
Abstract
Alterations of the frequencies of recent thymic emigrants (RTEs) in blood have been demonstrated to be associated with human immunodeficiency virus-1 (HIV-1) disease progression and immune reconstitution following antiretroviral therapy. Flow cytometry is a technology that offers many advantages over T-cell receptor excision circles (TRECs) quantification to determine the RTE subset in blood. However, the gating strategies for identifying RTEs by flow cytometry have not been evaluated and compared in much detail. In the present study, we compared the frequencies and the senescence levels of RTEs in 54 HIV-1 seronegative controls (HC) and 70 HIV-1 seropositive (HIV-1+) subjects using two different gating strategies by flow cytometry. Our analysis indicated that the CD4+ RTE population determined by the expressing pattern CD45RO-CD31+ contained the terminal effector memory T-cell population after HIV-1 infection, which could significantly affect the further phenotypic and functional studies. Our data demonstrate the necessity of including an additional marker such as CCR7 to distinguish better CD4+RTE subset in HIV-1+ subjects.
Introduction
R
Recent advances in flow cytometry technology offer new tools for phenotypic and functional studies in the immune system. In 2002, Kimmig et al. first reported the use of CD31 (platelet endothelial cell adhesion molecule-1; PECAM-1) to distinguish RTEs in the peripheral blood of healthy humans by flow cytometry (11). There is a significantly higher sjTREC content in CD31+ CD4+ naïve T-cells, and the absolute numbers of CD31+ CD4+ naïve T-cells decline during aging and significantly correlate with the decline in TRECs (9,12,13). This finding not only furthers the progress of understanding RTE functions under healthy condition, but also facilitates the research on RTEs in patients with different diseases, including HIV-1 infection. However, we found that the majority of the studies on CD4+ RTEs in HIV-1 infection applied the conventional gating strategy that was used in healthy humans, in which CD4+ RTEs were defined by the expression pattern: CD45RO− (CD45RA+) and CD31+ (7,14,18,20,21). This approach might not be suitable for the identification of CD4+ RTEs in HIV-1 infected individuals due to the possible contamination of effector memory T-cells that also lack CD45RO expression. To exclude the effects due to these contaminants on the studies of CD4+ RTEs, it is indispensable to improve the gating strategy for CD4+ RTEs in HIV-1 infected subjects.
Here, for the first time, we compare the differences in CD4+ RTEs determined by two gating strategies in HC and HIV-1+ Caucasian subjects, which we term the modified method (as method 1) and the conventional method (as method 2). The frequencies of RTEs and the expression of CD57 on RTEs were analyzed, and the cause of differences were discussed in order to recommend a reliable protocol for determining CD4+ RTEs in HIV-1 infected patients.
Material and Methods
Study populations
We obtained peripheral blood samples from 70 HIV-1 seropositive (HIV-1+) Caucasian subjects in the HIV outpatient clinic of the Medical University Hanover. Of these, 39 were HIV-1+ ART receiving individuals, and 31 were HIV-1+ ART naïve individuals (mean age 47 years). We also enrolled 54 age-matched HIV-1 seronegetive controls (HC; mean age 49 years). Compared to the HC group, the HIV-1+ group presented a significantly lower CD4+ T-cell count (486.9±42.3 vs. 829.5±50.8, p<0.001) and a significantly lower CD4+ T-cell frequency (25.67±1.58 vs. 39.44±2.13, p<0.001). None of the participants presented coinfection of HCV or TB. The study was performed in strict accordance with the ethical principles as outlined in the WMA Declaration of Helsinki. All study subjects gave written, informed consent prior to their participation, and the protocol was approved by the local ethics committee (Vote of Medical University Hanover ethic committee No. 3150).
Isolation of peripheral blood mononuclear cells
PBMCs were isolated from fresh blood as described previously (15,16). Aliquots of 107 PBMCs were cryopreserved in heat-inactivated fetal calf serum (FCS) supplemented with 10% dimethyl sulfoxide (DMSO; Merck, Darmstadt, Germany).
Flow cytometry
PBMCs thawed from previously cryopreserved samples were analyzed on a FACS LSR II flow cytometer (BD Biosciences, Heidelberg, Germany) to detect RTEs in CD4+ T-cells. For methods 1 and 2, immunofluorescent monoclonal antibodies PEcy7-CD3, PerCP-CD19, PerCP-CD14, 7-AAD, APCcy7-CD4, PacOrange-CD8, PE-CD31, and FITC-CD57 were purchased from BD Pharmingen (San Diego, CA), and ECD-CD45RO was purchased from Beckman-Coulter (Krefeld, Germany). Exclusively for method 1, anti-CCR7 (150503) was purchased from R&D systems (Abingdon, United Kingdom). Indirect staining of CCR7 using Pacific blue-conjugated goat antibodies against mouse immunoglobulins (Invitrogen, Darmstadt, Germany) subsequently followed by washing and blocking of remaining goat-anti-mouse antibody using mouse serum (Dako, Glostrup, Denmark) preceded the staining with directly labeled antibodies. All the above-mentioned monoclonal antibodies were used in optimized concentrations according to the instructions provided by the manufacturers. Analysis of flow cytometry data was carried out with FlowJo software (v8.8.6, Tree Star).
For the modified gating strategy (method 1), low forward and low side scatter lymphocytes were first gated and then continued to a second FACS plot to discriminate live CD3+ T-cells with no 7-AAD, CD14, and CD19 expressions. Following gating on CD4+ events, the naïve subset was further identified as the CD45RO− CCR7+ population. Lastly, RTEs were scrutinized for their expression of CD31 (Fig. 1A). For the conventional gating strategy (method 2), RTEs were analyzed directly as CD45RO− CD31+ cells within CD4+ CD3+ T-lymphocytes (Fig. 1B).
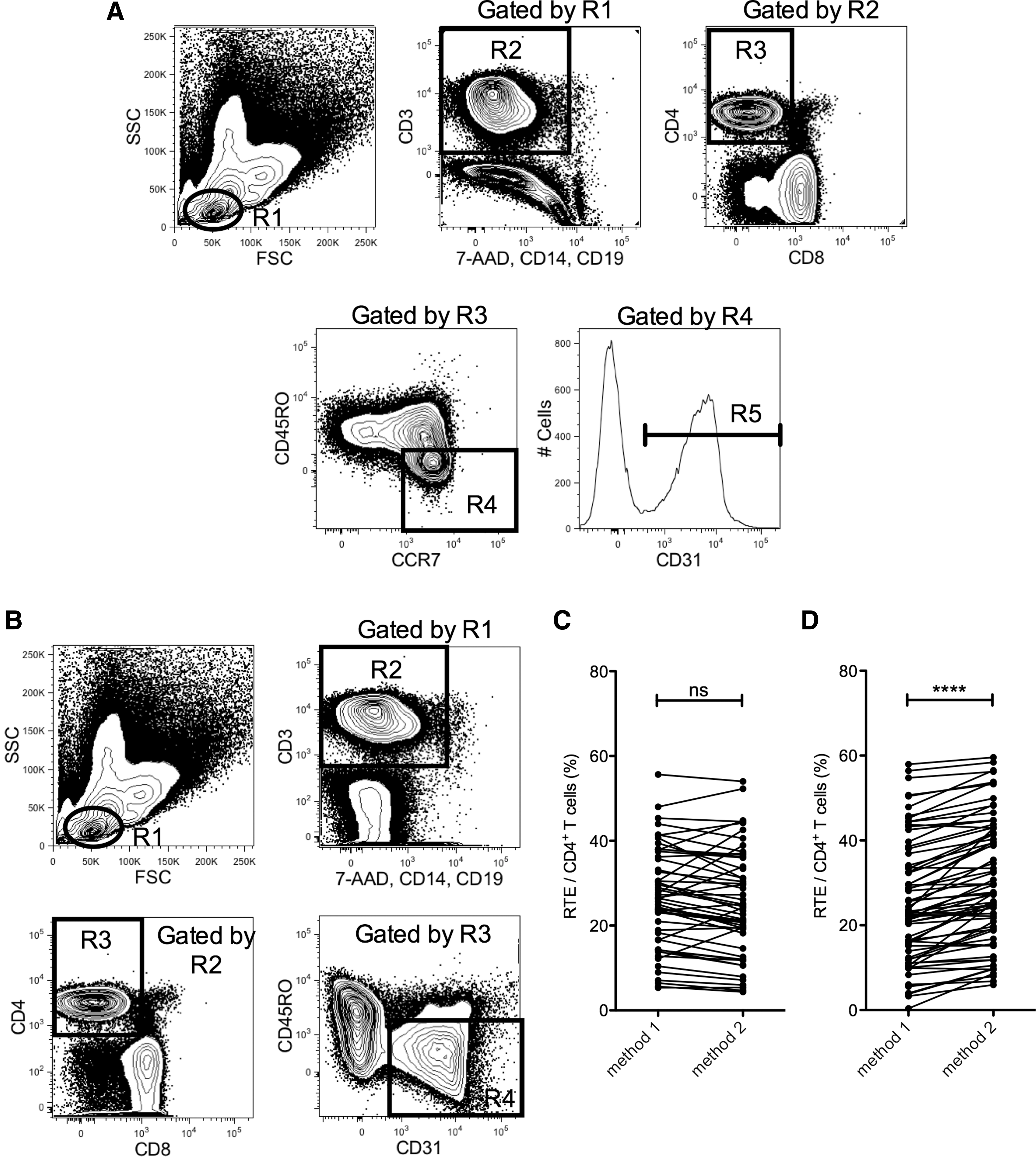
Comparison of recent thymic emigrants (RTE) frequencies determined by method 1 with method 2.
Statistical analysis
GraphPad Prism (v5.0) software was used for statistical evaluation of data. The two different methods for determining the RTE frequencies were compared using a two-tailed paired t-test. A two-tailed unpaired t-test was applied when comparing between two study groups. p-Values of <0.05 were considered significant.
Results
Comparison of RTE frequencies determined by method 1 with method 2
The CD4+ RTE frequencies in HC and HIV-1+ groups were compared using methods 1 and 2 as described above. As shown in Figure 1C, there was no significant difference between the CD4+ RTE frequencies determined by the two methods in HC. However, we found that there was a significant difference between the CD4+ RTE frequencies determined by the two methods in the HIV-1+ group (Fig. 1D; p<0.0001). In HIV-1+ subjects, we observed that the majority of the CD4+ RTE frequencies measured by method 2 were higher than those determined by method 1 (30.1±1.7% vs. 25.6±1.7%).
Expansion of CD4+ TEMRA cells in HIV-1 infection and its effect on the measurement of RTE frequencies
To understand the reasons that cause the significant difference between the CD4+ RTE frequencies determined by the two methods in the HIV-1+ subjects, we conducted a comprehensive phenotype analysis on the CD4+ T-cells. As shown in the representative contour plot from method 1 (Fig. 2A), CD4+ T-cells from HC comprised only low levels of the CCR7− CD45RO− population, which is also known as CD45RA+ effector memory T-cells (TEMRA). After HIV-1 infection, TEMRA populations were considerable increased in most of the patients (Fig. 2B). In addition, about 16% (11/70) of them had TEMRA/CD4+ T-cells >10% (Fig. 2C and D). The mean frequency of TEMRA in CD4+ T-cells was significantly higher in HIV-1+ group than in HC (Fig. 2D, 7.0±0.8% vs. 2.0±0.2%, p<0.0001).
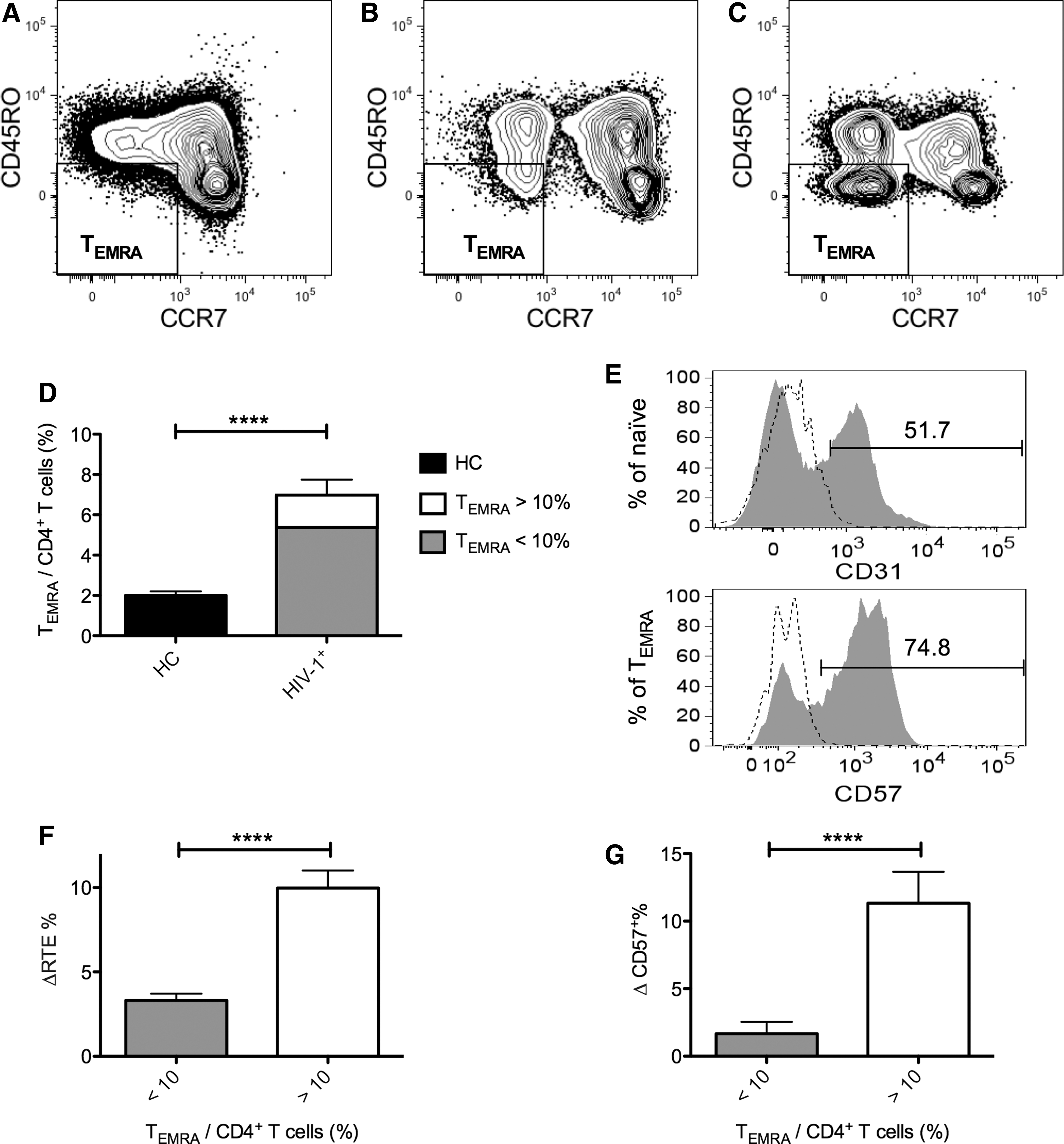
CD4+ TEMRA cells expression patterns in HC and HIV-1+, and the effect of TEMRA cells on the different gating methods for RTEs. After the identification of CD4+ T-cells using the same gating strategy mentioned previously, cells were then gated on CD45RO− and CCR7− expression to identify the TEMRA subset in
Because of the expansion of the TEMRA population and the lack of distinguishing TEMRA from RTEs by the conventional method, we further compared the disparity of the CD4+ RTE frequencies determined by the two methods (ΔRTE%=[RTE frequencies in CD4+ T-cells determined by method 2]−[RTE frequencies in CD4+ T-cells determined by method 1]) in HIV-1+ subjects. Representative histograms of CD31 and CD57 flow cytometry analysis of the naïve and TEMRA CD4+ subsets in HIV-1+ patients are shown in Figure 2E. Our results showed that the mean ΔRTE% of HIV-1+ individuals with TEMRA/CD4+ T-cells >10% was significantly higher than that from HIV-1+ individuals with TEMRA/CD4+ T-cells <10% (Fig. 2F, 10.0±1.0% vs. 3.3±0.4%, p<0.0001). To understand whether this significant difference has an impact on the following phenotypic study on RTEs, we analyzed the difference of CD57 (senescence marker) expressions on RTEs determined by the two methods (ΔCD57+%=[CD57+ frequencies in RTEs determined by method 2]−[CD57+ frequencies in RTEs determined by method 1]) in HIV-1+ subjects. The data showed that the mean ΔCD57+% of the HIV-1+ individuals with TEMRA/CD4+ T-cells >10% was significantly higher than that from HIV-1+ individuals with TEMRA/CD4+ T-cells <10% (Fig. 2G, 11.3±2.3% vs. 1.7±0.9%, p<0.0001).
Discussion
Increasing numbers of researches have focused on the role of CD4+ RTEs in HIV infection by using flow cytometry (10), but the majority of these studies have applied the conventional gating strategy to identify CD4+ RTEs, which might contain virus-specific TEMRA cell contamination. Here, we conducted a comparison on the RTEs determined by the conventional method (method 2) and the proposed modified method (method 1). Our results indicated that in HIV-1 infection the CD4+ RTE population determined by method 2 contained the TEMRA population, which could significantly affect the further phenotypic analysis. In order to exclude the TEMRA cell contamination in HIV-1 infection, method 1 is recommended for determining RTE population in HIV-1+ subjects.
CD31 has recently been designated as a marker to identify RTEs in the CD4+ naïve T-cells (12), which has greatly facilitated further phenotypic and functional studies of RTEs in the peripheral blood. Because CD31 is not exclusively expressed by CD4+ naïve T-cells but also by memory subsets, a careful identification of CD4+ naïve T-cells before using CD31 for determining RTEs becomes very important. It has been demonstrated that three markers, including CD45RO, CCR7, and CD62L, are sufficient to identify naïve T-cells (defined as CD45RO− CCR7+ CD62L+) and to exclude memory T-cell contaminants (4). In some other reports, only two markers are sufficient for identification of naïve T-cells. Since CD62L expression is easily lost with the freeze and thaw procedure (17), it is important to note that CD62L staining is not reliable when cryopreserved cells are used, especially for large-scale analysis that is not easy to carry out all assays on freshly isolated cells. Therefore, naïve T-cells were determined by CD45RO–CCR7+ expression patterns in method 1. Other T-cell differentiation markers for the naïve subset, such as CD27 and CD127, can also be optionally included. However, our results showed that >95% of the CD4+ naïve T-cells (defined as CD45RO–CCR7+) are CD27hi and CD127hi (data not shown), confirming that CD45RO and CCR7 markers are sufficient for the identification of CD4+ naïve T-cells.
After naïve T-cells encounter antigens, they switch from the CD45RA to CD45RO isoform expression. Memory T-cells, according to the differential CCR7 expression, can be divided into central memory T-cells (TCM cells; CD45RO+ CCR7+), which are able to migrate to lymph nodes and display a high proliferation potential, and effector memory T-cells (TEMRO cells; CD45RO+ CCR7), which exert effector functions in peripheral tissues (19). In addition, a proportion of TEMRO cells specific for some persistent viruses, such as HIV-1, human cytomegalovirus (HCMV), and Epstein–Barr virus (EBV), can revert to the RA isoform of CD45 (TEMRA cells; CD45RO− CCR7−) after the infection (2,16,22). Our data showed that about 16% (11/70) of the HIV-1+ subjects had TEMRA/CD4+ T-cells >10%. By using the conventional gating strategy in which CD4+ naïve T-cells are only defined by CD45RO− (or CD45RA+), the CD45RO− TEMRA cells cannot be separated from RTEs in the HIV-1+ subjects. Therefore, the percentage of RTE in CD4+ T-cells was significantly higher by applying the conventional method than by using the modified method.
Our previous study has shown CD57 expression on HIV-1 specific T-cells with both the TEMRO and TEMRA phenotype (16). CD57 serves as a marker for replicative senescence and indicates a history of numerous cell divisions as shown by shorter telomere lengths (1). Moreover, CD57 expression is associated with terminal differentiation and altered functional capacities in T-cells as well as in NK cells (3,8). In the present study, the expression levels of CD57 on RTEs in the HC group determined by the two methods were both below 0.5% (data not shown). In contrast, the expression level of CD57 on CD4+ RTEs determined by the conventional method was significantly higher than that determined by the modified method in HIV-1+ subjects with TEMRA/CD4+ T-cells >10%. These data indicate that the CD4+ RTEs determined by the conventional method includes effector memory T-cell contaminants, which can significantly affect any subsequent phenotypic analysis.
Conclusion
We demonstrate that the conventional gating strategy for CD4+ RTEs in HC is not suitable for identifying CD4+ RTEs in HIV-1+ subjects due to the contamination of TEMRA cells. Therefore, the modified method that we propose here can offer a more reliable protocol for the determination of CD4+ RTEs in HIV-1 infected patients.
Footnotes
Acknowledgments
We gratefully acknowledge all the blood donors. I.L. was supported by the Hannover Biomedical Research School (HBRS) and the PhD program “Molecular Medicine,” Germany. This work was supported by grants from the Bundesministerium für Bildung und Forschung, Stiftung Zukunfts- und Innovationsfonds Niedersachsen, the European AIDS Treatment Network (D.M.O. and R.E.S.), and DZIF 8000 101–3 (R.E.S. and D.M.O.).
Author Disclosure Statement
No competing financial interests exist.