
Abstract
The 2009 pandemic H1N1 mainly affected adolescents and children, and most of the elderly in Japan escaped clinical illness. To clarify the role of humoral immunity in the infection, the time kinetics of hemagglutination inhibition (HI), neutralization (NT), and IgG subclass antibody response directed against influenza A(H1N1)pdm2009 were analyzed in three consecutive specimens obtained from 51 young adults and children (group 1) who contracted pandemic influenza and from 74 pediatric clinic employees (group 2) inoculated with pandemic monovalent vaccine. In group 1 patients, 6 and 30 patients had lower HI and NT antibody in the acute phase respectively. Thereafter, HI and NT antibody titers increased fourfold or more in 50 patients with peak response in the third specimens obtained four weeks after the onset. IgG1 in 45 patients, IgG3 in 18 patients, and IgG4 in 29 patients showed elevated responses. Forty (54%) and 70 (95%) subjects in group 2 had positive HI and NT antibodies in the prevaccination samples, with increased antibody responses in the follow-up peaking in the second specimens. Forty of those vaccinated had increased IgG1 responses peaking in the third specimens, whereas elevated IgG3 was observed in 22 recipients with the highest level in the second samples. IgG4 did not show any increase in subjects in group 2. A few participants showed an IgG2 response in both groups. An immunologically naive population contracted influenza with apparent clinical symptoms. However, already primed subjects through subclinical infection elicited the unique pattern of IgG subclass responses by vaccination, which differed from those of naive populations.
Introduction
I
Although many authors describe an important role of specific cell-mediated immunity (CMI) in protection (10), actual studies of specific CMI in humans is quite difficult in the clinical setting mainly because of sample volume constraints, especially in those with a primary immune response to influenza who are generally small children and infants. Also, it is virtually impossible to handle short-lived samples such as lymphocytes in the clinic during the heavy workload created by a pandemic. However, it is established that induced subclass IgG composition specific to antigen reflects a balance of Th1/Th2 activation in animal models (14), suggesting that it is worthwhile trying to trace the imprint of Th1/Th2 activities occurring in subjects with pandemic influenza or post-vaccination by studying IgG subclass responses to pandemic influenza.
In linking data obtained in humans with that from mice, we need to consider the confusing historical background regarding IgG subclass nomenclature. Human IgG subclasses have been identified according to the time of their discovery, but for the mouse, IgG subclasses reflect their electrophoretic mobility (22). Functional difference between mouse IgG subclasses and their similarly named human fractions are not yet fully documented. In mice, these responses correlate with IgG2a, IgG2b, and IgG3, regulated by the production of IFN-γ (21), while in humans such interrelationships are not yet well defined. IgG1 is the most abundant subtype, making up more than 50% of total IgG, and IgG4 is the smallest component. Human IgG1 reflects not only Th1 cytokine response but also Th2 cytokine activation. IgG4 subclass switch depend on IL-4 and IL-13, which are considered the Th2 response (2,18).
Primary antibody responses require T-cell help through functionally different Th1 and Th2 cytokines secreted by antigen-specific CD4+ T-lymphocytes. Therefore, the analysis for IgG subclass responses after infection or vaccination may provide additional information in terms of CD4+ T-cell effector functions, which might be supposed to reflect the balance of Th1 and Th2 activation provided by the antigenic stimulus of the novel influenza pandemic. Thus, our aim in the present study was twofold: first, to clarify the role of humoral immunity in the infection of 2009 pandemic; second, to evaluate the induced subclass IgG composition. To achieve these goals, we determined the time kinetics of hemagglutination inhibition (HI), neutralization (NT), and IgG subclass antibody response directed against influenza A/California/07/2009(H1N1) in patients with natural infection and subjects immunized with pandemic monovalent vaccine.
Materials and Methods
Study subjects
The study groups consisted of 51 healthy young adults and children (24 females) with a mean age of 7.6 years (range 0.5–25 years), who contracted pandemic influenza caused by influenza A(H1N1)pdm2009, and 74 pediatric clinic employees (63 females) with a mean age of 38.5 years (range 19–72 years) inoculated with one dose of pandemic vaccine. The vaccine formulation used in this study was monovalent HA split product of influenza A(H1N1)pdm09 virus made by Japanese vaccine manufacturers. Diagnosis of influenza was made by rapid test (ImmunoAceFlu®; Tauns, Izunokuni, Japan) followed by virus isolation, polymerase chain reaction (PCR), and seroconversion with HI and NT antibody determination. A detailed history was taken from pandemic vaccine recipients as to symptoms suggesting influenza such as fever, cough, rhinorrhea, headache, diarrhea, muscle pain, and arthralgia. None had had any of these symptoms during the past year. Three consecutive peripheral blood specimens (acute phases, 2 weeks and 4 weeks after the onset of illness in patients with influenza and both prior to and 2 and 4 weeks after vaccination in recipients) were obtained. Blood sampling in patients with natural infection was based on the first day of fever designated as day 0, and the first specimens were collected after 2±2 days of the acute illness. Patients were recruited from September 16 through October 20 followed by an additional two samplings. Regarding pandemic vaccine recipients, the first specimens were obtained on the day of immunization, which extended from October 26 through November 21. Two late samplings were included. This study was approved by the ethical committee of Sapporo Medical University and by the separate institutions of the authors. Informed consent was obtained from the subjects after full explanation, and consent was also obtained from all children aged 6 years or more.
Influenza virus and antigen preparation used for antibody assay
A/California/07/2009 virus purified whole virion was prepared by means of sucrose density gradient centrifugation for HI test. HA split vaccine bulk material of influenza A(H1N1)pdm2009 was used to determine IgG subclass antibody titer, which was adjusted to 333 ng/mL of HA protein in phosphate-buffered saline (PBS) as antigen.
Determination of HI and NT antibody specific to influenza A/California/07/2009 virus
The serum samples were stored at −35°C until testing. The HI test was performed by a standard microtiter assay with human type O erythrocytes after removal of nonspecific inhibitors with receptor-destroying enzyme and of cold agglutinins by hemadsorption at 4°C. All specimens were tested on the same day using identical reagents.
Determination of NT antibody titers was carried out by the microneutralization (MN) assay (17). In brief, serially diluted sera (1:10 to 1:1280) in Dulbecco's minimal essential medium (DMEM) were prepared in a 96-well microplate. An equal volume of virus fluid (50 FFU/25 μL) was added and incubated at 37°C for 60 min in a 5% CO2 incubator. As a positive control, viruses were mixed with an equal volume of DMEM without serum. After incubation, 50 μL of the NT mixture was inoculated onto a monolayer of MDCK cells in a 96-well microplate. After incubation for 16–20 h, cells were fixed with absolute ethanol and reacted with mouse monoclonal antibody against influenza A virus nucleoprotein (NP; clone C43) (17), followed by incubation with horseradish peroxidase (HRP)-conjugated goat anti-mouse IgG antibody (A2304; Sigma). Cells expressing viral antigens were visualized by 3,3′-diaminobenzidine tetrahydrochloride hydrate (D5637; Sigma). The MN titer was indicated by the reciprocal value of the maximum dilution at which the number of foci showed 50% reduction compared with the positive control.
In both the HI and NT tests, positive cutoff was set at 1:10. Thus, an antibody level <1:10 was designated as negative. The criterion for seroconversion was defined as elevation of antibody level by fourfold or more. To investigate antibody responses in detail, we classified them into three categories: no elevation; antibody level unchanged or less than the first specimen; twofold increase and fourfold ore more elevation.
Determination of IgG subclass antibodies
We performed quantitative enzyme immunoassay (EIA) to detect IgG subclass antibodies against influenza A/California/07/2009 strain. Wells of a 96-well plate were coated with virus antigen, 33 ng/100 μL of split vaccine materials calibrated to the amount of HA protein for 16 h at room temperature. After four washes with PBS containing 0.05%Tween 20, wells were coated with Blocking One (Nacalai Tesque) for 45 min at room temperature. Serial twofold dilutions of serum samples were incubated in 100 μL/well for 60 min at room temperature, starting at 1:200 for IgG1, IgG2, and IgG4, and at 1:20 for IgG3. Secondary antibodies were added at 1:3,000 dilution for anti-human IgG1, at 1:1,000 dilution for IgG2, at 1:2,000 dilution for IgG3, and 1:2,000 dilution for IgG4 (Invitrogen). The plate was incubated for 60 min at room temperature. The wells were washed, and 100 μL of enzyme substrate was added, which was prepared with 40 mg of O-phenylenediamine (Wako Junyaku) in 100 mL of Na-citrate buffer, with 20 μL of 30% H2O2 added just before use. After 20 min incubation, 50 μL of 4N-H2SO4 was added, and optical density was measured at 450/630 nm. EIA antibody titers were expressed as the reciprocal dilution of 100×2 n that gave twofold value of optical density in the negative control well for each serum sample, derived from regression curve analysis with three dilution point plotting.
Statistical analysis
Consistency in HI, NT, and IgG subclass antibody directed against A/California/07/2009 virus at three points after onset of influenza and of pre- and post-vaccination was examined by using a nonparametric repeated-measures analysis of variance (Friedman's test) followed by Dunn's test as post-test employing StatFlex software v6. For calculation of geometric mean titer (GMT) estimates, a titer of <10 was assigned a value of 5, and a titer of ≥8 in HI, NT test, and that of 9 in IgG subclass determination were assigned a value of 9 and 10 respectively.
Results
Time course characteristics of the antibody response specific to pandemic H1N1 2009 in naturally infected subjects was profiled (Fig. 1). GMTs determined in the first, second and third specimens with 95% confidence intervals (Cis) were 5.4 [5.1, 5.8], 12.4 [8.6, 17.8], 120.4 [94.1, 154.1] in HI, and 8.7 [7.4, 10.2], 44.7 [28.1, 71.1], 407.9 [312.6, 532.1] in NT, respectively. Statistically significant antibody elevations were observed (Friedman's test, p<0.0001), and Dunn's test confirmed that peak antibody responses occurred in the third specimens (p<0.001), implying that a primary immune response accompanied natural infection. More specifically, 6 of 51 patients had low-level HI, and 30 of the 51 low-level NT antibody in the first specimens, and 50 showed a fourfold increase or more in HI and NT antibody titers in the follow-up. The remaining patient had HI antibody of 1:10 in the first specimen followed by elevation to 1:20 in the second specimen. They also had NT antibody of 1:40 in the first specimen and did not show any increase. Virus isolation and PCR were positive in this patient.
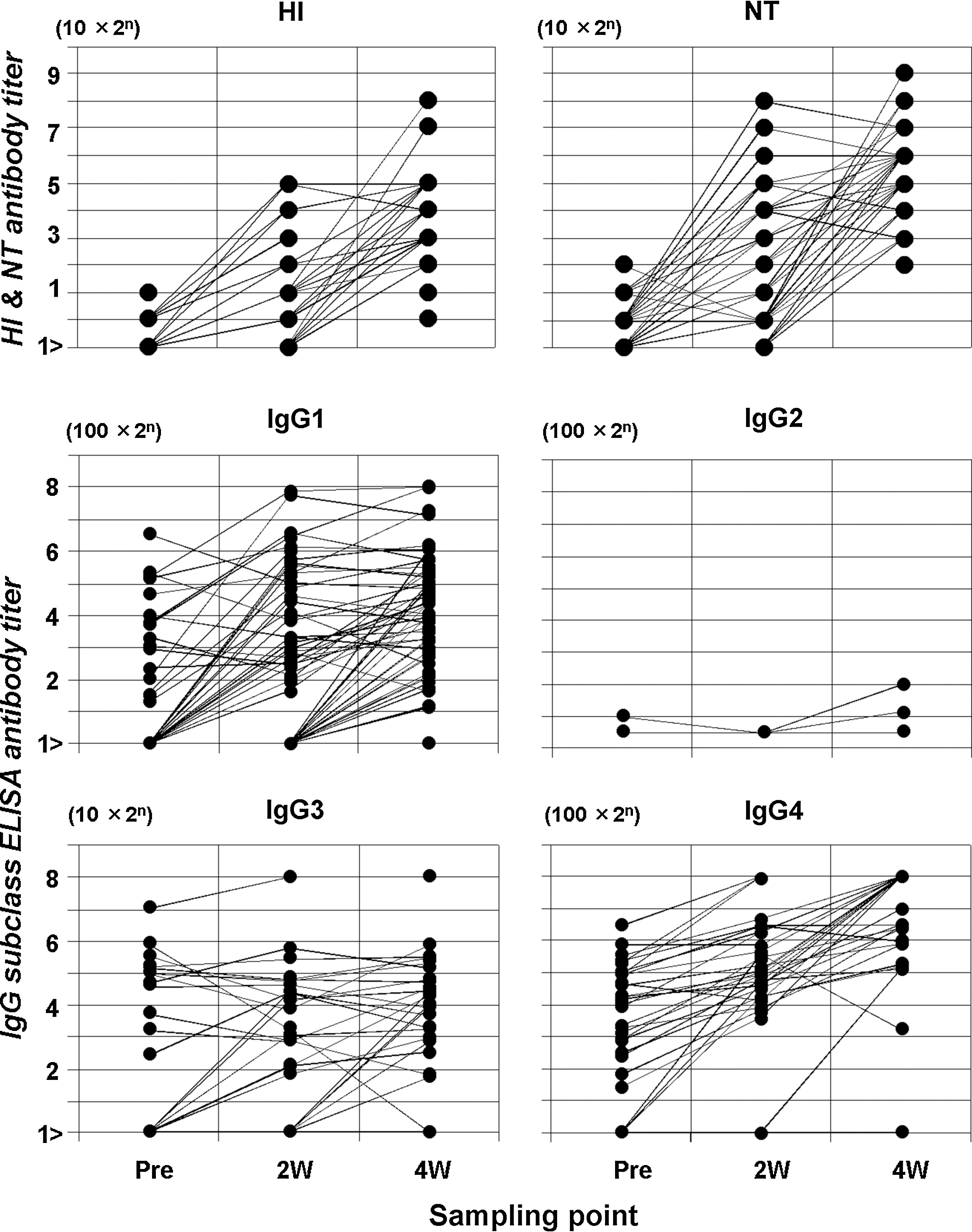
Hemagglutination inhibition, neutralization, and IgG subclass antibody titers specific to A/California/07/2009(H1N1) in group 1 patients with natural infection. Black dot shows antibody titer. Abscissa in sampling point after onset of illness. Ordinate in serum dilution.
IgG subclass antibody GMTs in three consecutive specimens obtained from patients were: for IgG1, 273.1 [198.0, 376.6], 737.3 [475.5, 1143.0], 1788.2 [1270.0, 2517.8]; for IgG3, 33.4 [22.4, 49.8], 39.8 [27.1, 58.3], 50.2 [33.7, 74.9]; and for IgG4, 476.6 [322.1, 705.2], 914.2 [548.8, 1522.9], 1780.1 [927.3, 3417.2]. IgG1, IgG3, and IgG4 antibody levels rose significantly (p<0.0001 for IgG1; p=0.0045 for IgG3; p<0.0001 for IgG4) and peaked in the third specimen (p<0.05 for IgG1; p<0.002 for IgG3; p<0.001 for IgG4). Only three cases developed IgG2 antibody at a very low level. Among 51 patients, initial specimens had some level of IgG1 (16 patients), IgG3 (15), and IgG4 (25) antibody activity, and 9, 2, and 21 of the above-mentioned cases exhibited a subsequent rise of fourfold or more. Twofold elevations were also observed in two cases for IgG1, three for IgG3, and four for IgG4. Five cases for IgG1 and 10 for IgG3 did not show any increase. IgG1 in 34 cases, IgG3 in 13 cases, and IgG4 in four cases were negative in the first samples but eventually manifested a fourfold or more rise in 32, 12, and 4 patients. The remaining two patients for IgG1 and one for IgG3 showed a twofold increase. No elevated response for IgG1, IgG2, IgG3, or IgG4 was observed in 1, 48, 23, and 22 subjects, respectively, throughout natural infection.
GMTs in HI and NT in pre- and post-vaccination serial specimens were 11.3 [8.9, 14.4], 70.1 [50.8, 96.9], 62.8 [45.4, 86.7], and 38.1 [28.2, 51.5], 329.0 [231.6, 467.4], 304.8 [215.6, 430.9], respectively (Fig. 2). Post-vaccination antibody titers were significantly higher than prevaccination (p<0.0001) titers for both HI and NT. Antibody titers peaked in the second specimens, then declined in the third (p<0.001). Of those vaccinated, 11 of 74 exhibited prevaccination HI antibody of ≥1:40, three of whom subsequently manifested fourfold or more increases. Four participants showed only a twofold rise thereafter, and the remaining four showed no elevation. Twenty-nine had prevaccination HI antibody of 1:10 to 1:20, and subsequent antibody increases were observed in all but two cases. Among these, 24 had fourfold or more increases, and three showed a twofold increase. Thirty-four had no HI antibody in prevaccination samples, and 23 of those showed fourfold elevation or more, 11 a twofold HI antibody rise, and two had no HI antibody response. Regarding NT antibody, 42 had a prevaccination titer of ≥1:40, and 29 of those showed fourfold or more increases thereafter. The remaining nine revealed a twofold rise, and four did not exhibit any increased antibody response. Twenty-eight had positive NT antibodies of 1:10 to 1:20 in prevaccination samples, and 22 of those developed antibody rises of fourfold or more, four showed twofold increases, and two revealed no elevation. The remaining four did not have NT antibody before vaccination but developed a fourfold or more antibody response.
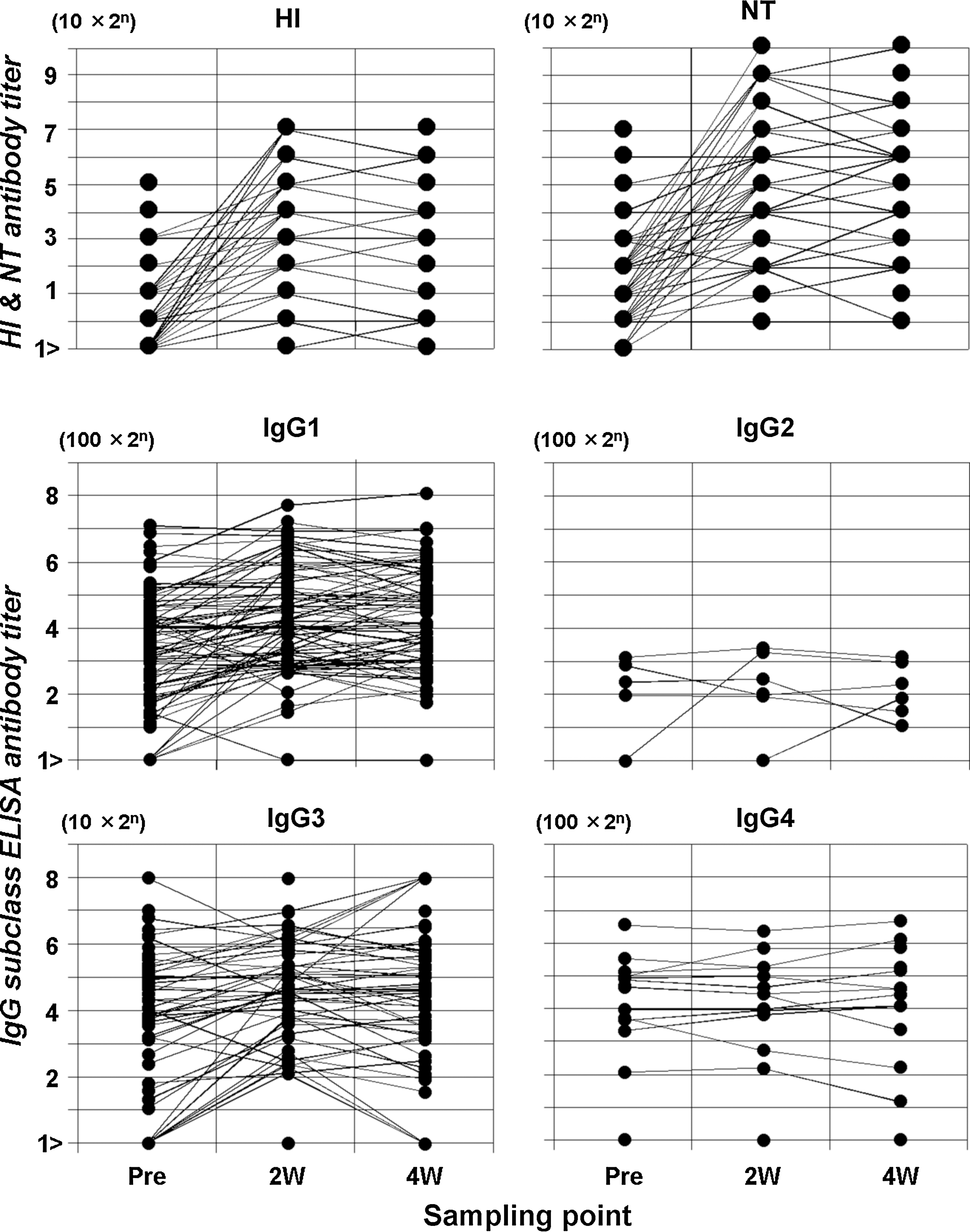
Hemagglutination inhibition, neutralization, and IgG subclass antibody titers specific to A/California/07/2009(H1N1) in group 2 vaccine recipients. Black dot represents antibody titer. Abscissa in sampling point before and after vaccination. Ordinate in serum dilution.
Regarding IgG subclass antibody developments in vaccine recipients, GMTs of IgG1 in consecutive three specimens were 1085.4 [827.4, 1423.7], 1970.0 [1541.7, 2517.3], 2020.2 [1587.3, 2571.1]; those of IgG3 were 88.4 [56.5, 138.5], 134.2 [88.5, 203.5], 128.8 [81.2, 204.6]; and those of IgG4 were 172.3 [129.0, 230.1], 172.2 [128.9, 230.0], 171.5 [127.7, 230.4]. IgG2 was found in only six subjects. IgG1 and IgG3 antibodies levels were significantly elevated (p<0.0001 for IgG1; p=0.0001 for IgG3), and the peak IgG1 responses occurred in the third samples (p<0.05), whereas the levels of IgG3 titers peaked in the second samples (p<0.001). IgG1 was positive in prevaccination specimens in 70 subjects, and 19 and 17 of those showed fourfold or more and twofold increases thereafter, respectively. Four recipients had no IgG1 antibody in the first specimens and showed a fourfold or more rise in the follow-up. No IgG1 antibody rise was observed in 34 patients who had positive antibody prevaccination. In terms of IgG3 in vaccine recipients, 49 had positive prevaccination samples, and 5 and 10 of those showed fourfold or more and twofold rises thereafter. Seven recipients had no IgG3 in the first specimens and acquired IgG3 fourfold or more. Thirty-four who had positive IgG3 in the first samples did not show any elevation thereafter. Eighteen recipients did not have IgG3 in any of three serial specimens. Only 13 recipients had positive IgG4 in prevaccination samples and showed no elevation in the follow-up. Sixty-one subjects did not have any IgG4 antibody in serial three specimens.
Discussion
Our main purpose in the present study was to determine the time kinetics of the development of HI and NT antibody to influenza A(H1N1)pdm2009, in order to clarify the role of the humoral immune response to newly appearing influenza subtypes. We observed a fourfold increase in HI antibody titers with the peak titer appearing 4 weeks after infection in almost all patients with natural infection, suggesting that a primary immune response was elicited during the course of pandemic influenza. A fourfold rise in antibody response does not necessarily imply a primary immune response if it occurred after contracting seasonal influenza. However, influenza A(H1N1)pdm2009 was a newly emerged reassortant subtype influenza virus, which did not strongly cross-react to any other subtypes, including even A/USSR/90/77 (H1N1) (1). Thus, it is likely that a fourfold elevation in antibody response to this particular strain strongly suggests a primary infection. A distinctive and unique point of the present study was the detailed time kinetic observation of antibody development using three consecutive specimens obtained from subjects with natural infection and those who were vaccinated.
Regarding antibody development in pandemic vaccine recipients, 40 (54%) and 70 (95%) of 74 subjects had positive HI and NT antibodies, respectively, in prevaccination specimens. These antibodies were assumed to be obtained through asymptomatic infection, which is supported by the fact that all pediatric employees enrolled in this study were free of symptoms of influenza during the pandemic in Japan. The percentage of positive HI is similar to other reports (3,4). However, the number of subjects with positive NT antibodies was remarkable. At least in part, this is because these subjects were employees of pediatric clinics where they were exposed to massive amounts of live viruses of pandemic influenza during a 6 month period. A time kinetic study showed that their antibody levels peaked in the second specimens, implying that the secondary immune responses were elicited by the subsequent pandemic vaccination.
The discrepancy in kinetic pattern of antibody development between HI and NT may depend on a fundamental difference in immunologic characteristics between the two antibodies. The mechanism of action of underlying antibodies in the interaction between antibody and cellular components in the human immune system is poorly understood. Thus, we need to interpret these data cautiously, and further characterization of the antibodies would be valuable. In this respect, we need to remember that influenza A(H1N1)pdm2009 virus was a natural reassortant strain with North America and Eurasian swine lineages originated from avian or human viruses (9).
With regard to this point, our study provides another set of unique data concerning specific IgG subclass responses to influenza A(H1N1)pdm 2009. Our observations are the first data to provide serial three point time kinetics of the development of IgG subclass antibodies specific to influenza A(H1N1)pdm 2009. IgG1, IgG3, and IgG4 antibodies were developed in natural infections, and peak responses were found in the third specimen. The majority (88%) of subjects with natural infection experienced IgG1 antibody elevation, whereas 18 and 29 showed a rise in IgG3 and IgG4, respectively. In vaccine recipients, peak IgG1 responses were found in the third specimens, whereas those of IgG 3 were found in the second samples, which may be important in explaining the findings that the peak responses in HI and NT appeared in the second specimens (25). IgG4 did not change in the follow-up, while IgG2 did not develop in most subjects with either natural infection or vaccinations.
Developmental patterns or characteristics of these specific IgG subclass antibodies in humans are so far quite distinct from those observed in mice (11,15). In mice, IgG1 is closely related to the activities of HI, but that is no longer the case in the humans in our study. However, caution must be used in interpreting these data because affinity of Fcγ receptors is totally different between humans and mice.
In an earlier study, Garçon et al. for the first time reported high levels of IgG1 antibodies with lower amounts of IgG2 and IgG3 detected after immunization with different vaccine formulations, including cold-adapted attenuated live influenza vaccine intranasal, trivalent inactivated, and purified HA conjugated to diphtheria toxoids (8). In natural infection with H3N2, IgG1 levels increased 18-fold after infection, and the other IgG subclasses increased five- to eightfold. The levels of IgG1 and IgG3 increased after immunization with live cold-adapted vaccines intranasally, and inactivated vaccine induced IgG1, IgG2, and IgG3 subclasses (13). IgG subclass responses were different between the vaccine formulations, and also the increased levels of IgG1 differed by the serological status before vaccination. Stepanova et al. observed the different responses according to the vaccine formulations and age (23). IgG1 and IgG4 responses were detected only in young adults immunized with live influenza vaccine and, contrarily, inactivated vaccine induced IgG1 and IgG3 in young adults and IgG1 alone in elderly. Recently, similar results were obtained by Frasca et al. in a study conducted in 2011–2012, after two seasons had elapsed following the 2009 pandemic, showing IgG subclass antibody responses to H1N1pdm2009 vaccination characterized by robust IgG1 and IgG3 elevation in young adults with lesser IgG3 response in older people basically in the manner of a secondary immune response (7). IgG2 and IgG4 antibody levels were indiscernible in their cases. Interestingly, surprisingly broad IgG subclass antibody responses including igG2 were reported by Yam et al., even in naive infants and young children. However, they carried out their study in the 2009–2010 season with AS03-adjuvanted H1N1pdm2009 vaccine, making direct comparison with our data difficult, although the finding of early development of IgG3 was interesting (25).
Although IgG2 was the second most abundant next to IgG1 in general, an extremely poor or no IgG2 response was also observed in both natural infection and vaccination in the present study. There have been several reports on prominent IgG2 responses after immunization with Streptococcus pneumoniae and Haemophilus influenzae, and these bacterial polysaccharides were potent stimulators of IgG2 response with different responses by age, regulated by Th1 cytokines (20). Divergent IgG2 responses were observed between several reports, with repeated infection or immunological maturation presumably influencing the response (6).
A potential weakness of the present study is its small sample size, which particularly can lead to statistical errors (i.e., not finding a difference that actually exists in the population). However, a high level of statistical significance for the time kinetics of antibody development was observed, even with this small cohort, suggesting that another statistical error (i.e., finding differences where nonexist) is unlikely and that the results are robust enough to demonstrate that influenza A(H1N1)pdm 2009 virus infection induced a unique pattern of antibody development in the human population and also to characterize the configuration of IgG subclass antibody generation.
In conclusion, an immunologically naive population contracted influenza with apparent clinical symptoms with a primary immune response in humoral immunity. However, some poorly understood immune mechanisms existed and worked to modify host experience to a subclinical infection during the pandemic period. Difference observed in the time kinetic patterns of IgG subclass antibody responses might reflect a hitherto unrecognized component of influenza immunity, which poses an attractive research question relevant to the development of the next generation influenza vaccine.
Footnotes
Acknowledgments
We are indebted to Professor Peter M. Olley (Professor Emeritus of Pediatrics, University of Alberta at Edmonton) for his invaluable help in revising the manuscript.
Author Disclosure Statement
No competing financial interests exist.