
Abstract
Vaccination remains a useful means for the control of avian influenza viruses (AIV) in chickens. Current vaccines can protect chickens from morbidity and mortality. However, they do not eliminate virus shedding into the environment. Therefore, novel measures must be considered in order to enhance the immunogenicity of AIV vaccines, such as through the administration of immunostimulatory compounds. One such group of compounds is Toll-like receptor (TLR) ligands, such as bacterial flagellin, as well as synthetic lipopeptides such as Pam3CSK4. The objective of the present study was to assess the adjuvant potential of TLR2 and TLR5 ligands flagellin and Pam3 respectively. Chickens were vaccinated twice with an inactivated H4N6 AIV vaccine, 14 days apart. Antibody-mediated responses were assessed in sera and lacrimal secretions, while cell-mediated immune response was assessed by stimulating splenocytes from vaccinated chickens in vitro with the vaccine antigen. To evaluate vaccine efficacy, chickens were challenged with the H4N6 virus, and virus shedding was assessed on day 7 post-challenge. The results suggest that both ligands significantly enhanced antigen-specific IgY antibodies, while only the Pam3 adjuvant induced greater IgM and IgA antibody levels. Chickens receiving the flagellin adjuvant had significantly higher IgY responses, as well as significantly higher hemagglutination-inhibition antibody titers compared to the no adjuvant control. With respect to cell-mediated responses, splenocytes isolated from chickens that received either TLR ligand adjuvant proliferated in response to an in vitro stimulation with vaccine antigens. Lastly, chickens receiving vaccines containing either flagellin or Pam3 adjuvants were partially protected from an experimental AIV challenge and shed significantly less virus compared to controls. Future studies may be aimed at examining the efficacy of Pam3 and flagellin adjuvants for highly pathogenic AIV strains.
Introduction
A
TLRs are pattern recognition receptors that bind conserved structural motifs on pathogens termed “pathogen associated molecular patterns” (PAMPs) (1). A number of TLRs have been identified in chickens where they mediate responses to pathogens (3,20). One example is TLR2, which binds peptidoglycans, lipoteichoic acids, and other types of lipopeptides such as the synthetic Pam3CSK4, and TLR5, which binds bacterial flagellin (6,9,10). Following binding of their respective ligands, TLR2 and TLR5 promote cellular activation and the production of cytokines including proinflammatory interleukin (IL)-6, as well as those cytokines that help to initiate and modulate the adaptive immune response, including IL-4, IL-12, and interferon (IFN)-γ (23).
Owing in part to their immunostimulatory properties, TLR ligands have been shown to be efficacious vaccine adjuvants. In the case of TLR2 ligands, studies have demonstrated superior adjuvant potential in enhancing antibody titers against human serum albumin and keyhole limpet antigen (5,15). In addition, TLR5 ligands have also been used as vaccine adjuvants, including for AIV vaccines. However, the efficacy of this adjuvant in protecting against viral challenges remains to be determined (4). As a result, the objective of the present study was to determine the adjuvant potential of Pam3CSK4 and flagellin in the context of AIV vaccines, and to determine whether these adjuvants can protect chickens from AIV challenges. The results suggest that both ligands augmented antibody-mediated and cell-mediated immune responses toward the LPAI H4N6. Importantly, chickens receiving vaccines containing either TLR ligand adjuvant shed less virus after an experimental AIV challenge.
Materials and Methods
Chickens
Specific pathogen-free (SPF) chickens were procured from the Canadian Food Inspection Agency (Ottawa, Canada) and housed in the isolation facility of the Ontario Veterinary College, University of Guelph. This research was approved by the University of Guelph Animal Care Committee and complied with the guidelines of the Canadian Council on Animal Care.
TLR ligands
Pam3CSK4 and ultrapure flagellin isolated from Salmonella typhimurium were purchased from InvivoGen (San Diego, CA). All of the ligands were resuspended in sterile, endotoxin-free water.
AIV
The low pathogenic AIV A/Duck/Czech/56 (H4N6) was propagated in 11-day-old embryonated chicken eggs as described (12). To generate the vaccine, virus-rich allantoic fluid was mixed with formalin (final concentration 0.02%) and incubated for 72 h at 37°C with constant stirring. The allantoic fluid was subsequently ultracentrifuged at 23,000 rpm (SW 40 TI rotor; Beckman Coulter, Inc., Brea, CA) at 4°C for 1.5 h, and the supernatant was decanted. the pelleted virus was resuspended in HNE buffer (5.0 mM HEPES, 150 mM NaCl, and 0.1 mM EDTA, pH 7.4), layered on top of a 20% sucrose cushion, and ultracentrifuged again at 23,000 rpm at 4°C for 1.5 h. The supernatant was decanted, and the virus was resuspended in HNE buffer. The process was repeated again for a total of two washes. The virus was resuspended in HNE buffer and stored at −80°C. Total virus protein was measured using a Pierce BCA protein assay kit (Thermo Scientific, Rockford, IL) according to the manufacturer's recommended protocol. Inactivation was confirmed both by passaging the virus in Madin-Darby canine kidney (MDCK) cells as well as in 11-day-old embryonated chicken eggs. The integrity of virus hemagglutinin (HA) protein was confirmed by hemagglutination assay.
Vaccine formulations
Eight to 10 chickens per group were immunized subcutaneously on days 7 and 21 post-hatch with inactivated H4N6 (15 μg/bird) virus diluted in phosphate-buffered saline (PBS), with either a Pam3 (50 μg/bird) or flagellin (2 μg/bird) adjuvant. Control groups received solely H4N6 virus with no adjuvant (referred to as the H4N6 group) or PBS with no antigen (referred to as the PBS group).
Hemagglutination inhibition assay
Serial twofold dilutions of serum samples collected on day 28 post-primary immunization (d35 post-hatch) were incubated with four HA units of H4N6 virus in a 96-well V-bottom plate for 30 min at room temperature. Subsequently, a solution containing 0.5% washed chicken red blood cells (RBCs) was added and incubated for another 30 min at room temperature. The hemagglutination inhibition (HI) titer was determined as the reciprocal of the highest dilution of serum completely inhibiting hemagglutination of RBCs.
IgY, IgM, and IgA ELISA
To determine antibody levels against influenza viral antigens, sera and tears were collected from four to six chickens in each group on days 14 and 28 post-primary vaccination (days 21 and 35 post-hatch). Sera were assessed for antigen-specific IgY and IgM, while tears were assessed for antigen-specific IgA as described previously (12).
Cell proliferation assay
In order to examine cell-mediated immune responses, proliferative responses following stimulation with 1 μg/mL of inactivated whole virus was performed at day 10 post-secondary immunization (day 31 post-hatch) using Alamar Blue (AbD Serotec, Kidlington, United Kingdom) as previously described (8). Briefly, spleens from four birds of each vaccinated group and the control group were collected and single cell suspensions of spleen mononuclear cells were prepared as described previously (23). Cells were adjusted to 1×106 cells/mL in RPMI (Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum, 200 U/mL penicillin, 80 μg/mL streptomycin, and 25 mg gentamicin, and 100 μL was added to a 96-well cell culture plate. Cultures in triplicate were stimulated with 1 μg/mL of inactivated whole virus, concanavalin A (ConA; Sigma-Aldrich, Oakville, Canada) at 20 μg/mL and with medium (RPMI). Following the addition of antigen or control treatments, cells were incubated for 24 h at 41°C in a humidified 5% CO2 environment. Subsequently, 22 μL of Alamar Blue was added to all the wells, and after 24 h, the absorbance was recorded. The percent reduction of Alamar Blue is proportional to the degree of proliferation, and was calculated as previously described (8).
RNA extraction, cDNA synthesis, and real-time polymerase chain reaction
Total RNA was extracted from mononuclear splenocytes (1×107 cells/mL) at 24 h post-stimulation with 1 μg/mL of inactivated whole virus. RNA extraction and cDNA synthesis were performed as previously described (22). Real-time PCR using SYBR Green was performed on diluted cDNA using the LightCycler® 480 II (Roche Diagnostics GmbH, Mannheim, Germany) as previously described (22). Primers were synthesized by Sigma-Aldrich, and their specific sequences around found in Table 1. Relative expression of all target genes was calculated relative to the housekeeping gene β-actin using the LightCycler® 480 Software (Roche Diagnostics GmbH, Mannheim, Germany).
Viral challenge
On day 35 post-hatch, all remaining chickens were challenged with 1×106 50% tissue culture infectious dose (TCID50) of H4N6 virus via the oculo-nasal route. On day 7 post-challenge, cloacal swabs were collected to assess virus shedding by titration in MDCK cells as previously described (21).
Statistical analysis
For all data except for the HI assay, statistical significance between groups was calculated using Student's t-test, and were considered significant if p≤0.05 or p≤0.01. In the case of the HI results, Mann–Whitney's U-test was performed, and results were considered significant if p≤0.05 or p≤0.01.
Results
Pam3 and flagellin adjuvants enhance serum antibodies
To assess antibody-mediated responses, we quantified serum IgM and IgY levels, as well as IgA from lacrimal secretions (Fig. 1). On day 14 post-primary immunization (day 21 post-hatch), chickens vaccinated with a Pam3, but not flagellin, adjuvant demonstrated significantly enhanced antigen-specific IgM antibodies (p≤0.05). However, on day 28 post-primary immunization (day 35 post-hatch), IgM antibodies were not significantly different from the H4N6-only group. In the case of IgY, antigen-specific antibodies were significantly elevated when compared to the no-adjuvant control on day 14 post-primary immunization (day 21 post-hatch) for Pam3 (p≤0.01), and on day 28 post-primary immunization (day 35 post-hatch) for flagellin (p≤0.05). With respect to secretory IgA, none of the adjuvants promoted enhanced IgA responses by day 14 post-primary immunization (day 21 post-hatch). However, on day 28 post-primary immunization (day 35 post-hatch), chickens vaccinated with only the Pam3 adjuvant demonstrated significantly higher levels of IgA (p≤0.05).

IgM
In addition to performing ELISAs, antibody-mediated responses were further assessed by performing HI tests on sera collected on day 28 post-primary immunization (day 35 post-hatch) (Fig. 2). The results indicate that only chickens receiving the flagellin adjuvant had significantly higher HI titers, with a titer of ∼50 HI units (p≤0.05). Chickens receiving a Pam3 adjuvant did not demonstrate significantly increased HI titers compared to the no-adjuvant control.
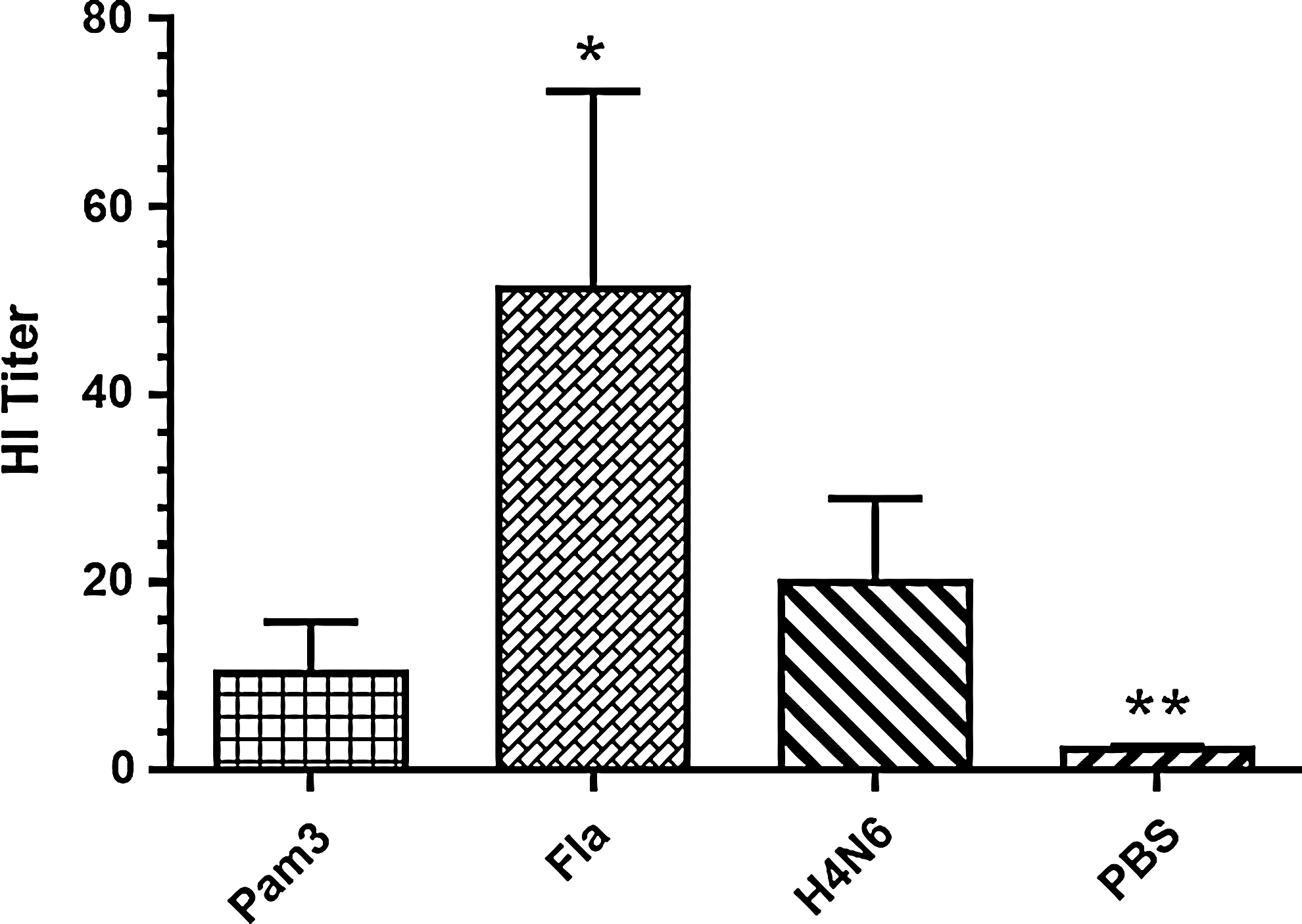
Hemagglutination inhibition titers of sera collected on day 28 post-primary immunization (day 35 post-hatch). Chickens were vaccinated twice with inactivated H4N6 virus in conjunction with either Pam3 or Fla. Control groups received either H4N6 antigen without any adjuvant (H4N6) or PBS with no antigen (PBS). Graphed values represent mean HI titer from four to six chickens/group±standard error. Statistical significance between treatment groups and the H4N6 control group was calculated using Mann–Whitney's U-test, and was considered statistically significant if p≤0.05 (*) and p≤0.01 (**).
Pam3 and flagellin adjuvants enhance cell-mediated immune responses
To gain insight into the cell-mediated immune response, splenocytes taken from vaccinated chickens were restimulated with inactivated H4N6 virus in vitro. Proliferation was assessed in response to the recall antigen (Fig. 3), and the results indicate that splenocytes isolated from chickens receiving either the Pam3 or flagellin adjuvant underwent robust proliferation. In contrast, proliferation of splenocytes isolated from chickens receiving no adjuvant was not statistically different from that of spleen cells from unimmunized controls.

Proliferative responses in splenocytes of vaccinated chickens on day 24 post-primary immunization (day 31 post-hatch). Mononuclear splenocytes isolated from vaccinated birds were stimulated with 1 μg/mL of inactivated H4N6 virus, and proliferation was assessed 48 h post-stimulation using Alamar Blue. Data represent mean percent reduction of Alamar Blue from four chickens/group±standard error. (**) denotes a statistically significant difference (p<0.01) between treatment groups and the H4N6 control group.
In addition to cell proliferation, gene expression was evaluated in splenocytes stimulated with the recall antigen (H4N6; Fig. 4). In the case of IFN-γ, transcripts were upregulated in splenocytes isolated from chickens receiving the flagellin adjuvant, but this was not statistically significant. With respect to IL-12, transcripts in the flagellin adjuvant group were significantly (p≤0.05) higher when compared with the no-adjuvant group. As for IL-4 and IL-13, transcripts were downregulated in the Pam3 and flagellin adjuvant groups. However, this was not statistically significant. In contrast, IL-2 transcripts were significantly upregulated in the Pam3 adjuvant group (p≤0.01) but not the flagellin adjuvant group. Lastly, transcripts for granzyme and perforin were upregulated in both the Pam3 and flagellin adjuvant groups, but this was not statistically significant.

Gene expression in splenocytes isolated from vaccinated chickens on day 24 post-primary immunization (day 31 post-hatch). Mononuclear splenocytes isolated from vaccinated birds were stimulated with 1 μg/mL of inactivated H4N6 virus and harvested for RNA extraction 24 h post-stimulation. Data represent mean relative expression levels of IFN-Y
As both adjuvants significantly enhanced the immunogenicity of the vaccine, we next assessed the efficacy of these vaccines in protecting against an experimental infection with live H4N6 virus in chickens (Fig. 5). Chickens receiving either the Pam3 or flagellin adjuvant shed less virus, as demonstrated by ∼4 log10 decrease in cloacal virus shedding (p≤0.01). In contrast, chickens receiving no adjuvant demonstrated no reduction in virus shedding when compared to the unvaccinated controls.

Cloacal virus shedding in vaccinated chickens on day 7 post-AIV challenge (day 42 post-hatch). All chickens were challenged with 1×106 TCID50 of H4N6 virus on day 28 post-primary vaccination. Graphed values represent the mean log10 TCID50/mL of swab fluid from four to six chickens/group±standard error. (**) denotes a statistically significant difference (p<0.01) between treatment groups and the H4N6 control group.
Discussion
Ligands for TLR2 and TLR5 promote cellular activation and the production of cytokines in a variety of immune cell subsets. Here, we report that the immunostimulatory properties of these ligands may be exploited to enhance the efficacy of AIV vaccines.
An important component of many vaccines is the induction of a robust antibody response. In this regard, TLR2 and TLR5 ligands such as Pam3 and lipoteichoic acid have been shown to enhance Ig responses to different vaccine antigens (20), a finding that we confirmed in the present study. It is important to note that although the Pam3 adjuvant significantly enhanced IgY and IgM antibody responses shortly after primary immunization, the same increase was not noticed after the secondary immunization. Instead, IgA antibodies were higher at 2 weeks post-secondary immunization in the Pam3-treated group but not in the flagellin-treated group or the no adjuvant control group. This class-switching to IgA may be attributed in part to the significant IL-4 response observed in splenocytes following treatment with Pam3 in vitro (23). IL-4 is typically associated with Th2-like responses, and has been shown to facilitate isotype switching to IgA (11). As secretory IgA is important for neutralizing influenza virus and reducing viral shedding from mucousal surfaces (16), this may contribute to the observed protection from viral challenge. Having said that, our results suggest that although a high IgA was observed, little serum IgM and IgY was noted and, as a result, little HI activity was detected in the Pam3-treated adjuvant group. In contrast, the flagellin adjuvant promoted a protective (>40 HI Units) HI titer, which correlated with significantly higher serum IgY levels. Although higher levels of IgM typically correlate with greater HI titers due to its pentameric structure, serum IgY may also prevent hemagglutination and is thought to help limit the spread of the virus if the virus overcomes the mucosal barriers (2,26).
An important component of many vaccines is the induction of cell-mediated immune responses. In the case of AIV, the importance of T-cell responses has been demonstrated through adoptive cell transfer of anti-influenza CD8+T-cells into naïve chickens, as this conferred immunity against a subsequent challenge with H9N2 virus (17). In this regard, the vaccine mediated induction of a robust T-cell response can help supplement the protective effects of antibodies. The results presented here suggest that both Pam3 and flagellin adjuvants enhanced cell-mediated responses, while a vaccine with no adjuvant induced little T-cell response. In addition to enhanced proliferative effects, T-cells isolated from Pam3 and flagellin vaccinated chickens demonstrated enhanced cytokine production, as well following in vitro AIV restimulation, especially with respect to higher IL-12 and IL-2 compared to no adjuvant controls. Furthermore, several other genes were upregulated but not significantly, which may be attributed to the smaller group sizes (n=4). The mechanism behind this enhanced T-cell response is not entirely clear, but it may be partly mediated by enhanced cross-presentation induced by the proinflammatory cytokines and type I IFN induced following TLR stimulation, a phenomenon that has been well described in mammals (18). In addition, it is also possible that the TLR ligands directly activate T-cells as well, as we have previously shown that chicken T-cells express several different TLRs and respond to Pam3 by upregulating IFN-γ transcripts (19). Nevertheless, it is clear that both adjuvants enhanced T-cell responses, which likely contributed in part to the protection observed following viral challenge.
In conclusion, we show here that both TLR2 and TLR5 ligands can serve as efficacious AIV vaccine adjuvants. Although neither adjuvant conferred total protection from AIV challenge, both adjuvants enhanced cell-mediated responses, while Pam3CSK4 enhanced secretory IgA and flagellin enhanced serum IgY titers. Future studies may be aimed at determining the efficacy of TLR2 and TLR5 adjuvants for HPAIV strains.
Footnotes
Acknowledgments
Michael St. Paul is a recipient of an Ontario Graduate Scholarship in Science and Technology. This work was funded by Natural Sciences and Engineering Research Council of Canada, Poultry Industry Council, and OMAF. We would like to thank the staff at the Ontario Veterinary College Isolation Facility for the housing and care of the chickens.
Author Disclosure Statement
No competing financial interests exist.