
Abstract
Avian influenza viruses (AIV) are of great concern to the worldwide community as well as the poultry industry. Although existing vaccines are successful in limiting the spread of the virus, these vaccines do not eliminate virus shedding into the environment. As a result, it is of great importance to enhance the efficacy of existing AIV vaccines. Therefore, the objective of the present study was to utilize the immunostimulatory Toll-like receptor ligands poly I:C, lipopolysaccharide (LPS), and CpG DNA motifs, either alone or in combination with each other, as adjuvants to enhance the immunogenicity of an inactivated AIV vaccine. Chickens were vaccinated twice, 14 days apart. Antibody-mediated responses were assessed by collected sera and lacrimal secretions, while cell-mediated immunity was assessed by stimulating splenocytes from vaccinated chickens in vitro with the vaccine antigen. The results suggest that CpG alone served as the best single-ligand adjuvant compared to poly I:C or LPS, as it significantly enhanced antibody-mediated responses, as determined by enzyme-linked immunosorbant assay. Furthermore, upon combining CpG with poly I:C, a robust antibody-mediated and cell-mediated immune response was elicited, resulting in an enhanced hemagglutination inhibition titer and splenocyte proliferation respectively. Future studies may be aimed at assessing the efficacy of the poly I:C and CpG combination adjuvant in protecting against AIV infection.
Introduction
T
TLRs are an evolutionarily conserved group of pattern recognition receptors that mediate host responses to pathogens by binding conserved structural motifs on pathogens termed pathogen associated molecular patterns (PAMPs). For example, TLR3 binds double-stranded RNA and synthetic molecules such as polyinosinic:polycytidylic acid (poly I:C), while TLR4 binds lipopolysaccharide (LPS) and TLR21 binds CpG DNA motifs (2). To date, several chicken cell subsets have been shown to express various TLRs including T-cells, B-cells, macrophages, and monocytes (7). Upon binding their ligand, TLRs promote cellular activation and the production of cytokines such as interleukin (IL)-12 and interferon (IFN)-γ. In the case of antigen presenting cells (APCs), TLR stimulation also promotes the up-regulation of major histocompatibility complex class II (MHC-II) and co-stimulatory molecules including CD80, thereby facilitating the elicitation of a robust adaptive immune response (19,29). Although these ligands are highly immunostimulatory on their own, a synergistic effect in cytokine production has been observed when combining certain ligands together, such as poly I:C and CpG-ODN (4,5).
As a result of their immunostimulatory properties, TLR ligands have previously been shown to be efficacious adjuvants for AIV vaccines in chickens (6,12,13,28). However, these previous studies have predominantly focused on ligands for TLR21 (functional homolog of mammalian TLR9 in chickens) and thus the adjuvant potential of other TLR ligands is relatively unexplored in chicken AIV vaccines. More importantly, the adjuvant potential of various TLR ligand combination adjuvants has not yet been explored in chickens. As such, we hypothesized that, in addition to TLR21 ligands, ligands for TLR3 and TLR4 may also demonstrate exceptional adjuvant potential. To this end, the objective of the present study was to utilize ligands for TLRs 3, 4, and 21, either alone or in combination, to enhance the immunogenicity of an inactivated AIV vaccine. The results suggest that CpG-ODN served as the best single-ligand adjuvant, since mean hemagglutination inhibition antibody titers were highest with this adjuvant, and IgM, IgY, and IgA responses were similar or higher than those with poly I:C or LPS. Chickens receiving a combination of CpG with poly I:C had robust antibody-mediated and cell-mediated immune responses that were significantly higher than responses of birds receiving vaccine without adjuvant.
Materials and Methods
Chickens
Specific pathogen-free (SPF) chickens were procured from the Canadian Food Inspection Agency and housed in the isolation facility of the Ontario Veterinary College, University of Guelph, Canada. This research was approved by the University of Guelph Animal Care Committee and complied with the guidelines of the Canadian Council on Animal Care.
TLR ligands
Poly I:C and LPS from Escherichia coli 0111:B4 were purchased from Sigma-Aldrich, while synthetic class B CpG ODN 2007 (5′-TCG TCG TTG TCG TTT TGT CGT T-3′) was purchased from Eurofins MWG Operon. All of the ligands were resuspended in sterile phosphate buffered saline (PBS; pH 7.4).
Avian influenza virus
The low pathogenic avian influenza virus A/Duck/Czech/56 (H4N6) was propagated in 11-day-old embryonated chicken eggs as described (12). To generate the vaccine, virus-rich allantoic fluid was mixed with formalin (final concentration 0.02%) and incubated for 72 h at 37°C with constant stirring. The allantoic fluid was subsequently ultracentrifuged at 66,802 g using a SW 40 Ti rotor (Beckman Coulter, Inc.) at 4°C for 1.5 h, and the supernatant was decanted. The pelleted virus was resuspended in HNE buffer (5.0 mM HEPES, 150 mM NaCl, and 0.1 mM EDTA, pH 7.4), layered on top of a 20% sucrose cushion, and ultracentrifuged again at 66,802 g at 4°C for 1.5 h. The supernatant was decanted, and the virus was resuspended in HNE buffer, and repeated again for a total of two washes. The virus was then resuspended in HNE buffer and stored at −80°C. Total protein was measured using a BCA assay (Thermo Scientific) according to the manufacturer's recommended protocol. Virus inactivation was confirmed both by passaging in Madin-Darby canine kidney (MDCK) cells as well as in 11-day-old embryonated chicken eggs. Integrity of virus hemagglutinin (HA) protein was confirmed by hemagglutination assay.
Vaccine formulations
Ten chickens per group were immunized subcutaneously on days 7 and 21 post-hatch with inactivated H4N6 virus (15 μg/bird) and one of the following adjuvants: poly I:C (100 μg), LPS (25 μg), CpG ODN (10 μg), or a combination of poly I:C+LPS, poly I:C+CpG, or LPS+CpG. Control groups received H4N6 virus only with no adjuvant (referred to as the H4N6 group) or PBS with no antigen (referred to as the PBS group).
Hemagglutination inhibition assay
Serial two-fold dilutions of serum samples collected on day 28 post-primary immunization were incubated with four HA units of H4N6 virus in a 96-well V-bottom plate for 30 min at room temperature. Subsequently, a solution containing 0.5% washed chicken red blood cells (RBCs) was added and incubated for another 30 min at room temperature. The hemagglutination inhibition (HI) titer was determined as the reciprocal of the highest dilution of serum completely inhibiting hemagglutination of RBCs.
IgY, IgM, and IgA enzyme-linked immunosorbant assay
To determine antibody levels against influenza viral antigens, sera and tears were collected from four to six chickens in each group on days 14 and 28 post-primary vaccination. Sera were assessed for antigen-specific IgY and IgM, while tears were assessed for antigen-specific IgA as described previously (12).
In vitro stimulation of mononuclear splenocytes
Mononuclear splenocytes (1×107 cells/mL) were seeded into 48-well plates and stimulated with either 1 μg/mL of inactivated whole-virus, medium, or ConA as a positive control. The cells were incubated at 41°C in a humidified 5% CO2 environment and harvested for RNA extraction at 24 h post-treatment.
Cell proliferation assay
In order to examine cell-mediated immune responses, proliferative responses following stimulation with 1 μg/mL of inactivated whole-virus was performed at day 24 post-primary immunization using Alamar Blue (AbD Serotec) as previously described (8). Briefly, spleens from four birds of each vaccinated and control groups were collected and single cell suspensions of spleen mononuclear cells were prepared as described previously (20). Cells were adjusted to 1×106 cells/mL in RPMI (Invitrogen) containing 10% fetal bovine serum, 200 U/mL penicillin, 80 μg/mL streptomycin, and 25 mg gentamicin, and 100 μL was added to a 96-well cell culture plate. Cultures were stimulated with 1 μg/mL of inactivated whole-virus or with medium (RPMI). Following addition of antigen or control treatments, cells were incubated for 24 h at 41°C in a humidified 5% CO2 environment. Subsequently, 22 μL of Alamar Blue was added to all the wells, and absorbance was recorded after an additional 24 h incubation. The percent reduction of Alamar Blue is proportional to the degree of proliferation and was calculated as previously described (8).
RNA extraction, cDNA synthesis, and real-time polymerase chain reaction
Total RNA extraction and cDNA synthesis was performed as previously described (18). Real-time PCR using SyBR Green was performed on diluted cDNA using the LightCycler® 480 II (Roche Diagnostics GmbH) as previously described (18). Primers were synthesized by Sigma-Aldrich, and the specific sequences are found in Table 1. Relative expression of all target genes was calculated relative to the housekeeping gene β-actin using the LightCycler® 480 Software.
Statistical analysis
Data were analyzed using SAS statistical software v9.2 (SAS Institute). Proc Univariate was used to assess the normality of the distribution of each variable of interest. Hemagglutinin inhibition titers and splenocyte proliferation data were transformed using a logarithmic transformation before analysis. Variables that were normally distributed were analyzed using a general linear model (Proc GLM), followed by Duncan's multiple range test, when models were significant. Variables that were not normally distributed were analyzed using the Kruskal–Wallis nonparametric test, followed by the Mann–Whitney U-test if significant differences were evident among groups. p-Values of<0.05 were considered statistically significant throughout the analysis.
Results
A Poly I:C and CpG ODN combination adjuvant enhances serum IgY, but not IgM or secretory IgA levels
To assess antibody-mediated immune responses, IgM and IgY antibody responses were determined in serum, as well as IgA in lacrimal secretions. On day 14, IgM concentrations did not differ significantly among the eight experimental groups (Fig. 1A). On day 28, IgM concentrations did not differ significantly among the seven groups that received H4N6 antigen (with or without adjuvant). On day 28, serum IgY concentrations in chickens that received a combination of poly I:C and CpG as adjuvant were significantly higher than those in birds that received H4N6 antigen alone (Fig. 1B). On day 14, IgA concentrations did not differ significantly among the seven groups that received H4N6 antigen (with or without adjuvant). On day 28, mean IgA concentrations were highest in chickens receiving CpG as adjuvant, but differences were not significant (Fig. 1C).
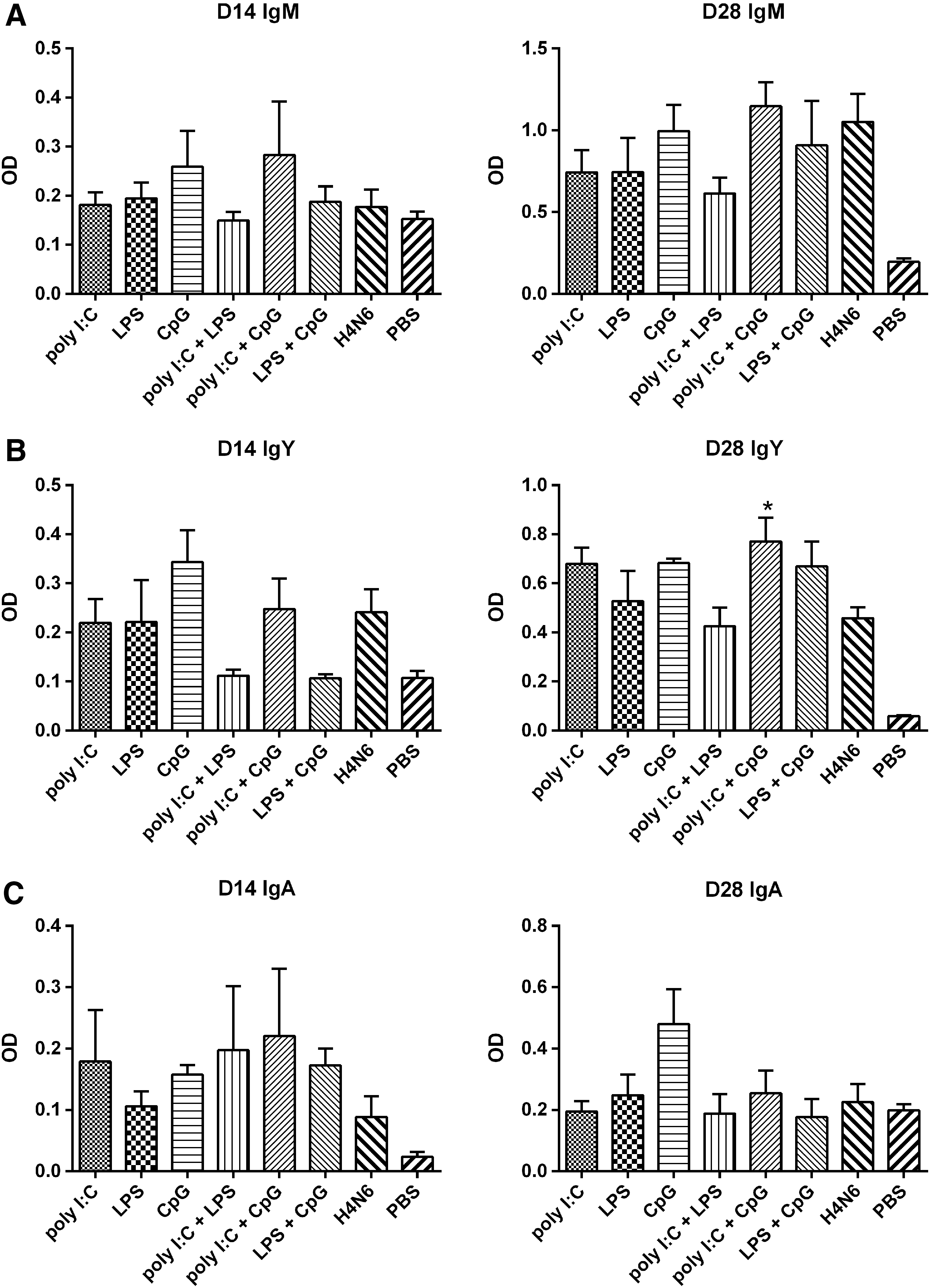
IgM
To gain further insight into the antibody response, we performed a hemagglutination inhibition assay using the serum taken from chickens on day 28 post-primary vaccination. The results indicate that the CpG+poly I:C combination adjuvant significantly enhanced HI titers, with a titer of ∼80 HI units (p≤0.05; Fig. 2). It is also important to note that chickens receiving CpG as adjuvant had higher mean HI antibody titers compared to birds receiving H4N6 alone, but the difference was not statistically significant.
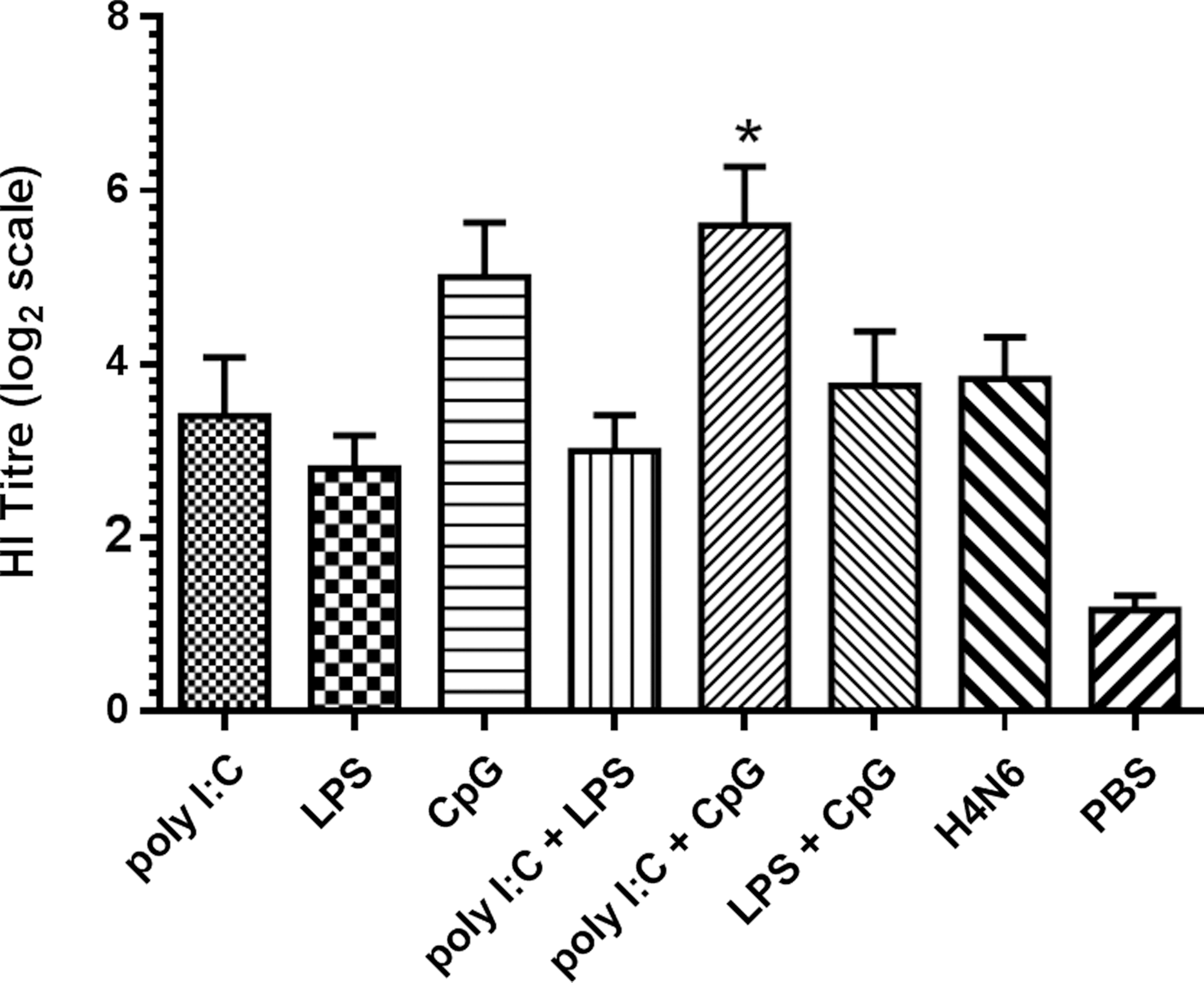
Hemagglutination inhibition (HI) titers of sera collected on day 28 post-primary immunization. Chickens were vaccinated twice with inactivated H4N6 virus in conjunction with either poly I:C (100 μg), LPS (25 μg), CpG ODN (10 μg), or different combinations of these ligands. Control groups received either H4N6 antigen without any adjuvant, or PBS with no antigen. Graphed values represent mean HI titer±standard error. Logarithmically transformed antibody titers were analyzed using a general linear model with Duncan's multiple range test. Comparisons were considered significant when p<0.05 (*).
All three TLR combination adjuvants enhance cell-mediated immune responses
To assess cell-mediated immune responses, we first performed a proliferation assay and then assessed cytokine production in splenocytes isolated from birds on day 31 post-primary vaccination (Figs. 3 and 4). In splenocytes isolated from chickens vaccinated with adjuvants consisting of any combination of TLR ligands, proliferative responses were observed in response to the recall antigen in vitro, which were significantly higher than those for birds that received H4N6 antigen alone (Fig. 3). Proliferative responses for birds receiving combinations of TLR ligands were also significantly higher than responses for birds receiving any of the single ligands.
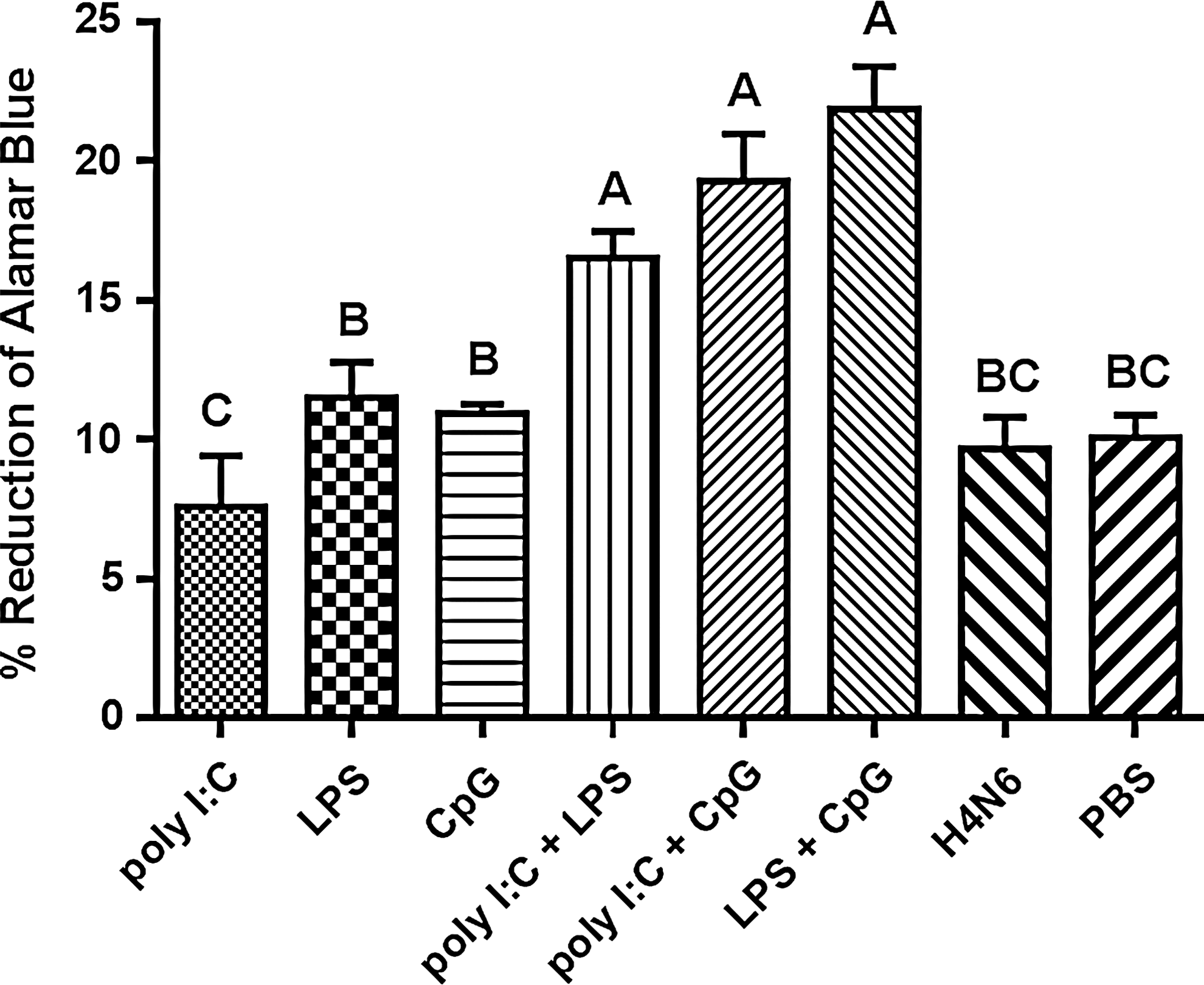
Proliferative responses in splenocytes of vaccinated chickens on day 24 post-primary immunization. Splenocytes were stimulated with 1 μg/mL of inactivated H4N6 and proliferation was assessed 48 h post-stimulation using Alamar Blue. Data represent mean percent reduction of Alamar Blue±standard error. Logarithmically transformed proliferation data were analyzed using a general linear model with Duncan's multiple range test. Group means with the same letter are not significantly different.
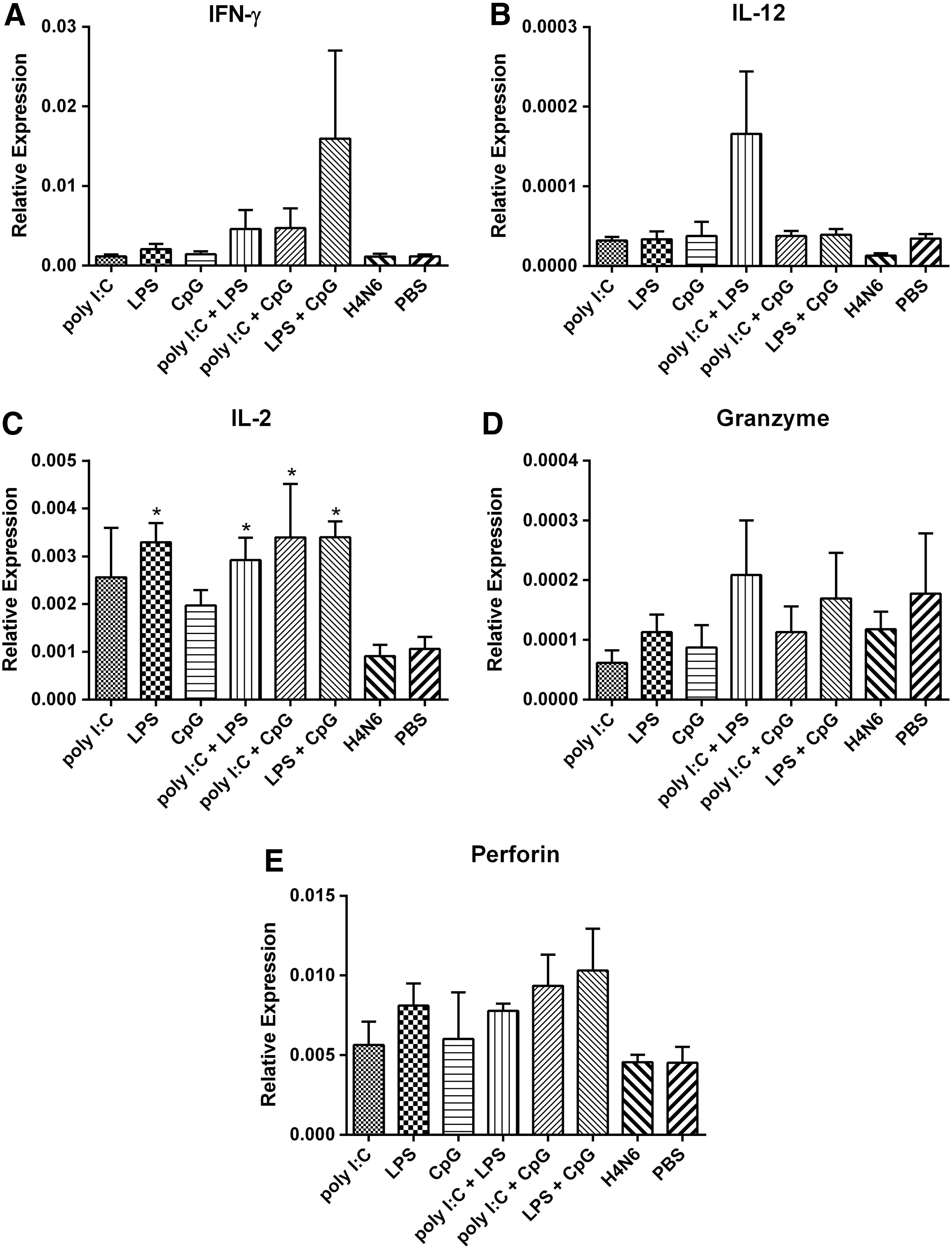
Gene expression in splenocytes isolated from vaccinated chickens on day 24 post-primary immunization. Splenocytes were stimulated with 1 μg/mL of inactivated H4N6 and harvested for RNA extraction 24 h post-stimulation. Data represent mean relative expression levels of IFN-Y
Effects of TLR ligands on expression of cytokines were variable. Mean expression of IFN-γ was highest for chickens receiving combinations of adjuvants, but the differences were not significant (Fig. 4A). Mean expression of IL-12 was highest for chickens receiving a combination of poly I:C and LPS, but the differences were not significant (Fig. 4B). Chickens receiving LPS, or any of the three adjuvant combinations had significantly higher IL-2 expression compared to birds receiving H4N6 antigen alone (Fig. 4C).
Expression of granzyme did not differ significantly among the eight experimental groups (Fig. 4D). Mean expression of perforin was higher in chickens receiving vaccine with any adjuvant, compared to birds receiving H4N6 antigen alone, but the differences were not significant (Fig. 4E).
Discussion
The immunostimulatory properties of TLR ligands have been exploited to enhance the immunogenicity of AIV vaccines. Here, we report that a poly I:C and CpG combination adjuvant enhances antibody-mediated and cell-mediated immune responses against AIV antigens.
In many instances, an important component contributing to efficacy of an AIV vaccine is the elicitation of antibodies targeting the HA surface protein (10). In this regard, our results suggest that, of the single TLR ligand adjuvants, CpG ODN was the most efficacious, as it induced the highest mean HI titer, although it was not significantly higher than the mean for the control group receiving H4N6 antigen alone. As previously mentioned, CpG containing adjuvants are well-documented enhancers of antibody-mediated responses against numerous vaccine antigens in chickens, including AIV (12), Newcastle disease virus (11), infectious bursal disease virus (27), and bovine serum albumin (26), among others (17). Having said that, the adjuvant potential of poly I:C in enhancing antibody responses has not yet been reported in chickens. In the current study, poly I:C and CpG induced similar IgY responses. In mammals, poly I:C-based adjuvants have been shown to enhance Ig responses against diverse antigens (21,25).
It is also important to note that none of the single TLR ligand adjuvants significantly enhanced HI titers. In fact, the only adjuvant to induce HI antibodies significantly was the poly I:C and CpG combination. However, based on the ELISA data, the amount of antigen-specific IgM and IgY antibodies was similar between CpG alone and poly I:C and CpG together. Taken together, these findings raise the possibility that the combination adjuvant enhances anti-HA antibodies and not necessarily total antibodies against AIV. A similar finding has been observed in murine T-cells, as a TLR ligand combination adjuvant used in an experimental HIV vaccine significantly enhanced the functional avidity and cytotoxic activity of the elicited CD8+effector T-cells, without resulting in a large increase in the number of tetramer positive CD8+T-cells (30). Nevertheless, the mechanisms mediating this enhanced antibody response are not yet known in chickens. One possible mechanism may be attributed in part to the synergistic IFN-γ response in chicken monocytes stimulated with poly I:C and CpG combinations (5). Since IFN-γ alone has been shown to enhance antibody responses when administered as a vaccine adjuvant in chickens (1), it is possible that a poly I:C and CpG adjuvant may result in the synergistic induction of IFN-γ upon acting directly on tissue resident macrophages and DCs soon after immunization. However, further studies are needed to assess the immunomodulating effects of TLR combinations in vivo.
Not only did the CpG and poly I:C combination adjuvant enhance antibody-mediated responses, but it also significantly enhanced cell-mediated immune responses. Vaccinating chickens with any of the double TLR ligand combination adjuvants induced robust proliferation in splenocytes upon stimulation with a recall antigen, which were significantly higher than responses with any of the single TLR ligands. Consistent with this, we observed a significant upregulation of IL-2 expression in all of the combination adjuvant groups. In mammals, it has previously been shown that poly I:C and CpG combinations act on antigen presenting cells and synergistically enhance the production of cytokines, which help induce and modulate T-cell responses (9). It has also been shown in mammals that poly I:C and CpG ODN when used alone induce cross-presentation and therefore facilitate the presentation of exogenous antigens to CD8+T-cells (3). As such, it is possible that when these two ligands are combined, an even greater increase in cross-presentation capabilities may occur. Nevertheless, the induction of T-cell responses is highly desired in an AIV vaccine, as effector T-cells have been shown to protect against lethal H5N1 AIV infections in chickens (16). Therefore, it is clear that a poly I:C and CpG ODN combination adjuvant addresses one of the limitations of existing inactivated whole virus AIV vaccines—the poor induction of T-cell responses (24).
In conclusion, we have shown that a poly I:C and CpG ODN combination adjuvant significantly enhances the immunogenicity of an inactivated AIV vaccine. Specifically, this combination adjuvant enhanced IgY and HI titers, as well as promoted the induction of a robust T-cell response. Future studies may be aimed at determining the efficacy of this vaccine adjuvant in protecting chickens against AIV infections.
Footnotes
Acknowledgments
M.S.P. is a recipient of an Ontario Graduate Scholarship in Science and Technology. This work was funded by Natural Sciences and Engineering Research Council of Canada, Poultry Industry Council, and OMAFRA. We would like to thank the staff at the Ontario Veterinary College Isolation Facility for the housing and care of the chickens.
Author Disclosure Statement
No competing financial interests exist.