
Abstract
The major shortcoming of nucleos(t)ide analogue therapy for chronic hepatitis B (CHB) is the frequent requirement for indefinite therapy. Withdrawal of treatment can result in viral rebound with an alanine aminotransferase (ALT) flare leading to hepatic decompensation, while in others this can lead to hepatitis B e antigen (HBeAg) seroconversion. The aim of the study was to identify host immune profiles associated with these different outcomes.
Eighteen HBeAg(+) patients, enrolled on a phase III trial with the nucleoside analogue adefovir dipivoxil, were followed for up to 128 weeks. For the first 48 weeks, all patients received continuous therapy. Subsequently, patients experienced cycles of treatment interruptions due to random drug/placebo misallocations. Host immune profiles were characterized by measuring a panel of serum cytokines before, during, and after each cycle of treatment withdrawal. Virus-specific T-cell responses were also determined at these time points in a subset of patients to elucidate the mechanisms utilized to control hepatitis B virus (HBV) replication post-treatment.
Significantly, elevated levels of IFN-γ, IP-10, and IL-2 on-treatment were associated with HBeAg seroconversion after treatment withdrawal. In these patients, treatment interruption induced further increases in serum IFN-γ levels and marked increases in virus-specific T-cells producing IFN-γ, but minimal alterations in viremia and ALT. In HBeAg(+) patients with low on-treatment levels of serum IFN-γ, the interruption of therapy induced significant elevations in HBV-DNA, ALT, IP-10, and increases in virus-specific T-cells producing IL-10.
Evaluating on-treatment serum cytokines in concert with virologic and clinical parameters may help to identify CHB patients who can successfully discontinue nucleos(t)ide analogue therapy.
Introduction
A
In this pilot study, we longitudinally investigated the relationship between treatment withdrawal, host immune profiles, and clinical outcome in a cohort of HBeAg(+) CHB patients who underwent cycles of treatment interruption as a consequence of random and intermittent drug/placebo misallocations during a clinical trial protocol.
Materials and Methods
Patients/study design
This is an extension of our previous work where we reported that HBeAg loss during 48 weeks of adefovir dipivoxil (ADV) treatment is associated with enhanced virus-specific CD4+ T-cell reactivity in a proportion of patients (3). We obtained longitudinal samples from a cohort of these patients for a further year. During this second year, the active drug (ADV) and placebo were randomly misallocated by the Interactive Voice Response System. This led to patients experiencing multiple cycles of treatment withdrawal with alternating periods on ADV (on-treatment) or placebo (off-treatment) randomly. The duration of each treatment interruption and the total number of interruptions experienced differed in each patient (Table 1). Samples were selected for analysis from each patient and included: (a) start of study (baseline); (b) time points during 48 weeks of continuous ADV therapy; (c) the end of 48 weeks of continuous ADV therapy; (d) before each treatment interruption (on ADV); (e) time points during each treatment interruption (on placebo); and (f ) after each interruption (on ADV).
Group 2 patients had significantly shorter treatment withdrawals than Group 1 (p=0.004) and Group 3 (p=0.041) patients.
TINT, treatment interruption—duration of interruption in weeks.
This study was conducted in accordance with the ethical principles originating in the Declaration of Helsinki and with all International Conference on Harmonization (ICH) Good Clinical Practices (GCP) Guidelines (ICH E6).
Hepatitis serology
Serum HBV markers (HBsAg, anti-HBs, HBeAg, and anti-HBe), anti-HCV, anti-HDV, and antibody to human immunodeficiency virus were tested by commercially available enzyme immunoassays (Abbott Laboratories). Serum HBV DNA levels were quantitated by Amplicor Monitor PCR assay (Roche Molecular systems) with a lower limit of detection (LLD) of 400 copies/mL.
Determination of host immune profiles
We tested a panel of serum cytokines/chemokines including IFN-γ, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12p70, IP-10, FAS-Ligand, and TNF-α. The serum level of these analytes was assessed for each patient at the specified time points. The cytokine levels were quantified by the Cytokine-Bead-Array® (BD Biosciences) according to the manufacturer's guidelines.
In a proportion of patients, the enumeration of HBV-specific, IFN-γ, and IL-10 producing CD4+ and CD8+ T-cells was assessed by ELISpot assays. Peripheral blood mononuclear cells (PBMCs) were isolated from heparinized blood by standard gradient centrifugation, and cryopreserved as previously described (3). The PBMC viability was found to be >95%, as assessed by a Trypan Blue exclusion test, and cell recovery was >90% in all samples. The ELISpot assays were performed as previously described (3). We evaluated the presence of the HLA-A2 haplotype using flow cytometry by staining PBMCs with a mouse anti-human HLA-A2 (OneLambda, Inc.), followed by a fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse IgG secondary antibody (Sigma). In those patients found to be HLA-A2 positive, we assessed the frequency of virus-specific CD8+ T-cells producing IFN-γ using synthetic peptides corresponding to two HLA-A2 restricted CTL epitopes: HBV core FLPSDFFPSVL (aa 18–27) and HBV surface WLSLLVPFV (aa 335–343).
Statistical analysis
Categorical variables were analyzed using the chi-square test. Continuous variables were compared amongst groups using the one-way analysis of variance (ANOVA) test. The comparison between “on” and “off” treatment data within each group was analyzed by independent samples t-tests adjusted for variance inequality or Mann–Whitney U-test whenever appropriate. Correlations between HBV-DNA or ALT and cytokine data were estimated using the Pearson's correlation coefficient. The level of statistical significance was set at p=0.05.
Results
Clinical outcomes
The baseline characteristics of the 18 patients recruited to this study are shown in Table 2. To allow analysis of the relationship between host immune profiles, treatment interruption, and clinical outcome, we stratified the patients into three groups based upon whether patients achieved or did not achieve virologic control before/after treatment interruption (Table 3).
Group 1 patients seroconverted to anti-HBe during the first 48 weeks of continuous adefovir dipivoxil treatment. Group 2 patients lost HBV-DNA and seroconverted to anti-HBe during the second year after experiencing treatment interruption. Group 3 patients did not lose HBV-DNA and did not seroconvert during the 2 years of the study.
TW, treatment week; EOFU, end of follow-up.
Group 1 patients achieved virologic control during the first 48 weeks of continuous ADV treatment, and despite experiencing cycles of treatment interruptions in the second year of the trial protocol, they maintained this control and remained anti-HBe(+) throughout the study.
Group 2 patients remained HBeAg(+) and HBV-DNA(+) during the first 48 weeks of continuous ADV treatment. However, they achieved virologic control after experiencing treatment interruptions in the second year of the study. Interestingly, all these patients seroconverted to anti-HBe during a cycle of treatment interruption.
Group 3 remained HBeAg(+) and HBV-DNA(+) and did not achieve virologic control at any point during the 2 years of this study, neither through continuous ADV therapy nor cycles of interrupted therapy.
None of the 18 patients became anti-HBsAg(+) during the study period, and no patient experienced hepatic decompensation as a consequence of the treatment interruptions. Quantitative HBsAg was not performed due to a lack of sample availability.
Impact of treatment interruption on clinical and viral dynamics
Interruption of therapy had a differential impact on the clinical and viral kinetics within the three groups of patients (Fig. 1). In Group 1 patients, who had achieved viral control during the first 48 weeks of continuous ADV treatment, interruption of therapy induced slightly higher levels of HBV-DNA (off- vs. on-treatment: 3.11±0.16 vs. 2.63±0.02 Log10 copies/mL, mean±SE; p=0.013) but no change in serum ALT levels.
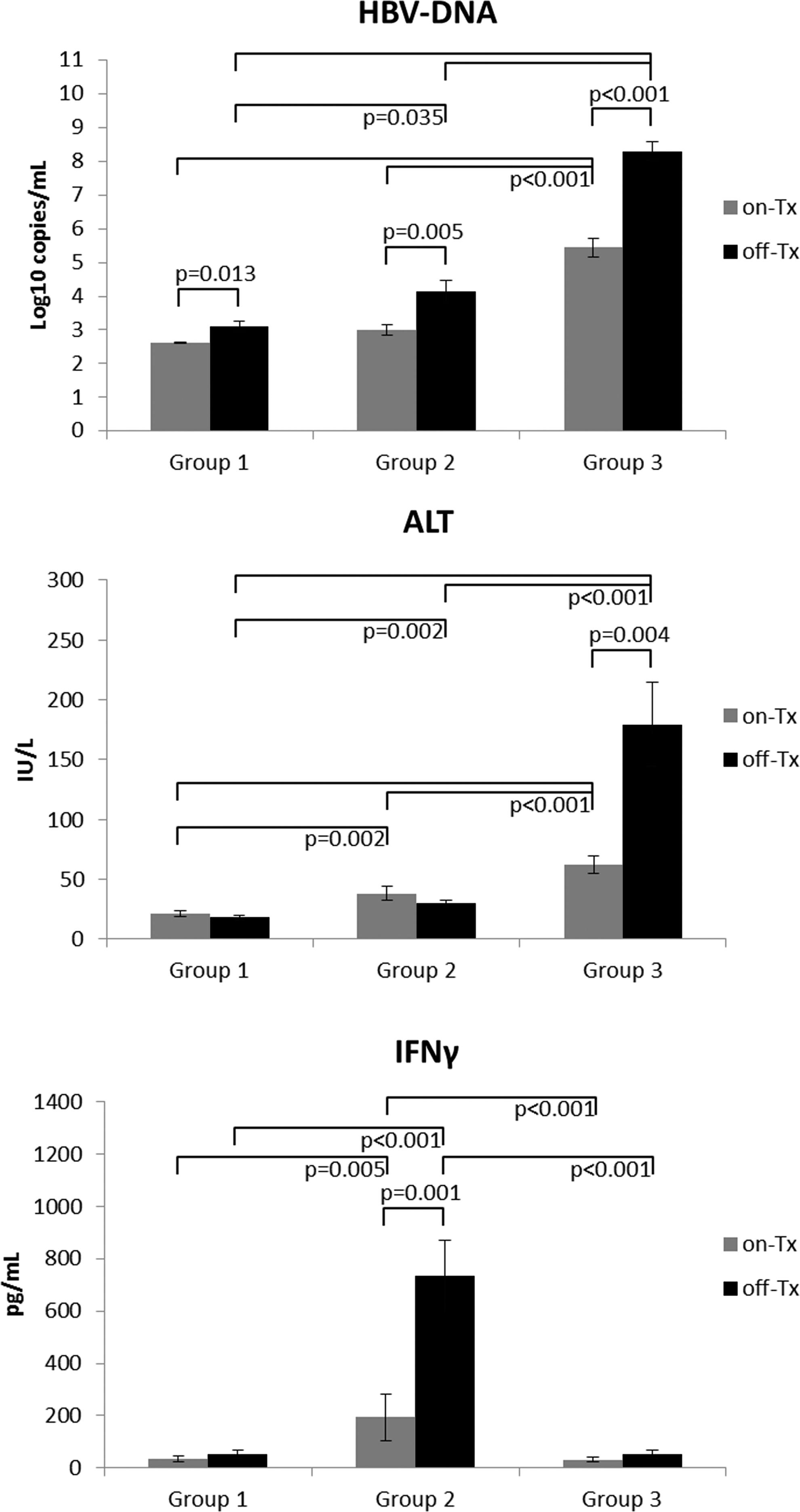
Comparison of clinical and immunologic parameters between patients in Groups 1, 2, and 3. Comparison of HBV-DNA, alanine aminotransferase (ALT), and serum IFN-γ levels between Groups 1, 2, and 3 patients while on and off treatment.
Treatment interruption in Group 2 patients resulted in significant elevations in serum HBV-DNA levels during the off-treatment cycles (off- vs. on-treatment: 4.13±0.33 vs. 3.01±0.16 Log10 copies/mL; p=0.005), and no changes were observed in serum ALT levels. Interestingly, all patients in this group experienced anti-HBe seroconversion during treatment interruption.
Stopping treatment in Group 3 patients, who remained HBeAg(+) throughout the study, induced a marked rebound HBV-DNA (off- vs. on-treatment: 8.30±0.27 vs. 5.44±0.28 Log10 copies/mL; p<0.001), which was significantly different from Group 1 (p=0.001) and Group 2 (p=0.029) patients. Group 3 patients also experienced ALT flares during the off-treatment periods (off- vs. on-treatment: 179.24±35.12 vs. 62.26±7.45 IU/L; p=0.004), which were significantly higher than observed during the off-treatment periods for patients in Group 1 (p=0.001) or Group 2 (p=0.001).
Serum cytokine profiles during treatment interruption
The pattern and evolution of serum cytokine profiles did differ amongst the three groups during the study. Patients in Group 1 who had achieved viral control prior to initiation of treatment interruption experienced very little change in the profile of serum cytokines when treatment was removed (Figs. 1 and 2), reflecting minimal changes observed in HBV-DNA and ALT.
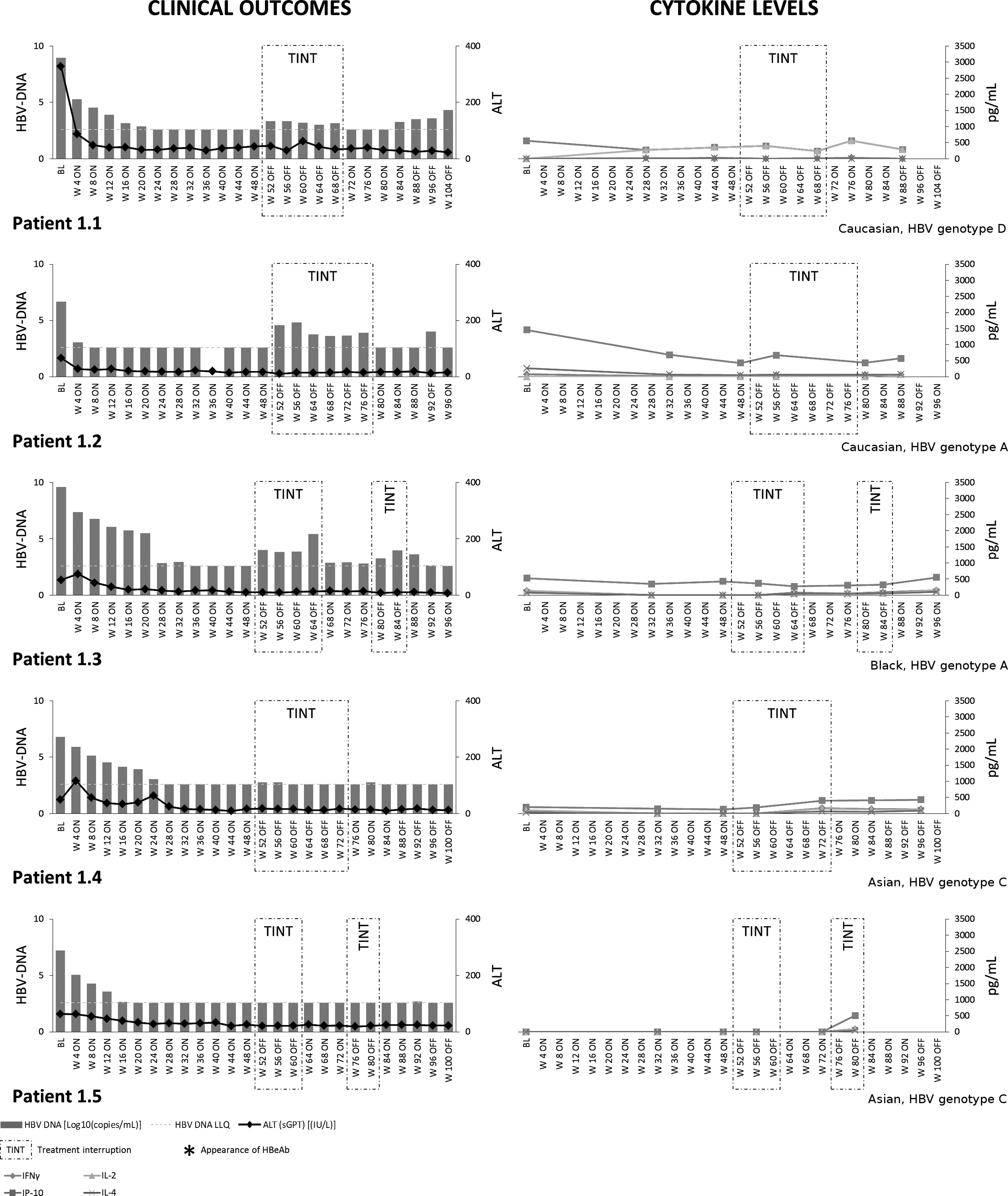
Serum cytokines and clinical outcomes in Group 1 patients. Serum cytokine profiling and clinical outcomes in Group 1 patients. Due to random drug/placebo misallocations, each patient is represented individually.
Patients in Group 2, who became anti-HBe(+) during treatment interruptions, always had on-treatment serum levels of antiviral IFN-γ significantly higher than patients from Group 3 or Group 1 (p<0.005; Figs. 1 and 3). This was also true for serum IP-10 (p<0.032) and IL-2 (p<0.015). When off-treatment, Group 2 patients exhibited further increases in serum IFN-γ levels (on- vs. off-treatment: 193.08±89.11 vs. 733.81±135.64 pg/mL (mean±SE); p=0.001), which were correlated with HBV-DNA (r=0.332; p=0.024).
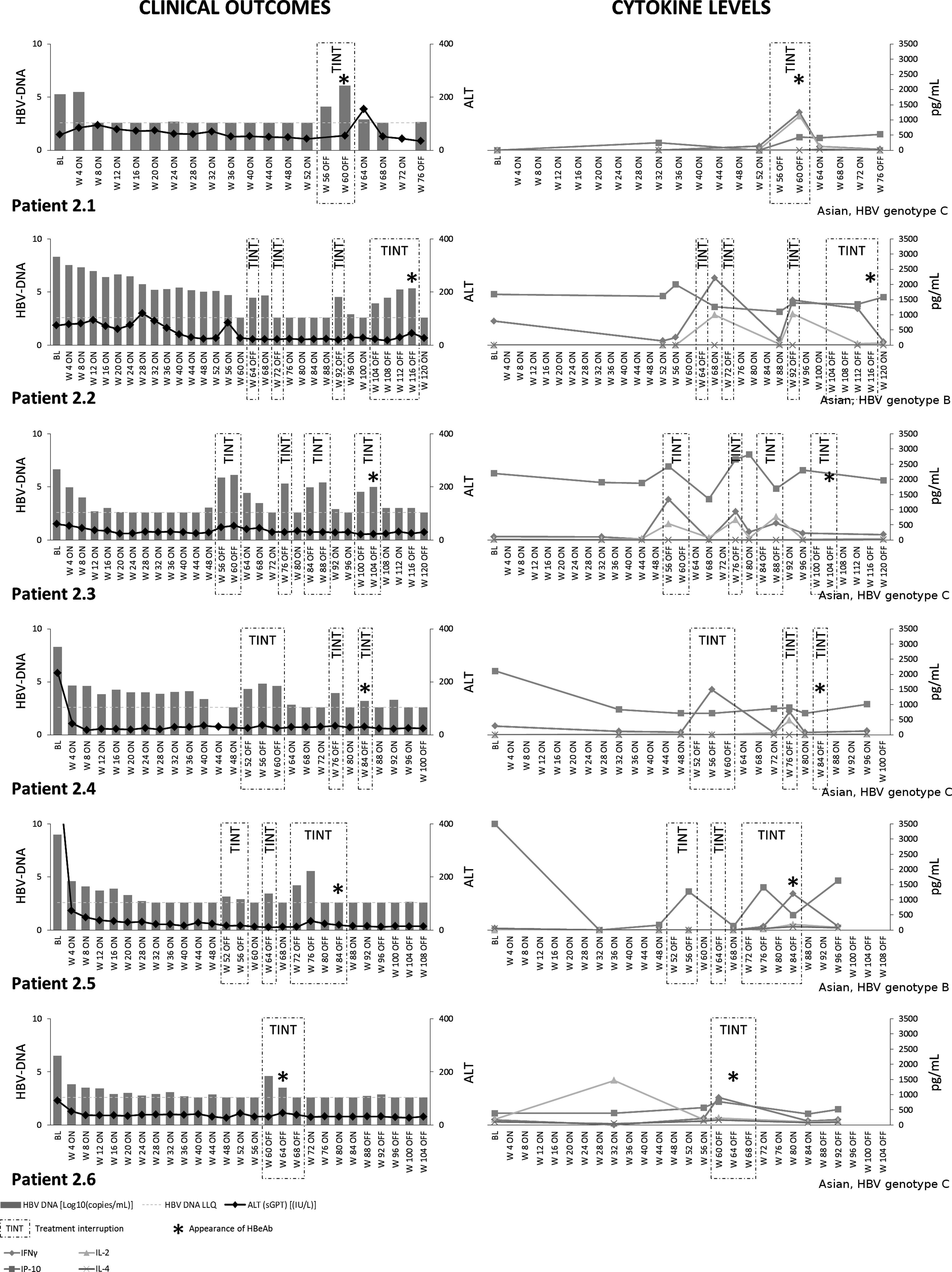
Serum cytokines and clinical outcomes in Group 2 patients. Serum cytokine profiling and clinical outcomes in Group 2 patients. Due to random drug/placebo misallocations, each patient is represented individually.
Group 3 patients exhibited the highest induction of serum HBV-DNA levels and ALT fluctuations as a consequence of the treatment interruption (Figs. 1 and 4). These patients always had lower serum levels of antiviral IFN-γ (p<0.001), IP-10 (p=0.032), and IL-2 (p<0.001) prior to treatment interruption in comparison to Group 2. The off-treatment cycles in Group 3 patients induced marked increases in the serum levels of chemokine IP-10 (off- vs. on-treatment: 936.71±117.76 vs. 533.62±102.84 pg/mL; p=0.013) but not IFN-γ. Detectable levels of IL-4 were also further induced by treatment interruption in these patients (off- vs. on-treatment: 72.72±8.28 vs. 30.06±2.96 pg/mL; p=0.025).
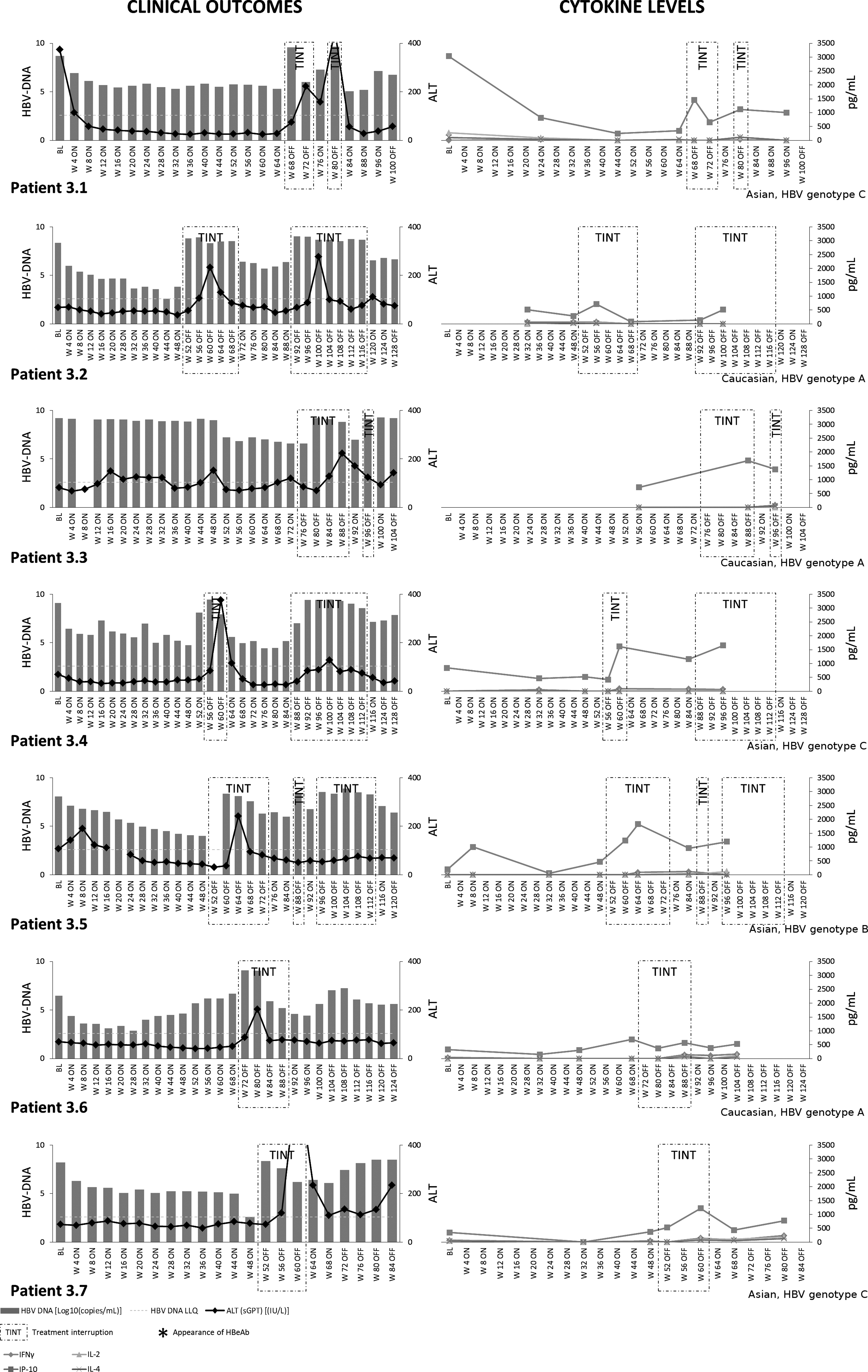
Serum cytokines and clinical outcomes in Group 3 patients. Serum cytokine profiling and clinical outcomes in Group 3 patients. Due to random drug/placebo misallocations, each patient is represented individually.
We observed no significant changes in serum levels of IL-1β, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12p70, FAS-Ligand, and TNF-α between the groups.
Virus-specific T-cell responses
HBV-specific CD4+ and CD8+ T-cell responses were assessed in order to understand whether the differences observed in the serum cytokine profiles between the three groups were associated with changes in the strength and modality of the host adaptive immune response.
Patients in Group 1 showed no changes in the virus-specific T-cell activity as a consequence of interrupted therapy (Fig. 5). In Group 2 patients, off-treatment periods were associated with a pronounced induction of HBV-specific Th1-type T-cell responses with marked increases in the number of HBc- and HBs-specific IFN-γ producing CD4+ T-cells. In an HLA-A2+patient (patient 2.3), we also observed an induction of HBc 18–27 specific IFN-γ producing CD8+ T-cells (Figure 5). In contrast, the patients in Group 3 showed an exacerbation of Th2-like responses during treatment interruption, associated with an induction of HBc- and HBs-specific IL-10 producing CD4+ T-cells (Fig. 5).
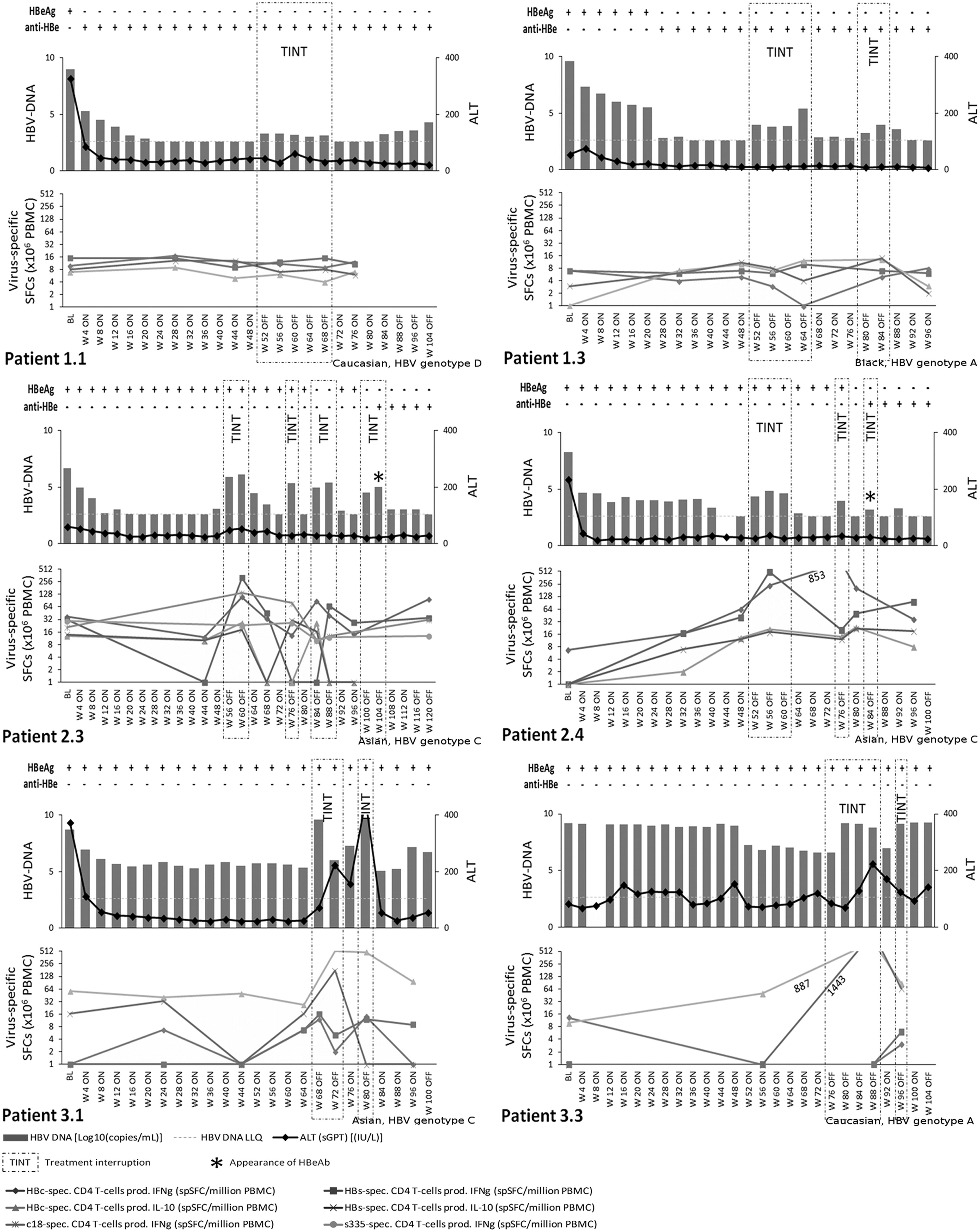
Virus-specific T-cell responses. Longitudinal analysis of HBc- and HBsAg-specific T-cell responses in patients from Groups 1, 2, and 3.
Discussion
In this study, we describe an on-treatment serum cytokine profile that is associated with HBeAg seroconversion following antiviral treatment interruption. The findings, while preliminary, suggest that monitoring serum cytokine levels in CHB patients on treatment with nucleo(s)tide analogues may identify patients who could successfully discontinue therapy. Given the improvement in potency associated with nucleos(t)ide analogue therapy (21), indicators of a favorable outcome after treatment cessation would be useful in the management of HBeAg(+) CHB patients.
We show that elevated serum levels of IP-10, IL-2, and in particular IFN-γ in HBeAg(+) CHB patients on-treatment are associated with a favorable clinical outcome after treatment withdrawal. Furthermore, we show that this treatment interruption can then result in a further induction of serum IFN-γ production and a marked increase in virus-specific T-cells producing IFN-γ. Conversely, low on-treatment serum levels of IFN-γ are associated with exacerbation of disease and uncontrolled viral replication once treatment is ceased, in conjunction with an induction of virus-specific T-cells producing IL-10.
The key role of IFN-γ in mediating the host's control of HBV infection has been described in multiple studies. This potent cytokine has been shown to be associated with resolution of acute hepatitis, whereas weak induction of a Th-1 response results in the progression to HBV persistence (6,24,32). Moreover, we have previously shown that induction of a favorable IFN-γ specific T-cell response in CHB patients on antiviral treatment is associated not only with a beneficial treatment outcome but maybe also be a prerequisite to maintain control of replication (3,5,12,17,26).
However, this is the first report suggesting that the induction of serum IFN-γ levels on-treatment in CHB patients may be an indicator of host immune activation and that treatment interruption in these subjects can lead to virologic control and HBeAg seroconversion without the exacerbation of disease. These findings support our in vitro observations where, in a human cell line model of HBV infection, we found that noncytolytic T-cell immune responses, mediated by IFN-γ, could effectively suppress HBV replication in the absence of cytolytic destruction of infected hepatocytes (22). This suggests that the T-cell effector functions are skewed toward a cytotoxic destruction of infected cells in patients that cannot control viral replication after discontinuation of therapy, whereas virologic control after stopping treatment is associated with a prevailing noncytolytic T-cell effector function, mediated via the production of antiviral cytokines such as IFN-γ. This has important implications for the development of immunotherapeutic approaches for CHB, where the aim of inducing multispecific and broad-ranging anti-HBV immunity should be tailored to enhance predominantly noncytolytic T-cell effector functions to control HBV replication without inducing immunopathology in the host.
The level of the viremia increase observed during the interruption of treatment could be, in part, responsible for induction of favorable T-cell responses and the control of HBV infection. The magnitude of the HBV-DNA increase during the off-treatment periods in patients who control the infection (Group 2) was significantly lower than patients who remain HBeAg(+) (Group 3). However, this may be sufficiently high to serve as an immunological stimulus, exposing viral antigens and inducing an effective immune response, whereas the pronounced and significantly greater increase in viremia observed in Group 3 patients during the interruption of therapy may elicit immune exhaustion or suppression. Indeed, the higher frequency of HBV-specific T-cells producing the immunoregulatory cytokine IL-10 during the off-treatment episodes in Group 3 patients would strengthen this theory, as it has been associated with T-cell exhaustion, viral persistence, and depletion of HBcAg-specific Th1 cells in vivo (1,10,19,23). This observation is supported by previous findings from Webster et al. showing that a HBV-DNA load of <107 copies/mL is the threshold below which circulating multispecific HBV-specific T-cells can be consistently detected (31). Furthermore, our findings are in line with Tan et al., who elegantly showed that lower levels of HBV replication following treatment withdrawal were associated with an increased virus-specific T-cell reactivity but only in patients without hepatic flare (29).
Our results suggest that measuring serum cytokine and chemokine levels to evaluate host immune profiles while patients are on-treatment and during follow-up after stopping treatment may be a complementary tool to clinical and virologic assessments. It would provide criteria to differentiate between patients who may benefit from treatment cessation, resulting in sustained HBeAg seroconversion, from those where stopping treatment would trigger exacerbations of ALT levels and disease.
While the data from this pilot study are compelling, larger prospective studies are required to address some of the limitations of this study. These include the small cohort size, the unstructured nature of the treatment withdrawals, and the lack of sample availability. It would also be important for future studies to investigate changes in the levels of immunotolerant viral antigens, such as HBsAg (25,28) and HBeAg (2,18,20). Further, an in-depth analysis of immunosuppressive mechanisms associated with lack of virologic control in patients with CHB infection would be required. These may include the programmed death-1 pathway (5), T-regulatory cells (9), and their role in determining clinical outcome after nucleo(s)tide analogue treatment withdrawal.
Despite these limitations and the paucity of understanding regarding the use of host immune profiles to aid in the discontinuation of antiviral therapy, our approach is supported by mathematical modeling (11). These models suggest that the optimal timing of when therapy should be stopped during an immunosuppressive infection can be determined by monitoring immune responses in concert with clinical and virologic parameters. Therefore, evaluating on-treatment serum cytokines levels as a measure of immune function in addition to clinical parameters may help to identify CHB patients who can successfully discontinue nucleos(t)ide analogue therapy.
Footnotes
Acknowledgments
This work was supported in part by Gilead Sciences, Foster City, California, and The Foundation for Liver Research, London, United Kingdom.
Author Disclosure Statement
Nikolai Naoumov is currently an employee of Novartis Pharma AG, Basel, Switzerland. Anuj Gaggar is currently an employee of Gilead Sciences, Foster City, California.