thus, in this study, the effect of decrease in viral load and liver dysfunction resulting from telbivudine treatment on Th17 cells and the related cytokines IL-17, IL-22, and IL-23 were analyzed. Peripheral blood mononuclear cells and serum from twenty-four CHB patients were harvested at 0, 12, 24, 36, and 48 weeks after initiation of telbivudine treatment. In parallel to the reduction of HBV DNA and normalization of serum ALT, significant declines in circulating HBV-specific Th17 cells and IL-22 production were found during antiviral therapy. The expression of serum IL-22 and IL-23, but not IL-17 also decreased during therapy. Our findings suggest that antiviral effect of telbivudine may attribute to both direct virus inhibition and regulation of inflammation, which further improve the understanding of pathogenesis of HBV infection and develop antiviral strategy for controlling viral hepatitis.
Introduction
In recent years, a separate T cell lineage, called Th17 cells, producing interleukin (IL)-17 family cytokines (IL-17A and F) (39), has been identified (10,32). Th17 cells represent a new T helper lineage that requires transforming growth factor-β (TGF-β) and IL-6 for initiation (2) and IL-23 to become an established population (16). IL-17 is believed to contribute to the pathogenesis of autoimmune and inflammatory diseases by acting as a potent proinflammatory mediator (19). IL-17 is also important in the host defense against extracellular bacteria such as Klebsiella pneumoniae (9), Bacteroides fragilis (6), or Mycobacterium tuberculosis (17), against virus such as hepatitis C virus (HCV) (35) and human immunodeficiency virus (HIV) (29), and against fungi such as Candida albicans (14). Th17 cells also expressed IL-22, which acted cooperatively with IL-17 to enhance expression of antimicrobial peptides associated with host defense (24). IL-22 not only induces the expression of antimicrobial proteins that are needed for defense against pathogens in the skin (40), but also display pro-inflammatory functions in several diseases such as dermal inflammation (46), psoriasis (41), and Crohn's disease (3).
Th17 cells and the secreting cytokines, IL-17 and IL-22, have been demonstrated to be closely correlated with chronic hepatitis B (CHB) (44,45). The high viral load present in peripheral blood of HBV patients could possibly provide a stimulus to sustain the Th17 function. To gain further understating of Th17 cells and the related cytokines in chronic HBV infection, we investigated longitudinally the relationship between circulating Th17 cells, IL-17/22, and viral load in chronic hepatitis B patients who underwent telbivudine therapy.
Patients, Materials and Methods
Patients
Twenty-four CHB patients (21 males and 3 females, average age 33.29±8.22 years, HBV DNA 7.035±1.176 log10copies/mL, alanine aminotransferase [ALT] 146.75±63.46 U/L) were enrolled in this study. All patients were HBV e antigen (HBeAg) positive, and hospitalized or followed-up in the Department of Infectious Diseases, Tangdu Hospital, Xi'an, China, between March 2010 and February 2012. Diagnoses of chronic hepatitis B were made in accordance with the standard of Chinese National Program for Prevention and Treatment of Viral Hepatitis. No enrolled patients were co-infected with HIV, HCV, or other hepatitis viruses. Moreover, patients who received antiviral or immunomodulatory treatments within 1 year of baseline sampling were also excluded from the study. All patients took 600 mg of telbivudine (Novartis, Beijing, China) orally once daily with no restriction of food intake. Blood samplings were made on five different time points from all patients: baseline, 12 weeks, 24 weeks, 36 weeks, and 48 weeks after initiation of telbivudine therapy.
Peripheral blood mononuclear cells (PBMCs) isolation and stimulation
PBMCs were isolated by Ficoll-Hypaque solution (Sigma-Aldrich, St. Louis, MO, USA) using density gradient centrifugation methods from 10 mL of fresh EDTA anticoagulant bloods, and were immediately cryopreserved in a medium containing 90% heat-inactivated fetal bovine serum (FBS) and 10% dimethylsulfoxide (DMSO) after isolation. The PBMCs were thawed 1 day before the experiments, and cell viability was over 85% by trypan blue exclusion. PBMCs were stimulated with PMA (50 ng/mL) and ionomycin (1 μg/mL), with Brefeldin A (BFA, 10 μg/mL) for 5 hours for the measurement of nonspecific Th17 cells. Otherwise, cells were incubated with HBV envelope peptide FLLTRILTI (1 μg/mL) and core peptide FLPSDFFPSV (1 μg/mL), with BFA (10 μg/mL) for 12 hours for detection of HBV-specific Th17 cells. In some experiments, PBMCs were stimulated with recombinant HBV surface antigen (HBsAg, AbD Serotec, Oxford, UK, 1 μg/mL) in 96-well plates for 4 days, and supernatants were harvested for further analyses.
Flow cytometry
The stimulated PBMCs were transferred into FACS tubes, and anti-CD3-PerCP (BD Bioscience, San Jose, CA, USA), anti-CD4-APC (BD Bioscience), and anti-CD8-PE (BD Bioscience) were added and incubated at 4°C in the dark for 30 min for surface staining. Cells were then fixed by adding 100 μL of Fixation and Permeabilization Medium A (Caltag Laboratories, Invitrogen, Carlsbad, CA, USA) for 15 min incubation in the dark at room temperature. PBMCs were resuspended in 100 μL of Fixation and Permeabilization Medium B (Caltag Laboratories) containing anti-IL-17A-FITC (eBioscience, San Diego, CA, USA) for 20 min incubation. Isotype control antibodies were used to separate positive and negative cells in the PerCP, APC, FITC, and PE fluorescence channels. Samples were analyzed with a four-color FACS Calibur analyzer (BD Biosciences Immunocytometry Systems, San Jose, CA, USA). Acquisitions and analyses were performed with CellQuest Pro software (BD Biosciences Immunocytometry Systems), and data were analyzed by FlowJo version 5.7.2 for Windows (Tree Star Inc., Ashland, OR, USA).
Enzyme-linked immunosorbent assay (ELISA)
Concentrations of IL-17, IL-22, and IL-23 were measured using a commercial ELISA kit (eBioscience) according to the manufacture's instruction.
Virological and biochemical assessments
HBsAg, HBeAg and anti-HBe were quantified using ARCHITECH HBsAg, HBeAg and anti-HBe Reagent kit (Abbott GmbH and Co. KG, Wiesbaden, Germany), respectively. HBV DNA was quantified by real-time polymerase chain reaction (PCR) kit (Da'an Gene Co Ltd, Guangzhou, China) with detection limit of 3log10 copies/mL. Serum biochemical assessments (including albumin, ALT, bilirubin, blood urea nitrogen, creatine, and creatine kinase) were measured by an automatic analyzer (Hitachi 7170A, Hitachi Ltd, Tokyo, Japan).
Statistical analyses
Data were analyzed using SPSS version 12.0 for Windows (SPSS, Chicago, IL, USA). Wilcoxon matched pairs test were used to the comparison between groups. Pearson correlation tests were performed for correlation analysis. A value of p<0.05 was considered to indicate a significant difference.
Results
Reduction of HBV DNA and normalization of ALT during telbivudine therapy
During 48 weeks of telbivudine therapy, the HBV DNA of all enrolled patients revealed an immediate and significant decline to a low or undetectable level 12 weeks after initiation of telbivudine treatment. The average decrease of HBV DNA is 3.28 log10copies/mL 12 weeks of therapy, and the inhibition of viral replication was maintained until the end of observation (Fig. 1A). However, there were four patients who did not obtain virological response with detectable viral load in the serum after 48 weeks therapy. Seven patients achieved HBeAg/anti-HBe seroconversion 48 weeks after treatment, but HBsAg loss was not found in any of the patients. Furthermore, 48 weeks of treatment led to normalization of ALT in 18 of patients (Fig. 1B), and 17 of patients reached both virological and biochemical response at 48 weeks of treatment.
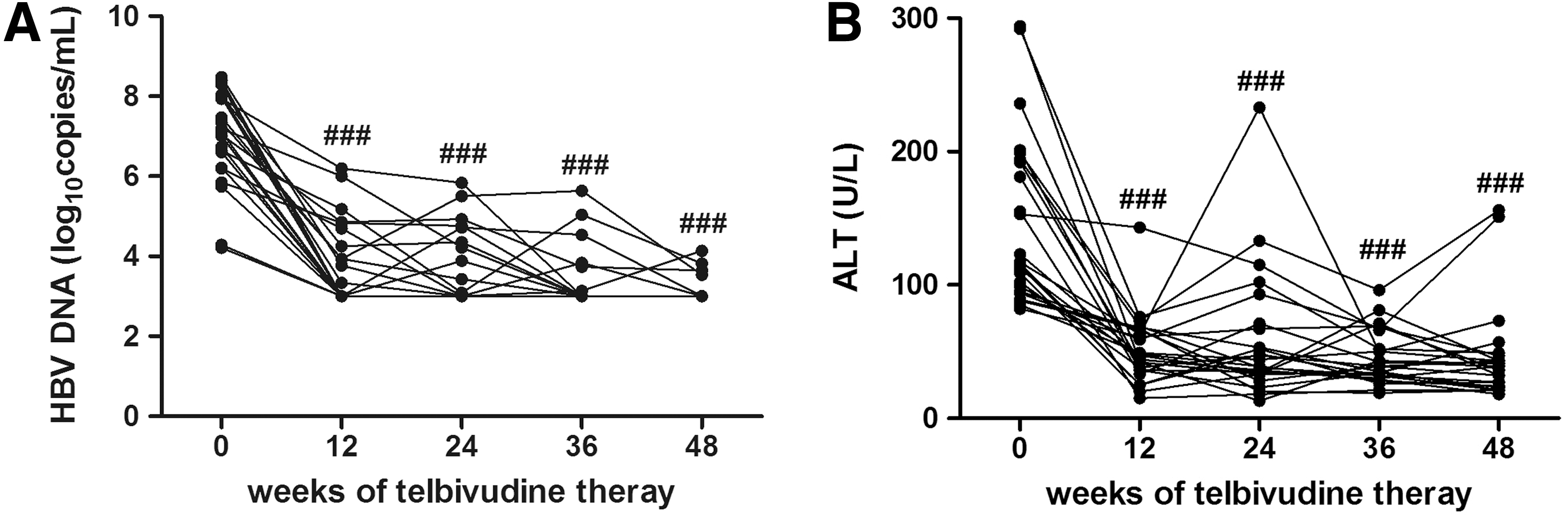
Inhibition of viral replication and normalization of ALT during telbivudine therapy.
Reduction of HBV-specific Th17 cells proportion during telbivudine-induced inhibition of viral replication
To assess the effect of telbivudine therapy on the Th17 immune response, we first determined the IL-17 secretion in response to either PMA+ionomycin or HBV peptides stimulation. Mean fluorescence intensity (MFI) values corresponding to CD4 were notably downregulated by PMA stimulation. Thus, the percentage of CD4+ T cells can not be determined as those cells gated without stimulation. Therefore we gated the CD3+CD8-IL-17A+ cells as the Th17 cells challenged with mitogens. The typical flow cytometry analyses of the proportions of Th17 cells were shown in Figure 2A. The frequency of viral non-specific Th17 cells accounted for CD4+ T cells was 1.905±1.353 % (baseline), 1.932±1.484 % (12 weeks), 1.829±1.375 % (24 weeks), 1.606±1.556 % (36 weeks), 1.754±1.300 % (48 weeks), respectively (p=0.690). No significant changes showed of nonspecific Th17 population in response to antiviral treatment (Fig. 2B). Moreover, there was a consistent trend of reduction of HBV-specific Th17 cells after 12 weeks of telbivudine therapy, but these differences failed to achieve significance (baseline 1.292±0.693 %, 12 weeks 1.011±0.449 %, p=0.225). Furthermore, we found to remarkable decrease in the percentage of viral-specific Th17 cells 36 and 48 weeks after treatments when compared with baseline (24 weeks 0.743±0.439%, 36 weeks 0.678±0.341%, 48 weeks 0.583±0.362%, p=0.049, p=0.0011, and p=0.0007, respectively, Fig. 2C). In addition, a weak correlation was found between the reduction in ALT and the proportion decrease in HBV-specific Th17 cells (r=0.386 and p=0.025), but no correlation was observed between the decrease in HBV DNA and in the proportion of Th17 cells (data not shown).
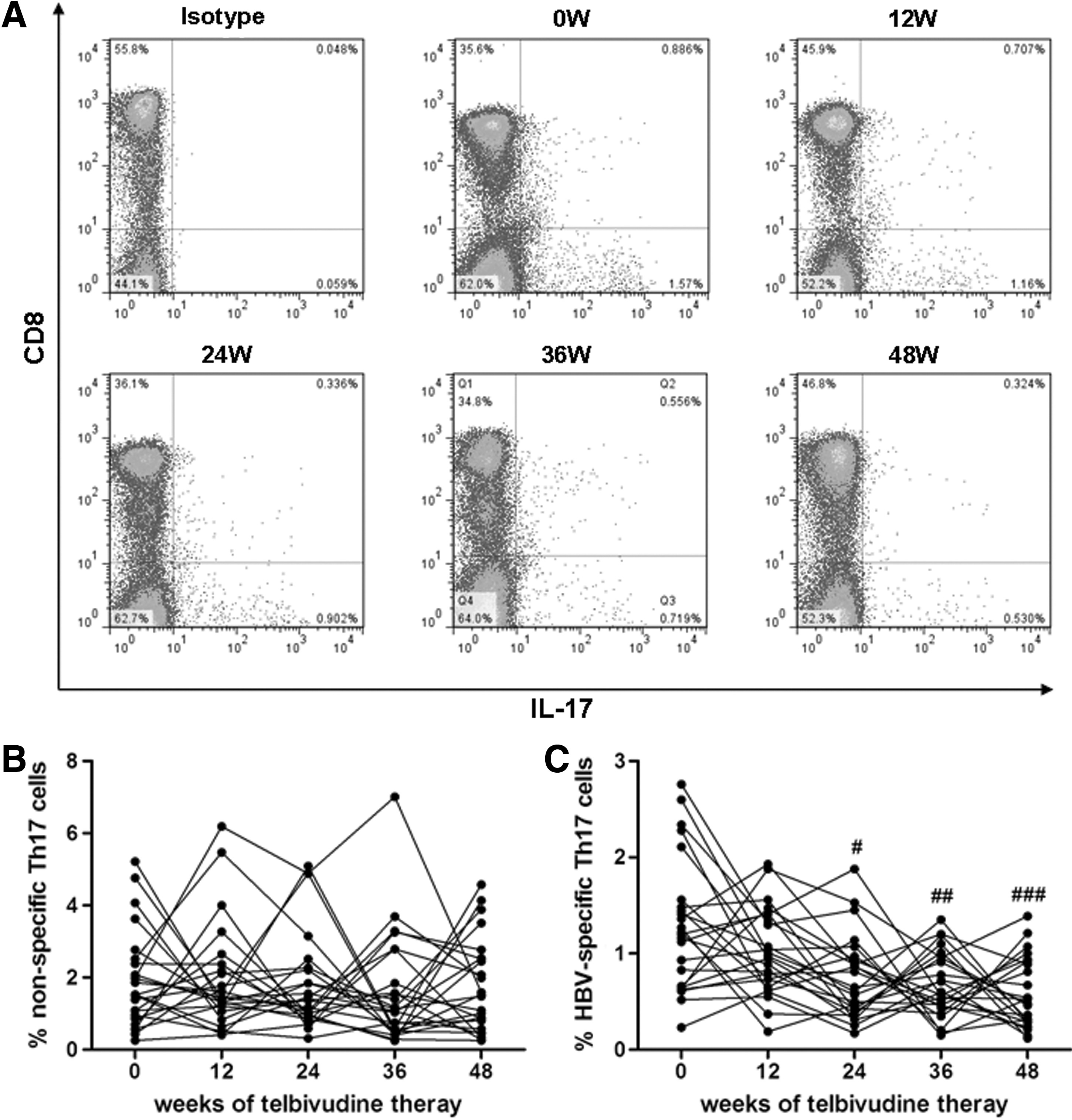
The frequency of peripheral Th17 cells during telbivudine-induced HBV suppression. Isolated PBMCs from a representative enrolled subject were separated using the gates shown by flow cytometry after stimulated with PMA and ionomycin or HBV peptides were stained with labeled antibodies. IL-17A expression in CD3+CD8- subsets (CD4+ subsets) was shown as Th17 cells.
Decrease of serum IL-22 and IL-23, but not IL-17 in response to telbivudine therapy
Th17 secreting cytokines, IL-17 and IL-22, as well as Th17 differentiation cytokine IL-23 were tested at baseline, and after telbivudine treatment. The concentrations of IL-17 were 19.70±5.392 pg/mL (baseline), 20.21±12.01 pg/mL (12 weeks), 19.43±12.27 pg/mL (24 weeks), 16.34±9.350 pg/mL (36 weeks), and 14.90±5.167 pg/mL (48 weeks), respectively, and there was no notable difference during the observation period (Fig. 3A). In addition, IL-22 level was moderately decreased 12 weeks after initiation of telbivudine therapy (baseline 39.76±13.68 pg/mL, 12 weeks 30.62±13.04 pg/mL, p=0.023), while the concentration was significantly reduced in response to the antiviral treatment (24 weeks 27.11±9.855 pg/mL, p=0.0026; 36 weeks 22.36±5.198 pg/mL, p<0.0001; 48 weeks 26.32±10.41 pg/mL, p=0.0013, Fig. 3B). Moreover, a slow reduction of IL-23 expression was found in response to telbivudine therapy. IL-23 levels were downregulated slowly at 12 and 24 weeks (baseline 8.523±7.065 pg/mL, 12 weeks 6.803±4.667 pg/mL, 24 weeks 4.835±1.979 pg/mL, p>0.05), and the concentrations were maintained in low levels after 36 and 48 weeks treatment (36 weeks 4.367±4.345 pg/mL, P=0.0005, 48 weeks 3.145±2.384 pg/mL, p=0.0001, Fig. 3C).

Serum IL-17, IL-22, and IL-23 concentrations during telbivudine therapy.
Reduction of IL-22 secretion in response to HBsAg stimulation during telbivudine therapy
To evaluate the viral specific cytokine secretion during telbivudine treatment, PBMCs from 10 patients of each observation point were seeded into 96-well plates with recombinant HBsAg stimulation. The supernatants were harvested 4 days later and IL-17, IL-22, and IL-23 concentrations were measured by ELISA. Unexpectedly, there were very little IL-23 productions in response to HBsAg stimulation. IL-17 concentrations revealed no significant changes in the supernatants during the observation period (baseline 260.2±193.6 pg/mL; 12 weeks 276.2±162.0 pg/mL; 24 weeks 208.4±117.8 pg/mL; 36 weeks 199.1±118.2 pg/mL; 48 weeks 222.0±84.03 pg/mL, p=0.890, Fig. 4A). However, HBsAg-induced IL-22 was continuously suppressed in response to HBsAg stimulation (baseline 66.01±25.97 pg/mL; 12 weeks 63.35±20.06 pg/mL; 24 weeks 54.07±16.24 pg/mL; 36 weeks 49.30±12.92 pg/mL), and a remarkable reduction was found 48 weeks after telbivudine treatment (48 weeks 25.30±14.67 pg/mL, p=0.002 compared with baseline, Fig. 4B).
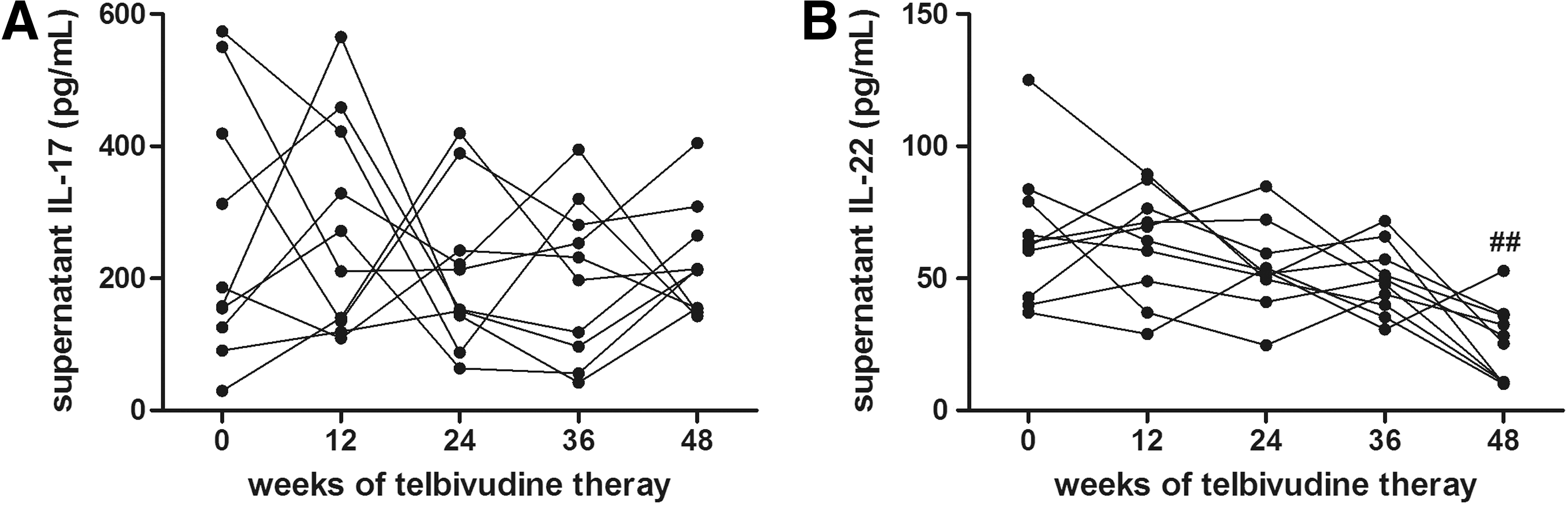
HBsAg-specific IL-17 and IL-22 production in response to telbivudine treatment. PBMCs from 10 of CHB patients were stimulated with HBsAg for 4 days in 96-well plates. Supernatants were harvested and expression of IL-17
Discussion
Telbivudine is a β-L-nucleoside analogue of thymidine that inhibits HBV replication by obligating chain termination of DNA synthesis (27). The GLOBE study demonstrated that telbivudine revealed superior antiviral efficacies to lamivudine regardless of the HBeAg status (21,25). In the present study, we examined telbivudine therapy of HBeAg-positive chronic hepatitis B patients in China, and showed that 83.33% (20/24) of patients achieved seroclearance of HBV DNA and 75.00% (18/24) with normalization of ALT after 48 weeks of treatment. All patients were well-tolerated to telbivudine. The current results were consistent with the previous large-cohort reports in China (12), and further confirmed that telbivudine is an effective and safe drug for the therapy of chronic HBV infection.
Recent studies showed that treatment of telbivudine could not only inhibit viral replication and improve liver function, but also modulate immune response of patients with CHB. Telbivudine therapy increased the numbers/frequencies of CD4+ T cells (4), CD8+ T cell and NK cells (28), as well as improved the function of CD4+ T cells (47), myeloid dendritic cells (31), iNKT cells (36), and cytokines secretion (4). On the other hand, antiviral treatment with telbivudine reduced the proportion of CD4+CD25high regulatory T cells (30) and downregulated the expression of PD-1 and PD-L1 (8). However, little study focused on the numbers and functions of Th17 cells in response to telbivudine therapy. In the current study, we found that the HBV-specific Th17 cells were decreased during anti-HBV therapy. However, viral nonspecific Th17 cells did not reveal significant difference during the observation period. In contrast, Zhang et al. found that nonspecific Th17 cells showed a reverse “V”-type change with the peak level at 1 month after entecavir treatment (43). It have been proven that Th17 cells promoted persistent viral infection and associated with chronic inflammatory diseases (13). In HBV infection, peripheral and intrahepatic nonspecific Th17 cells positively correlated with HBV DNA load, serum ALT, and histological activity index (44,45). We found that HBV-specific Th17 cells were reduced in accordance with inhibition of viral replication and normalization of ALT in response to telbivudine treatment, but only decrease of ALT demonstrated a statistical correlation with Th17 reduction. Previous study has demonstrated that pathogen-infected apoptotic cells could preferentially induce Th17-mediated immunity (38), and HBV infection could also trigger apoptosis of hepatocytes and cause the increase of serum aminotransferases (7,18). Telbivudine treatment may suppress the apoptosis and induction of immune response via inhibition of HBV replication, however, no correlation of viral load suppression and Th17 reduction was found. Altogether this may indicate that HBV-induced immunologic injury and apoptosis, which cause the inflammation and impaired liver function, is associated with viral specific Th17 cells.
Interestingly, the HBV-specific Th17 cells, detected by flow cytometry, were decreased during the course of antiviral treatment; whereas IL-17 secretion in the serum or the supernatant of HBsAg-stimulated PBMCs, detected by ELISA, was not affected by the antiviral therapy. On the one hand, the disparity of flow cytometry and ELISA to detect Th17/IL-17 may contribute to this difference. Flow cytometry tests the cytokine secretion of cells in response to stimulation, which reveal the potential of cytokine productions in designated cell populations. However, ELISA is used to measure the soluble protein expressions in the serum or supernatants, which represent total cytokines from the whole cell populations. Thus, the IL-17 expressions in the serum and supernatants might involve not only CD4+ T cells (mainly Th17 cells) production but also CD8+ T cells secretion (Fig. 2A). On the other hand, we used two epitope-dominant peptides to induce IL-17 secretion to identify Th17 cells population, whereas recombinant HBsAg protein were used to stimulate the PBMCs for measuring cytokines productions in the supernatants. Furthermore, the incubation periods were also varied with 12 h for flow cytometry and 4 days for ELISA. Thus, the current condition to induce Th17 cells and IL-17 productions in vitro might be another reason to cause the differences.
IL-23 plays an important role in stabilization of Th17 differentiation and production of IL-17 and IL-22 (20). IL-23 could amplify the Th17 cells mediated immunopathogenesis of HBV infection (15). However, the mechanisms of Th17 secreting cytokines and IL-23/Th17 cells axis in HBV induced liver injury is still not well elucidated. Previous study has provided evidence that hepatocytes from mice deficient in IL-22 are highly sensitive to the concanavalin A-induced acute hepatitis, and IL-22 expressing Th17 cells could provide protection during hepatitis in IL-22 deficient mice, but IL-17 has no observable role in liver inflammation (42). Similar results were found in HBV-infected patients, as IL-22 but not IL-17 were elevated expression in the serum of acute hepatitis B, but a proinflammatory role of IL-22 was found in the immune response to HBV infection (45). Our results revealed the downregulation of both HBsAg-specific and nonspecific IL-22 production gradually declined during telbivudine therapy, however, IL-17 expression did not show notable differences in response to treatment. Together with previous studies in which they observed that IL-22 was positively correlated with inflammatory environments (45), the current results revealed that IL-22 but not IL-17 may be the major effector cytokine that secreted by Th17 cells during HBV infection and may serve as a protective molecule to counteract and limit the severe injury of liver function that caused by destructive effect of the immune response.
IL-23/Th17-mediated immune response plays a critical role in the pathogenesis of several intestinal inflammatory and autoimmune diseases (37). Downregulation of IL-23 expression was found in response to telbivudine treatment. The reduction of viral specific Th17 and IL-23 showed a similar trend during anti-HBV therapy, but this consistent trend failed to achieve significant correlation (data not shown). Due to the importance of IL-23 pathway in Th17 development, we speculated that reduction of HBV DNA and normalization of ALT by telbivudine may decline viral-specific Th17 cells response via reduction of IL-23 expression. Further studies should be performed to examine the factors that involved in the regulation of IL-23/Th17 cells axis in HBV infection.
Conclusion
In summary, telbivudine could induce the inhibition of viral replication and normalization of ALT. This results in a suppression of HBV-mediated inflammatory response, as symbolized by a significant reduction in the percentages of HBV-specific Th17 cells and IL-22 production. The current findings suggest that the antiviral efficacy of nucleoside analogs may contribute not only to the direct viral suppression but also to the decreased inflammation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.