
Abstract
B cells play an important role in humoral immunity and antibody production. Use of a B cell ELISPOT assay to quantify antigen-specific B cells can assist other assays to achieve a more complete profile of the humoral immune response after vaccination. We utilized a B cell ELISPOT assay to measure the number of influenza A/H1N1-specific B cells at key timepoints after seasonal influenza vaccination in 106 older adults (50–74 years of age). Blood was drawn from these subjects on Day 0, Day 3, Day 28, and Day 75 after vaccination to represent baseline, early, peak, and late response, respectively, of influenza A/H1N1-specific B cells. A significant increase in A/H1N1-specific B cells (median 36 spot-forming units/SFUs per 200,000 cells, p<0.0001) was seen on Day 28 compared to baseline and Day 3, and this number decreased (23 SFUs, p<0.0001) by Day 75, but not to baseline level. These data suggest that the B cell ELISPOT can be used to profile and monitor the humoral immune responses in older subjects after influenza vaccination, and serve as an immune signature marker.
Introduction
T
A key aspect in the effectiveness of the influenza vaccine is its ability to activate the adaptive immune system, preparing the host to generate a quicker response during subsequent infections (24,27). Central to humoral adaptive immunity is the production of antibodies that are effective at binding and neutralizing viruses. The detection and quantification of influenza-specific serum antibodies provides information regarding an individual's immune system preparedness for a subsequent influenza infection. This is commonly achieved through the hemagglutination inhibition (HAI) assay and viral neutralization (VN) assay, which detect influenza-specific antibody levels in serum. Historically, the HAI titer is used as a correlate of protection when measuring vaccine-induced humoral immunity (7,10,22). Influenza vaccine failure in high-risk subpopulations has called into question whether HAI and VN data are sufficient in assessing humoral immunity preparedness for an influenza A/H1N1 infection. Other measures of humoral immunity could be further assessed to address the discrepancy seen in the high percentage of influenza vaccine failures (18,23).
One such approach to measure influenza humoral immunity is the quantification of influenza-specific B cells. The recognition of the influenza virus by B cells and production of influenza-specific antibodies are crucial for the amplification of the immune response and neutralization of the influenza virus. While the VN and HAI assays provide important data regarding humoral protection from infection, influenza-specific B cell counts provide an alternative perspective for assessing humoral immunity preparedness for a subsequent influenza infection (4,9,12). Therefore, influenza-specific B cells could serve as important markers of active humoral immunity, and these data can be used along with other measures of immunity to predict influenza vaccine efficacy. Precise quantification of the humoral immune response after influenza vaccination can be achieved using an influenza-specific B cell enzyme-linked immunosorbent spot (ELISPOT) assay. Since the first description of the B cell ELISPOT as a detection of specific antibody-secreting cells was published (5), the assay has been used to study vaccine-induced B cell immune responses due to its specificity by pre-coating plates with specific antigen. The time-course of the influenza-specific B cell immune response is studied by obtaining peripheral blood mononuclear cells (PBMCs) from blood samples taken at different timepoints pre- and post-vaccination.
While virus-specific B cell ELISPOTs have been used in the past to test post-vaccination humoral immunity, there are limited data that quantify the influenza A/H1N1-specific B cell counts post-vaccination in an older adult and elderly cohort. We recruited a relatively large older and elderly cohort (n=106) in order to assess and compare post-vaccination influenza-specific B cell counts to existing literature. The aim of this study was to utilize an ELISPOT assay to profile the humoral immune response to the influenza A/H1N1 virus by monitoring numbers of influenza A/H1N1-specific memory-like B cells at key timepoints after seasonal influenza vaccination in an older adult and elderly population.
Materials and Methods
Study subjects
Study subjects (ages 50–74 years) were recruited at the Mayo Clinic. These subjects were generally healthy with no immunocompromising conditions, including medication regimens, no new diagnoses in the 3 months preceding enrollment, and who have had no drug or treatment changes in the 3 months preceding the enrollment. Subjects were excluded from enrollment or subsequent participation if they exhibited symptoms consistent with influenza prior to or during the study. Study subjects received the 2010–2011 licensed trivalent influenza vaccine, which contains the influenza A/California/7/2009 H1N1-like, A/Perth/16/2009 H3N2-like, and B/Brisbane/60/2008-like viral strains, at enrollment in the study. Written informed consent from subjects was obtained at the time of enrollment. The Mayo Clinic Institutional Review Board granted approval for the study.
Isolation of peripheral blood mononuclear cells (PBMC)
The methods described below are similar or identical to those published for our previous studies (29,30). PBMCs were isolated from 100 mL of whole blood at timepoints pre-immunization (Day 0) and post-immunization (Day 3, Day 28, and Day 75) from study subjects who received one documented dose of the 2010–2011 seasonal trivalent inactivated influenza vaccine.
Cell processing was performed using cell preparation tubes with sodium citrate and a standard laboratory protocol, as previously described (29). Purified PBMCs were resuspended at a concentration of 1×107/mL in freezing medium [RPMI 1640 medium containing L-Glutamine (Invitrogen, Carlsbad, CA) supplemented with 10% dimethyl sulfoxide (DMSO; Protide Pharmaceuticals, St. Paul, MN) and 20% fetal calf serum (FCS; Hyclone, Logan, UT)] frozen overnight at −80°C, and then transferred to liquid nitrogen for further storage, as previously described (29,30).
Preparation of infuenza virus stock
The influenza A/California/7/2009/H1N1-like virus (Centers for Disease Control and Prevention, CDC, Atlanta, GA) was propagated in 9-day-old embryonated chicken eggs at 37°C and 82% humidity. The allantoic fluid was harvested 48 h post-inoculation and influenza virus titers were determined by hemagglutination (HA) and the Tissue Culture Infectious Doses 50% (TCID50) method in MDCK cells using standard protocols (32 –34). To reduce variability, one viral stock was used for all assays.
Influenza A/H1N1-specific B cell ELISPOT assay
An influenza virus-specific ELISPOT assay was used to quantify influenza virus-specific B cells in subjects' PBMCs using the Mabtech ELIspotPLUS kit for human IgG (Mebtech Inc., Cincinnati, OH) and the manufacturer's instructions as previously described with a few modifications (4,19). In brief, 96-well Millipore Immobilon-P-Membrane multiscreen filter (PVDF) plates (EMD Millipore, Billerica, MA) were pre-coated with whole influenza A/H1N1 virus at a dilution of 1/10 (50,000 TCID50 per well) in phosphate-buffered saline (PBS, pH 7.4), or with 15 μg/mL of anti-human total IgG capture mAb MT91/145 (1.5 μg per well), and incubated overnight at 4°C. For each subject, one control well was coated with PBS, pH 7.4, to represent a subject-specific background measure (negative control; coating with PBS was shown to give similar background measures as coating with a virus-free allantoic fluid [data not shown]). For each subject, we quantified the influenza virus-specific response in four wells and the total IgG response (a positive control) in triplicate. Cryopreserved PBMCs were thawed as previously reported (29), extensively washed, counted and pre-stimulated in 24-well plates (4×106 cells/well) with human recombinant IL-2 (final concentration 10 ng/mL; Mabtech ELIspotPLUS kit for human IgG, Mebtech Inc., Cincinnati, OH) and R848, a TLR7/8 agonist, (final concentration 1 μg/mL; Mabtech ELIspotPLUS kit for human IgG, Mebtech Inc.) at 37°C in a 5% CO2 humidified incubator for 72 hours. Before assay setup, ELISPOT plates were washed (5X with sterile PBS, pH 7.4) and blocked for 1 h at room temperature with RPMI medium containing 10% FCS. Pre-stimulated PBMCs were collected, washed to ensure removal of any secreted antibodies, counted and plated in the antigen-coated ELISPOT plates at 2×105 cells/well (for the influenza virus-specific response and the negative control) or at 1×104 cells/well (for the total IgG response, positive control) in RPMI medium containing 5% FCS. Plates were then incubated for 20 h at 37°C, in 5% CO2. After the incubation period, the plates were extensively washed (5X with PBS, pH 7.4) and incubated with a detection biotinylated anti-human IgG mAb MT78/145 (at 1 μg/mL in PBS-0.5% FCS) for 2 h at room temperature, following the manufacturer's specifications. Plates were scanned and analyzed using pre-optimized counting parameters on an automated ImmunoSpot® S6Macro696 Analyzer (Cellular Technology Ltd., Cleveland, OH) with the ImmunoSpot® version 5.1 software (Cellular Technology Ltd.). Quality control (QC) was performed by a single operator to ensure consistency of QC and results consistency (29). The results are presented in spot-forming units (SFUs) per 2×105 cells.
Statistical analyses
The influenza virus-specific B cell ELISPOT response is defined as the subject's median influenza virus-specific response (measured in four wells) minus the background response (measure in one negative control well coated with PBS). The summary measures are presented as medians and interquartile ranges (IQRs: 25th, 75th percentiles). Results are presented as percentiles of the distributions, and are presented graphically using box-and-whisker plots and histograms. Differences between the B cell ELISPOT responses at different timepoints were compared using the nonparametric Wilcoxon signed-rank test. Intra-class correlation (ICC) was measured between the four influenza virus-specific wells using the Shrout and Fleiss's method (26).
Results
Study subjects
The study cohort of 106 eligible subjects consisted of 65 (61.3%) females and 41 (38.7%) males with a median age of 59.7 years (IQR 55.3, 67.6). One hundred four subjects (98.1%) were Caucasian, and two of the subjects were of other races (i.e., one subject was Asian and one was multi-race) (Table 1).
QR, interquartile range.
Influenza-specific B cell response kinetics after vaccination
We selected each timepoint to represent a key moment in the immune response to vaccination or infection. Day 0 represents the baseline level of immune status, and T and B cell-related immune measures are at baseline prior to vaccine administration. Day 3 represents the early phase of the immune response when the innate pathways are activated and viral antigens are being presented by antigen presenting cells (APCs). Day 28 represents the adaptive immune response is at its apex, and B cell proliferation and differentiation into plasma cells has resulted in high-titer antibody production. This is when the initial peak of antigen-secreting cells (ASCs) seen at days 5–10 has subsided and influenza virus-specific memory-like B cells are considered to be at peak levels (4,34). On Day 75, primary lymphocyte responses have subsided and memory B cell population has returned to a homeostatic resting state.
Figure 1 provides an example of a developed ELISPOT plate that we used to collect our data. Table 2 includes the overall median B cell counts for each timepoint, including IQRs. The minimum and maximum B cell counts, along with 10th and 90th percentile B cell counts, are included at each day's range of influenza A/H1N1-specific B cell counts. Figure 2 and Figure 3 provide further illustration of the collected data seen in Table 2. The influenza A/H1N1-specific B cell ELISPOT values are relatively low at Day 3 with eight SFUs per 200,000 cells (IQR 3, 20), and then increase to the peak response at Day 28 with 36 SFUs (IQR 16, 60), with a decrease at Day 75 to 23 SFUs (IQR 11, 38). A significant increase in the influenza A/H1N1-specific B cells occurs by Day 28 compared to Day 0 (p<0.0001) and Day 3 (p<0.0001). On Day 75, when both the humoral and cellular response has subsided and memory cells primarily remain, the number of influenza A/H1N1-specific B cells is lower (compared to Day 28, p<0.0001), yet higher when compared to the responses at Days 0 and 3 (p<0.0001). Intra-class correlation coefficients, measuring the correlation between replicate measurements, for our B cell ELISPOT assay were high (overall ICC=0.91, Table 4).

A representative 96-well ELISPOT plate used to quantify influenza A/H1N1-specific IgG B cells, as described in the Methods section. Each set of four columns contained three subjects tested at the four timepoints. For example, columns 1–4 represent one subject's B cell counts from Day 0, 3, 28, and 75, respectively. Row A was the negative control, rows B–D the positive control of total IgG, and rows E–H were pre-coated with the influenza A/H1N1 virus, as described in the Methods section.
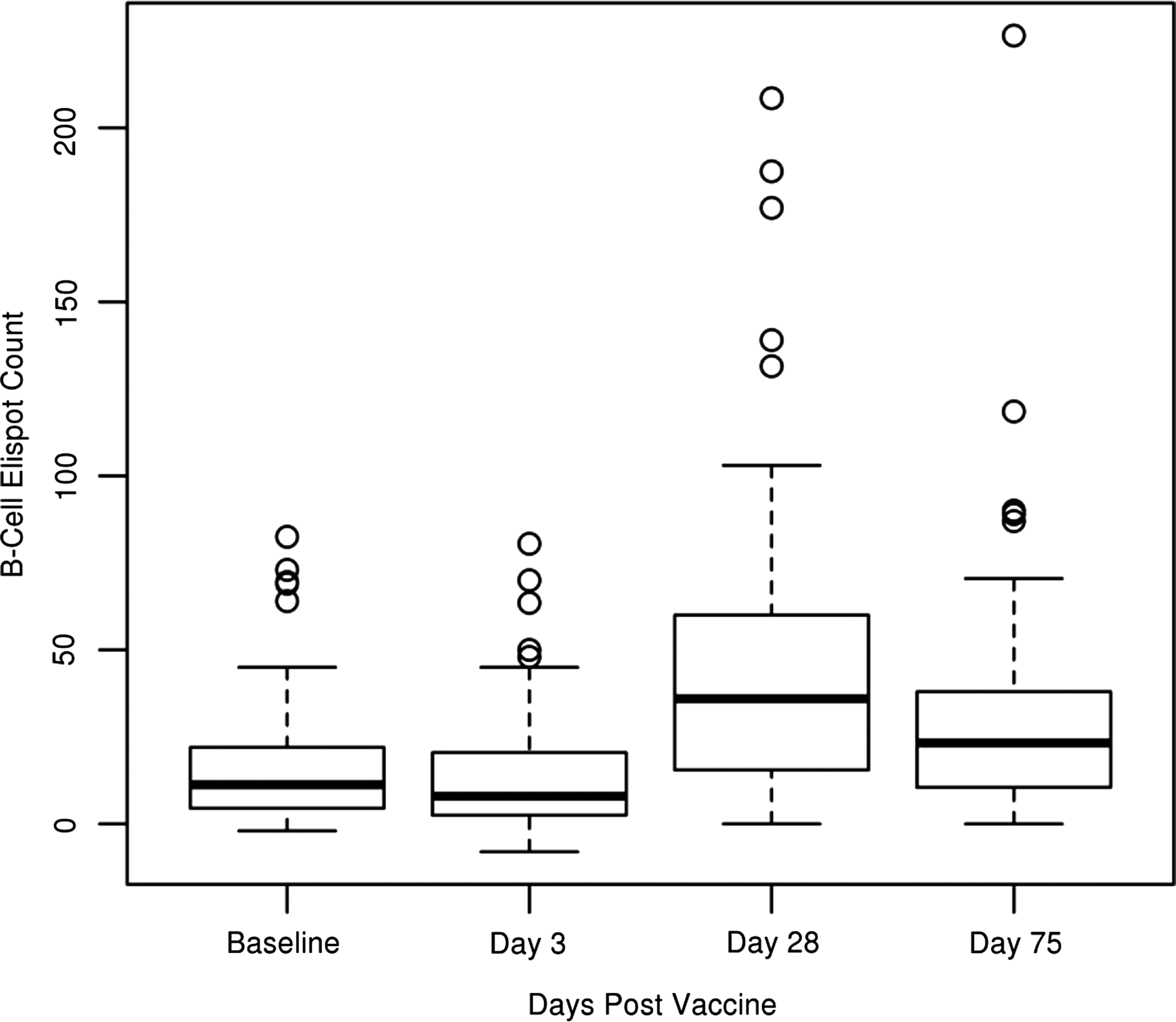
Box and whisker plots of the total (n=106) study subjects' pre-vaccination (Baseline) and post-vaccination (Day 3, Day 28, and Day 75) counts for the influenza A/H1N1-specific IgG-producing memory-like B cells. B cell ELISPOT counts are represented as the average spot-forming units (SFUs), inter-quartile ranges, minimum and maximums of 2×105 cells per well.
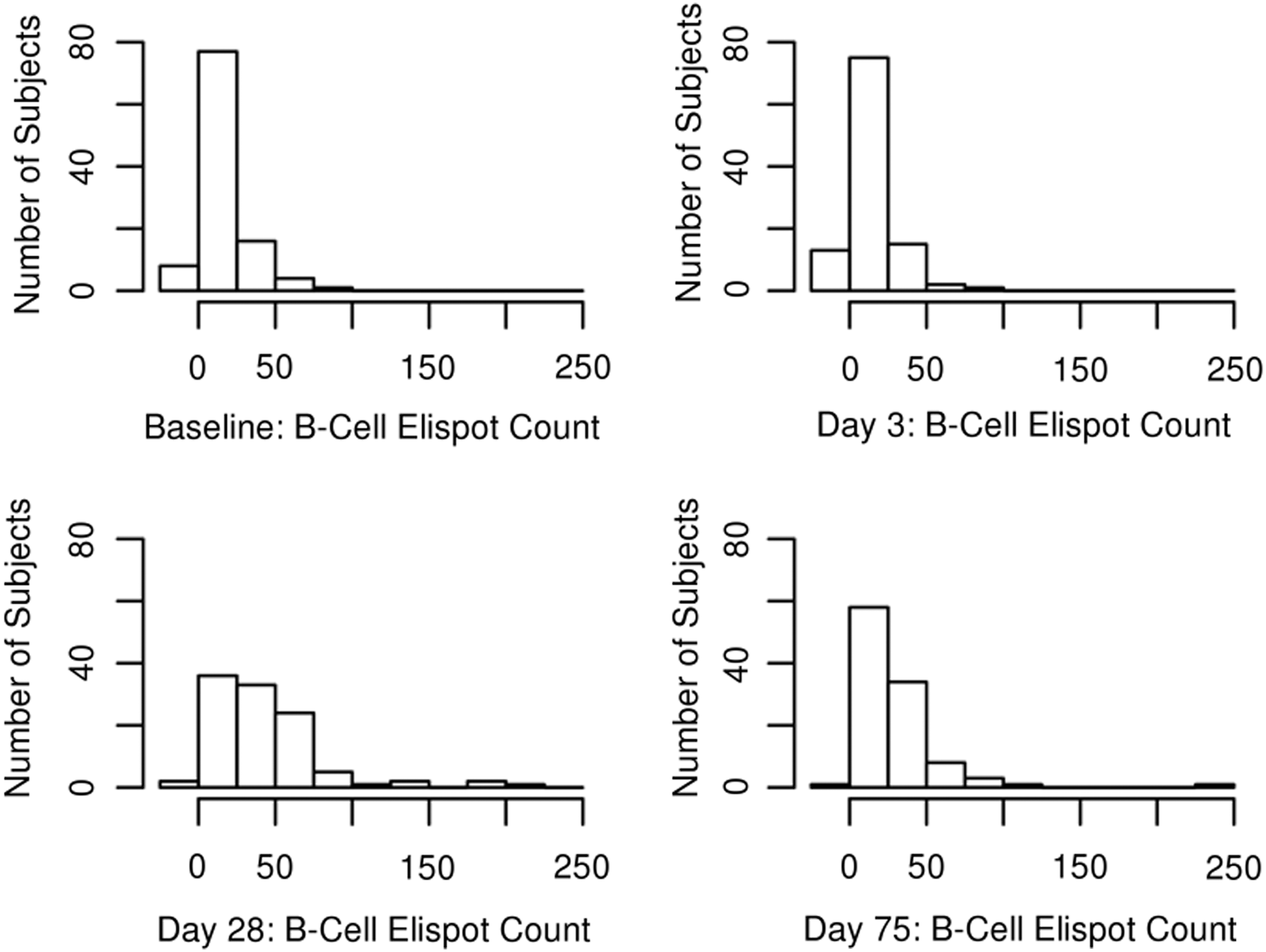
Histograms illustrating the change in distribution over time in the influenza A/H1N1-specific IgG-producing B cell counts at each timepoint, measured in spot-forming units (SFUs).
Median of the stimulated samples—unstimulated sample.
We compared fold-change ratios (Table 3) in order to observe the magnitude of response in influenza A/H1N1-specific B cell counts from one timepoint to a second timepoint. For example, a value of 1 would mean the subject experienced no increase or decrease in response between the timepoints being compared. As expected, the average magnitude of B cell responses increased by a factor of 2.49 (IQR 1.09, 5.42) from Day 0 to Day 28, and decreased by Day 75 with measurable counts above baseline levels (median 1.77; IQR 0.3, 3.28). A slight drop was observed in the magnitude of B cell counts from Day 0 to Day 3 (0.67; 0.33, 1.16; p=0.001).
Total number of samples included. Some subjects were excluded due to 0 influenza A/H1N1-specific B cells at a timepoint that formed a ratio with an undividable number in the denominator.
Discussion
In order to address the challenge of decline in influenza vaccine response in the elderly, current research outlines the need to determine other immune correlates to assess influenza-specific immune response to infection (17 –19, 23). Exposing the immune system to influenza-specific antigens, such as the hemagglutinin (HA) and neuraminidase (NA) surface glycoproteins, activates the adaptive immune system to provide an influenza-specific response. A key step in increasing circulating protective antibodies is the proliferation and differentiation of efficient antigen-specific B cells. Naïve B cells differentiate into plasma B cells and memory B cells that provide the host immune system with short-term and long-term influenza virus-specific humoral protection (3,4,13,16,25). While the ASC/plasmablast population peaks near Days 5–10 post-vaccination, memory B cells peak near Day 28 and function to reinitiate the influenza strain-specific humoral immune response after a subsequent infection (4,34). Enumerating the influenza-specific B cells could provide an alternative assessment of humoral immunity and protection against a subsequent infection. Combining the highly sensitive B cell ELISPOT with other assays would help create a more comprehensive understanding of the humoral immune response after vaccination, leading to the development of improved influenza vaccines.
While influenza-specific B cell ELISPOTs have been utilized to test post-vaccination humoral immunity, limited data exists for the influenza-specific memory-like B cell response in an elderly population. In our study, we used an ELISPOT assay to provide precise quantification of the influenza A/H1N1-specific B cells, pre- and post-vaccination, in a relatively large older and elderly cohort. By using the B cell ELISPOT assay, we were able to enumerate influenza A/H1N1-specific B cells. Our data confirm that antigen-specific memory B cell detection after vaccination using the ELISPOT assay is practical. Our data show that a peak response in antigen-specific B cells occurred at Day 28 post-influenza immunization. We also observed a significant difference between baseline (pre-vaccination) influenza-specific B cell responses, B cell responses at the peak of the adaptive response, and B cell responses at homeostasis. The reproducibility of our assay was high (intra-class correlation coefficients, ICC=0.91) and is comparable to other ELISPOT assays (11,20). It is interesting to note the decrease seen in influenza A/H1N1-specific B cells from baseline to Day 3 (p=0.001). This could reflect the expansion of IgG+ASCs that originate from the memory compartment and peak Days 5–10 after vaccination (4,34).
Our cohort included older subjects who were between the ages of 50 to 74, which is an age group that has a general decrease in immunity and responsiveness to vaccination. However, our data suggest that a peak in influenza A/H1N1-specific memory-like B cells occurs at Day 28 after vaccination, despite an older (ages 50–74) cohort. Our data and data from the literature suggest that the number of influenza-specific memory-like B cells consistently peaks near Day 28 post-vaccination in young adult (above age 9), adult (ages 21–48), and elderly (age 65 and older) populations (14,25). Wrammert et al. determined the dynamics of humoral influenza-specific B cell responses, observing a peak of IgG antibody-secreting cell (ASC) response at Day 7, while a peak of memory B cells occurred 14 to 28 days post-vaccination (35). Therefore, memory B cell responses near Day 28 can serve as a measurement for humoral immune preparedness.
The strength of this study is that the assay that was used has precise antigen-specificity in detecting B cell counts. Only B cells expressing IgG specific to influenza A/H1N1 are detected and quantified. By using whole virus for the B cell ELISPOT assay, we measure B cells expressing IgG specific for the influenza A/California/7/09 H1N1-like viral strain that we studied. Such specificity makes this assay a strong complement to other assays that provide a more complete assessment of post-vaccination immune responses. For example, when compared to the ELISA assay, ELISPOT provides a rapid quantification of antigen-specific B cells (including memory B cells) instead of measuring antibodies in serum (24,27). In comparison to flow-based assays, ELISPOT is more cost-efficient and has the capability to measure cell function (2).
Another strength of our study is the relatively large size of our study cohort. Jahnmatz et al. recently utilized the B cell ELISPOT assay to detect the presence of memory B cells in a small population of subjects (n=8) with vaccine-induced immunity to five different viral antigens (12). However, no studies based on influenza A/H1N1 vaccine-induced detection of antigen-specific memory B cells in a large elderly population have been performed. We selected an elderly population and specifically focused on influenza A/H1N1-specific memory B cells. Our data support the conclusion that the B cell ELISPOT assay is a highly effective method to detect influenza A/H1N1-specific B cells after vaccination (1,12,23). Several strains of the influenza virus exist due to the known variances in the HA and NA surface antigens. Therefore, our data support the hypothesis that the described ELISPOT assay could be used to detect B cell responses to other strains of the influenza virus.
There are limitations to this study, as whole virus was utilized to detect the presence of IgG on the surface of B cells. The infectious cycle for the H1N1 virus begins with the H1 protein binding to the sialic acid of human cells. Therefore, coating the plates with recombinant H1 viral glycoprotein instead of whole influenza virus would exclusively detect H1-specific B cells, thus measuring the B cell response against the major viral surface protective antigen—the hemagglutinin. This type of assay is observed in the work of Baer et al., who optimized a B cell ELISPOT detection of influenza-specific H5-specific B cells (1). However, the H1 protein is the primary surface antigen known for initial cellular interaction, and neuraminidase (N1) is responsible for the hydrolysis of the sialic acid-H1 complex for viral release. Therefore, we predict that data obtained from performing an H1-specific B cell ELISPOT would be similar to the data that we collected in our study, which utilized whole influenza A/H1N1-specific B cell ELISPOT.
In conclusion, we developed and utilized an influenza A/H1N1-specific B cell ELISPOT assay for monitoring the kinetics of the humoral immune response after vaccination in older adult and elderly subjects. This protocol was successfully used as a method for quantification of influenza-specific humoral immune response in elderly subjects, demonstrating a peak on Day 28. While research data broadly support the use of the B cell ELISPOT assay for the quantification of vaccine-induced B cell responses, our results confirm the applicability of this influenza A/H1N1-specific B cell ELISPOT assay for other influenza vaccination studies that seek to assess humoral immunity. Data from this assay can be used to profile and monitor the humoral immune responses in older subjects after influenza vaccination, as well as other viruses targeted by vaccination, and serve as an immune signature marker.
Footnotes
Acknowledgments
We thank the Mayo Clinic Vaccine Research Group and the subjects who participated in our studies. We thank Caroline L. Vitse for her editorial assistance and Krista M. Goergen for help with statistical analysis. The authors acknowledge support from NIH Grant U01AI089859 for this work. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Author Disclosure Statement
Dr. Poland is the chair of a Safety Evaluation Committee for novel investigational vaccine trials being conducted by Merck Research Laboratories. Dr. Poland offers consultative advice on vaccine development to Merck & Co. Inc., CSL Biotherapies, Avianax, Sanofi Pasteur, Dynavax, Novartis Vaccines and Therapeutics, PAXVAX Inc., and Emergent Biosolutions. These activities have been reviewed by the Mayo Clinic Conflict of Interest Review Board and are conducted in compliance with Mayo Clinic Conflict of Interest policies. This research has been reviewed by the Mayo Clinic Conflict of Interest Review Board and was conducted in compliance with Mayo Clinic Conflict of Interest policies.