
Abstract
Whole inactivated virus (WIV) vaccines for influenza A virus (IAV) provide limited cross-protection to diverse antigenic strains that are circulating or may emerge in a population. Maternal vaccination is used to protect neonatal animals from disease through passive transfer of immunity. It is desirable to vaccinate at a young age to induce active immunity that provides protection against infection before maternal immunity wanes. However, maternal-derived immunity (MDI; antibody or cells) can interfere with vaccine priming. Previous work indicates that vaccine-associated enhanced respiratory disease (VAERD) occurs in pigs following heterologous IAV challenge if pigs were previously vaccinated with WIV vaccine in the presence of matched MDI. However, the component of MDI (antibody or cells) that is required for the mispriming of piglet immunity has not been determined. While antibody from colostrum is absorbed into piglet circulation regardless of the sow from which it receives colostrum, transfer of maternal cells requires colostrum from the biological dam. We used cross-fostering (CF) as a tool to determine if maternal cells are required for the mispriming of piglet immunity upon WIV vaccination in the presence of MDI. Piglets vaccinated in the presence of MDI, regardless of CF, displayed characteristics of VAERD following heterologous challenge. MDI alone (no piglet vaccination) did not provide cross-protection against the antigenic variant. However, it did not induce VAERD. WIV vaccination provided complete protection against homologous challenge when delivered to piglets without MDI. Vaccination in the presence of MDI inhibited an increase in hemagglutination inhibiting (HI) antibody titers to vaccine antigen, but did not alter development of total immunoglobulin levels to vaccine virus. Taken together, the cellular component of MDI did not contribute to the mispriming of piglet immunity to WIV vaccine, but maternal-derived antibody (MDA) alone was sufficient. Future work is aimed at understanding how MDA alters WIV vaccine immunogenicity.
Introduction
M
Influenza A virus (IAV) is endemic in the North American swine population, and strains of the H1N1, H1N2, and H3N2 subtypes co-circulate. While multivalent vaccine formulations are commercially available, their efficacy is limited because of the numerous H1 and H3 subtype antigenic variants circulating in swine. Nearly all licensed vaccines for swine IAV are whole inactivated virus (WIV) preparations (21), yet this vaccine platform induces poor cross-reactive peripheral immunity and no mucosal immunity (13). It is a common management practice in U.S. swine production systems to administer multiple doses of IAV vaccines to sows using WIV vaccine preparations. While this may increase passive immunity in the piglets, MDI can also interfere with vaccine immunogenicity and efficacy in piglets vaccinated within the first months of life (23,24). Unless MDI is matched to the strains to which the piglets are exposed, MDI does not prevent infection or transmission of IAV (1). In addition, vaccine-associated enhanced respiratory disease (VAERD) can occur in pigs that are vaccinated in the presence of MDI and subsequently infected with a heterologous strain of IAV (24). VAERD is a phenomenon that occurs when pigs are given an adjuvanted WIV and are subsequently exposed to a virus of the same subtype that is antigenically distinct from the vaccine virus (7,8,22). In certain instances, MDI presence during piglet vaccination has been shown to lead to VAERD (24). VAERD is characterized by severe pneumonia and exacerbated lung lesions, including bronchointerstitial pneumonia, interlobular and alveolar edema, and hemorrhage (7,8). The component and mechanism by which pre-existing passively acquired MDI alters the subsequent response to vaccine is currently unknown and was a focus of the current study. We hypothesized that IAV-specific memory lymphocytes from the sow, passively transferred to the piglet through colostrum and re-exposed to WIV antigen in the piglet, would contribute to mispriming of piglet immunity that results in VAERD upon heterologous challenge.
Pigs are a natural host of IAV and serve as a relevant model to understand IAV-associated respiratory disease phenomena. Conventional pigs are an outbred population; thus, it is not possible to perform MHC-matched adoptive transfer studies in which lymphocytes from one pig are put into another pig in order to understand the cells contribution to protection or pathology. However, creative methods to circumvent the challenges of using swine to study immune mechanisms are available. In the reported study, we used CF as an experimental approach to determine if MDC played a significant role in vaccine mispriming that resulted in IAV-related VAERD, or if the presence of only MDA alone was sufficient to drive the mispriming response. Synchronized breeding and induced farrowings with piglets born within a short time frame allowed for half of each litter to be CF to a surrogate sow. Thus, piglets suckled colostrum from their respective dam or were CF and received either MDA and MDC (MDI), or MDA only (CF-MDI), respectively. Piglets were then vaccinated with the same WIV as the sows and subsequently challenged with homologous or heterologous IAV to evaluate protection versus development of VAERD. Our results indicate that it was unlikely that MDC played a role in mispriming, as piglets vaccinated in the presence of MDI displayed VAERD when challenged with heterologous virus regardless of CF status. Thus, MDA alone interfered and altered vaccine immunogenicity such that efficacy against homologous IAV was diminished and disease was exacerbated upon exposure to heterologous IAV.
Materials and Methods
Viruses and vaccines
The H3N2 virus used for antigen in the vaccine (A/sw/Texas/4199-2/1998 [TX98]) was propagated in Madin-Darby canine kidney (MDCK) cells. Virus from infected cell culture was clarified, hemagglutinin units (HAU) determined with 0.5% turkey red blood cells, and UV-irradiated as previously described (Vincent and others 2012). The WIV was formulated with TX98 at 64 HAU with commercial adjuvant (Emulsigen-D; MVP Laboratories) added at a 1:5 ratio immediately prior to administration. The WIV-TX98 vaccine was delivered by the intramuscular route in a final volume of 2.5 mL. The challenge viruses—TX98 and heterologous H3N2 A/sw/Colorado/23619/99 (CO99)—were also propagated and titered in MDCK cells. Each pig was challenged with 2 mL of virus at 106 TCID50/mL by the intranasal route. Nonchallenge controls received phosphate-buffered saline (PBS) by the same route.
Experimental outline
Nine sows were procured from a high-health status herd free of IAV and porcine reproductive and respiratory syndrome virus and housed at the National Animal Disease Center. Five sows were vaccinated with WIV-TX98 by the intramuscular route prior to heat synchronization and breeding by artificial insemination. Four nonvaccinated sows were synchronized and bred on the same schedule to produce nonimmune piglet controls. Sows assigned to the vaccinated group received additional doses of WIV-TX98 at 6 and 3 weeks before anticipated farrowing date (three doses in total). Sows were induced to farrow with an intramuscular injection of dinoprost tromethamine (Lutalyse®; Pfizer) the day prior to anticipated farrowing to facilitate synchronized parturition for snatch-farrowing and CF before suckling from their respective dam. As piglets were born, they were kept from suckling, dried, and placed in a tub under a heat lamp. After farrowing was complete, piglets from WIV-TX98 sows were ear-tagged and either CF to another WIV-TX98 vaccinated dam or returned to their respective dam for suckling. The same protocol was used for piglets from naïve sows. No piglet went more than 6 hours before being returned to a sow. Of the five vaccinated sows, only four of the litters were CF, and it was done between pairs of sows. Half of each litter was placed with the other sow. However, CF piglets from each sow were included in each treatment group, and this was also done for the non-CF pigs. This assignment was random and not based on any immune measure (e.g., HI titers). There were seven or eight pigs in each MDI group and five pigs in each naïve group (nonvaccinated sows).
At approximately 2 weeks of age, all piglets were weaned, treated with ceftiofur crystalline-free acid (Excede®; Pfizer), and separated into isolation rooms such that each room had pigs from each treatment group (MDI, CF-MDI, naïve). At 3 weeks of age, pigs were vaccinated with WIV-TX98 and boosted 2 weeks later or remained nonvaccinated (NV). Two weeks following the booster, pigs were challenged by the intranasal route with homologous TX98 or heterologous CO99 virus. Table 1 describes the groups included in the experiment. Piglets were bled by venipuncture on the day of vaccination (pre-vac) and again the day of challenge (post-vac) to evaluate IAV-specific antibody levels. Five days postinfection (dpi), pigs were necropsied to evaluate lung macroscopic lesions and to collect bronchoalveolar lavage fluid (BALF) and sections of lung for microscopic evaluation. BALF collected at necropsy was used to evaluate lung viral titers and levels of IAV-specific IgG. All animal work was conducted in accordance with the National Animal Disease Center's Institutional Animal Care and Use Committee.
CF, cross-foster; MDI, maternal-derived immunity; MDA, maternal-derived antibody; MDC, maternal-derived cells; NV, nonvaccinated; NC, nonchallenged; TX98 and CO99, virus strain; WIV, whole inactivated virus.
Pathological examination of lungs
At necropsy, lungs were removed and the percentage of lung affected by purple-red consolidation typical of IAV infection was determined. The total percentage for the entire lung was calculated based on the weighted proportion of each lobe relative to the total lung volume (9). Lung sections from the right middle lobe or affected lobe were fixed in 10% buffered formalin and processed by routine histopathologic procedures, and slides stained with hematoxylin and eosin. Microscopic lesions were evaluated by a veterinary pathologist blinded to the treatment groups using a scoring system as previously described (7,24). A composite score was computed for each animal and used for statistical analysis.
Virus isolation and titration
At necropsy, BALF was collected by lavaging the lungs with 50 mL of minimal essential media. All samples were stored at −80°C until viral titers and IAV-specific antibody levels were determined after the completion of the study. For virus isolation, 0.2 mL of BALF sample was inoculated onto MDCK-monolayers in a 24-well plate. For virus titration, 0.1 mL of 10-fold serial dilutions of sample were inoculated onto MDCK monolayers in 96-well flat-bottom plates in triplicate. Samples were incubated at 37°C at 5% CO2 for 48 h. For both virus isolation and titration, cells were fixed with 4% phosphate-buffered formalin and stained using immunocytochemistry as previously described (11). The monoclonal antibody (Hb65) specific to IAV nucleoprotein was used for staining. The method of Reed and Muench was used to calculate a TCID50/mL titer for each sample (17). All samples were first evaluated for virus isolation and positive samples were then titered.
Antibody evaluation
For hemagglutination inhibition (HI) assays, serum samples were heat inactivated at 56°C for 30 min, treated with receptor destroying enzyme (RDE), and adsorbed with 0.5% turkey red blood cells to remove nonspecific agglutinins. The HI assay was performed with 0.5% turkey red blood cells using homologous TX98 or heterologous CO99 virus as antigen. Reciprocal HI titers are reported as the reciprocal geometric mean (GM) titer for each group.
To evaluate levels of IgG to homologous TX98 and heterologous CO99 virus, an enzyme-linked immunosorbent assay (ELISA) was performed as previously described with minor modifications (Loving and others 2013). Briefly, ELISA plates were coated with 50 μL of whole virus at 10 μg/mL. Serum was diluted 1:1,000 and BALF diluted 1:4 prior to use in the assay. Horseradish-peroxidase labeled anti-swine IgG (Kirkgaard and Perry) was diluted 1:1,000 and used as detection antibody. Antibody levels are reported as the mean optical density (O.D.) for each treatment group. Serum collected on the day of challenge (post-vac, dpi 0) and BALF collected at necropsy (dpi 5) were used in the assay.
Data analysis
Macroscopic pneumonia scores, microscopic lung scores, log10 transformed BALF virus titers and O.D. for IgG assays were analyzed using analysis of variance (ANOVA) with a p-value of ≤0.05 considered significant and a Tukey post-test performed for pairwise comparisons (GraphPad Prism; GraphPad Software).
Results
MDI, regardless of CF, interfered with induction of antibody responses to intramuscular WIV-TX98 vaccination
Serum was collected from piglets on the first day of vaccination (pre-vac, approximately 3 weeks of age) to evaluate titers of IAV-specific MDA by HI assay (Table 2). HI titers to the IAV virus used to vaccinate dams (TX98) and the antigenic variant (CO99) were evaluated. Groups of piglets that suckled from sows vaccinated with WIV-TX98 (MDI and CF-MDI) had GM HI titers to TX98 of 63 and 102, respectively (Table 2). There was not a significant difference in GM HI titers between CF and non-CF piglets (reciprocal titers to TX98 ranged from 40 to 160). GM HI titers to CO99 ranged from 21 to 24, indicating that WIV-TX98 vaccination induced antibody that minimally cross-reacted against CO99 IAV. Piglets that suckled nonvaccinated sows were seronegative to IAV TX98 and CO99 with HI titers ≤10.
Values reported as reciprocal geometric mean for group.
Prevaccination is day vaccine administered (3 weeks of age); postvaccination is day of challenge (2 weeks post-boost).
Two weeks after administration of the second dose of WIV-TX98 (post-vac) to piglets reared on naïve sows, serum GM HI titer to TX98 was 212 and cross-reactive GM HI titer to CO99 was 31. However, piglets that were vaccinated in presence of MDI had GM HI titer of 27 to TX98, which was not significantly different from CF-MDI vaccinated piglets or nonvaccinated piglets with MDI or CF-MDI (Table 2). Cross-reactive GM HI titer to CO99 in MDI and CF-MDI piglets, regardless of vaccination, were ≤10. Thus, TX98-specific MDA had waned by the day of challenge to below the GM titer considered protective (≥40), while HI titers against CO99 were not detectable.
IgG reactive to whole TX98 and CO99 were detected in post-vac serum as well (Table 3). The amount of TX98-specific IgG in serum was reduced in pigs vaccinated with MDI, regardless of CF status, when compared to naïve WIV-TX98 vaccinated piglets. The average O.D. (±SD) to TX98 in naïve WIV-TX98 vaccinated piglets was 1.15±0.30, but only 0.73±0.30 and 0.77±0.37 in MDI/WIV-TX98 and CF-MDI/WIV-TX98 groups, respectively. However, unlike the HI titers, the IgG O.D. means for the vaccinated piglets with MDI or CF-MDI were higher than piglets that were not vaccinated but suckled WIV-TX98 vaccinated dams, indicating an active response to vaccination. Serum IgG levels cross-reactive to CO99 in naïve WIV-TX98 vaccinated piglets (0.86±0.33) were not significantly different than levels in vaccinated MDI pigs, regardless of CF status (0.76±0.39 and 0.78±0.45, respectively), but they were higher than levels in nonvaccinated piglets with MDI or CF-MDI (0.38±0.16 and 0.41±0.16, respectively). The CO99-specific IgG O.D. means did not correlate with HI titers, as piglets vaccinated with WIV-TX98 in the presence of MDI did not have detectable HI titers to CO99 (Table 2), indicating a production of non-neutralizing antibodies. In addition, piglets without MDI vaccinated with WIV-TX98 had cross-HI GM titers of 31 to CO99, which was higher than MDI-vaccinated groups, but a mean O.D. of 0.86 in the CO99-specific IgG ELISA, similar to the MDI-vaccinated groups. Piglets that were not vaccinated but suckled WIV-TX98 vaccinated dams (MDI/NV and CF-MDI/NV) had measurable serum IgG to both TX98 and CO99 at the time of challenge (Table 3), despite the low or undetectable HI antibody titers (Table 2).
Values with different letters in respective column are statistically different.
Values reported as optical density, mean±standard deviation for indicated group.
Serum was collected on the day of challenge, 2 weeks post-booster vaccination.
CF did not prevent VAERD following heterologous challenge of pigs previously vaccinated in the presence of MDI
Mild lung pathology was observed following challenge of naïve pigs with either TX98 or CO99, with the percent pneumonia averaging less than 5% of the total lung (Fig. 1A and B). Naïve pigs vaccinated with WIV-TX98 and then challenged with homologous TX98 were protected from challenge, as lung lesion scores were significantly reduced when compared to the naïve NV/TX98 group. While there were significant differences in percent pneumonia within the WIV MDI-vaccinated groups following TX98 challenge, percentages were not different compared to NV/TX98 groups. In fact, percent pneumonia group means were numerically higher in the WIV vaccinated pigs with MDI, regardless of CF, compared to NV groups challenged with TX98. In addition, microscopic lesion scores were not significantly different among any of the groups challenged with TX98 (Fig. 1A). These data indicate that WIV-TX98 vaccine, administered to pigs with MDI and regardless of CF, did not elicit a protective immune response. Only naïve pigs given WIV-TX98 were subsequently protected against lung pathology associated with TX98 challenge.
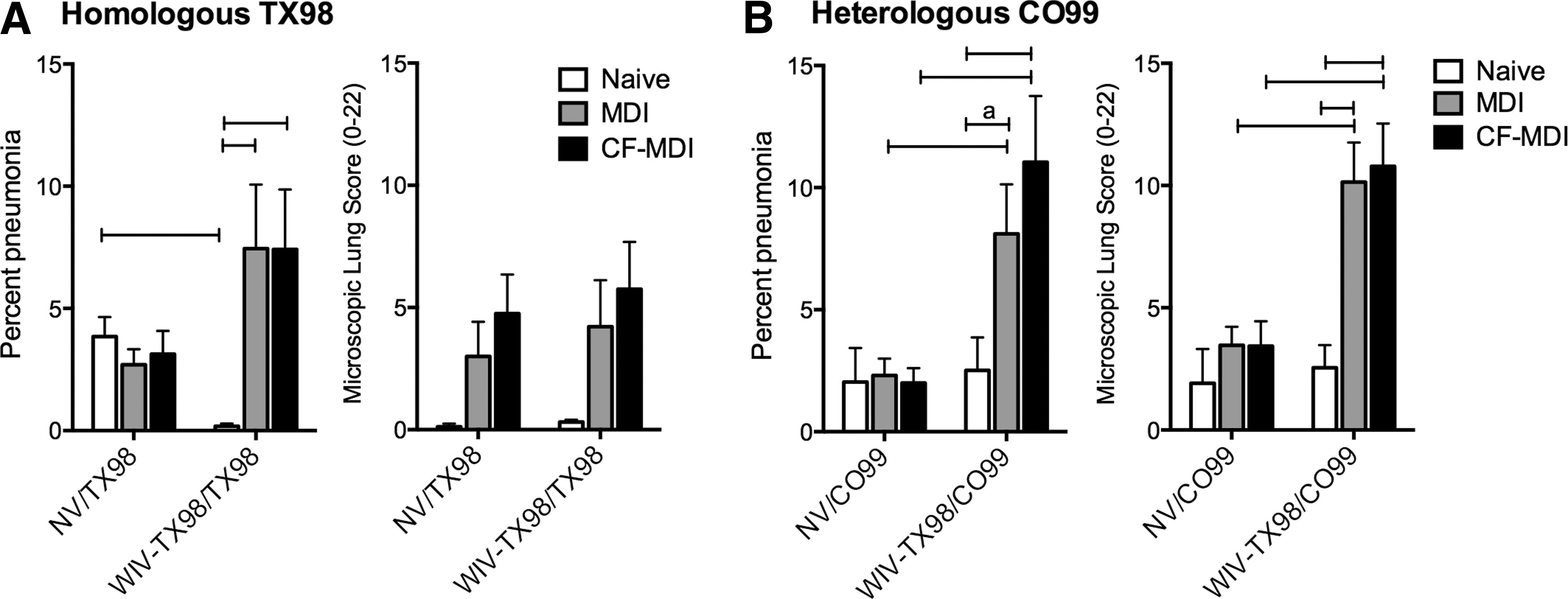
Cross-fostering (CF) does not prevent increased lung pathology following heterologous CO99 challenge of pigs previously vaccinated in presence of maternal-derived immunity (MDI). The percent pneumonia and lung microscopic lung lesions were assessed following
While WIV-TX98 elicited an immune response that provided protection against homologous TX98 challenge in naïve pigs, it did not provide protection against heterologous CO99 challenge, as there was not a significant reduction in lung pathology between naïve WIV-TX98/C099 and naïve NV/C099 groups (Fig. 1B). WIV-TX98 administered to pigs in the MDI and CF-MDI groups resulted in enhanced macroscopic and microscopic lung lesions following heterologous CO99 challenge (Fig. 1B). The microscopic characteristics of the lung lesions in the MDI/WIV-TX98 and CF-MDI/WIV-TX98 groups challenged with CO99 were consistent with previous reports of VAERD in this model (24). This included peribronchiolar lymphocytic cuffing, enhanced neutrophil accumulation in bronchioles and alveoli, and intrapulmonary airways with epithelial necrosis and/or proliferation. While these features may be independently observed in naïve pigs infected with IAV, the magnitude of severity and consistent presence of all features is a hallmark of VAERD (7).
Viral titers in the lungs were not reduced following heterologous CO99 challenge after WIV-TX98 vaccination in the presence of MDI
Residual TX98-specific MDI, present in both MDI/NV and CF-MDI/NV pigs, did not provide protection against TX98 challenge, as viral titers in naïve NV/TX98, NV/MDI, and NV/CF-MDI pigs were not significantly different (Fig. 2A). While pigs in the MDI/WIV-TX98 and CF-MDI/WIV-TX98 groups challenged with TX98 displayed an increase in macroscopic pneumonia (Fig. 1A) over NV groups, titers of TX98 in the BALF were significantly reduced in WIV-TX98 groups to respective NV groups, regardless of MDI and CF-MDI status (Fig. 2A). However, pigs from naïve sows that were given WIV-TX98 had a significant reduction in lung viral titers compared to pigs given WIV-TX98 in the presence of MDI, regardless of CF (Fig. 2A). Thus, WIV-induced immune protection from TX98 replication in the lungs was reduced in pigs vaccinated in the presence of MDI and CF did not significantly impact the outcome.
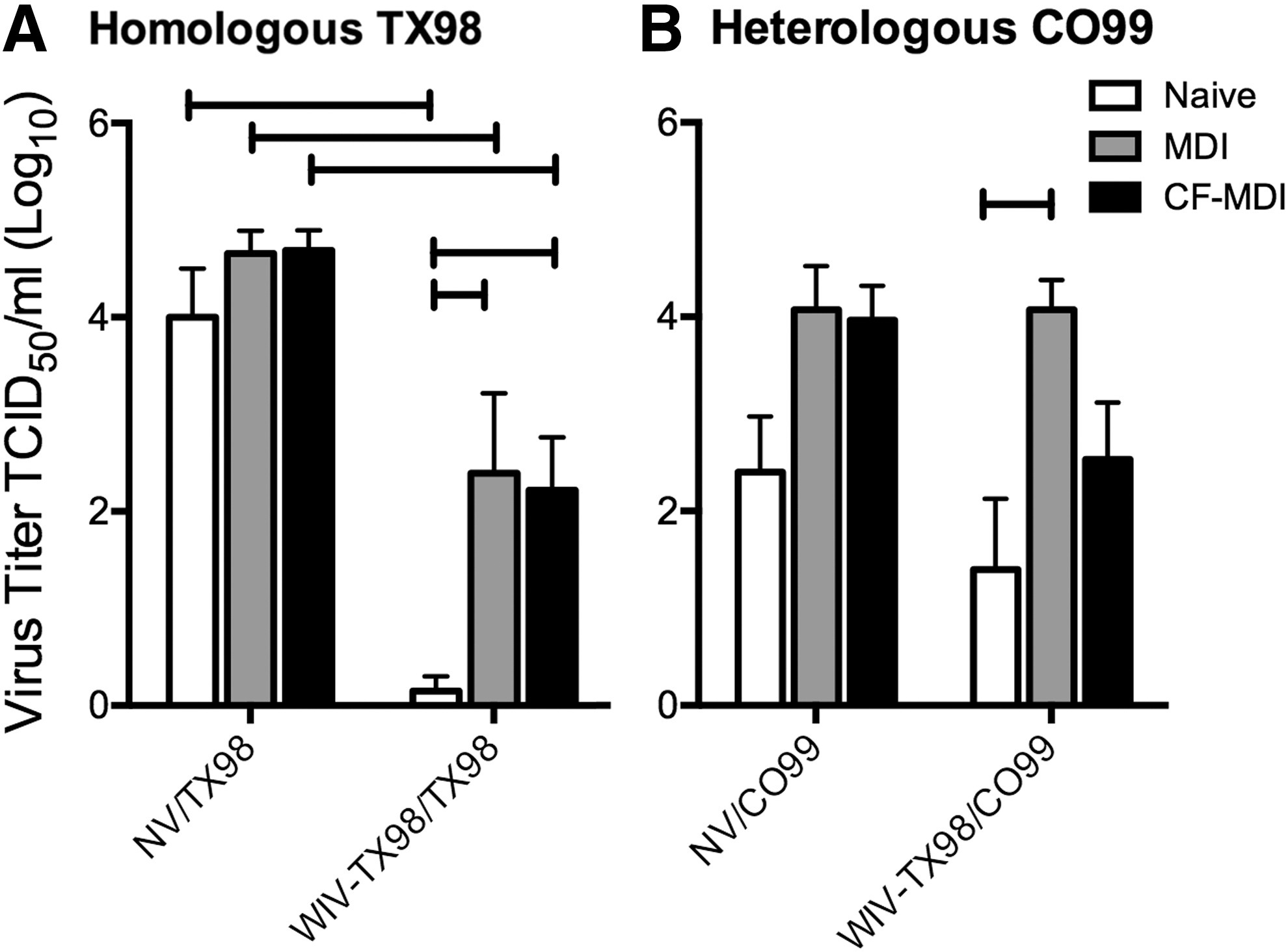
Viral titers in the lungs were not reduced following heterologous CO99 challenge after WIV-TX98 vaccination in the presence of MDI. The presence of MDI, regardless of CF, interfered with WIV-TX98 vaccine efficacy. Viral titers in the bronchoalveolar lavage fluid (BALF) collected at 5 days postinfection were determined following
Residual TX98-specific MDI, present in both MDI and CF-MDI pigs, did not provide protection against heterologous CO99 virus replication in the lungs (Fig. 2B). In addition, immunity elicited following WIV-TX98 vaccination of naïve pigs did not provide protection against viral replication of heterologous CO99 challenge virus, as there was not a significant difference in viral titers between naïve NV/CO99 challenged and WIV-TX98/CO99 challenged groups. There was a trend for increased CO99 virus titers in the BALF of nonvaccinated MDI/CO99 and CF-MDI/CO99 pigs compared to naïve/CO99 pigs (Fig. 2B). However, these differences were not statistically significant (p=0.07). Pigs given WIV-TX98 in the presence of MDI, but not CF-MDI, had increased titers of CO99 virus in the BALF at necropsy when compared to naïve pigs given WIV-TX98. However, none of the WIV-TX98 groups were significantly different from respective NV/CO99 groups (Fig. 2B).
IAV-specific IgG levels in the BALF were not significantly different among pigs vaccinated under differing immune statuses
To determine if antibody in the lungs possibly contributed to differences in disease pathogenesis following challenge, IgG reactive to TX98 and CO99 in BALF was evaluated. Nonvaccinated pigs, regardless of receiving MDI through colostrum, did not have significant levels of TX98- or CO99-specific IgG in the BALF following challenge (Fig. 3). However, all pigs that received WIV-TX98 had IgG reactive to both TX98 and CO99 in the BALF following challenge. The immune status of the piglet at the time of vaccination (naïve, MDI, or CF-MDI) did not impact the amount of IgG reactive to TX98 or CO99 detected in BALF, nor did the challenge virus used (TX98 or CO99) impact the presence or levels of IgG measured (Fig. 3).
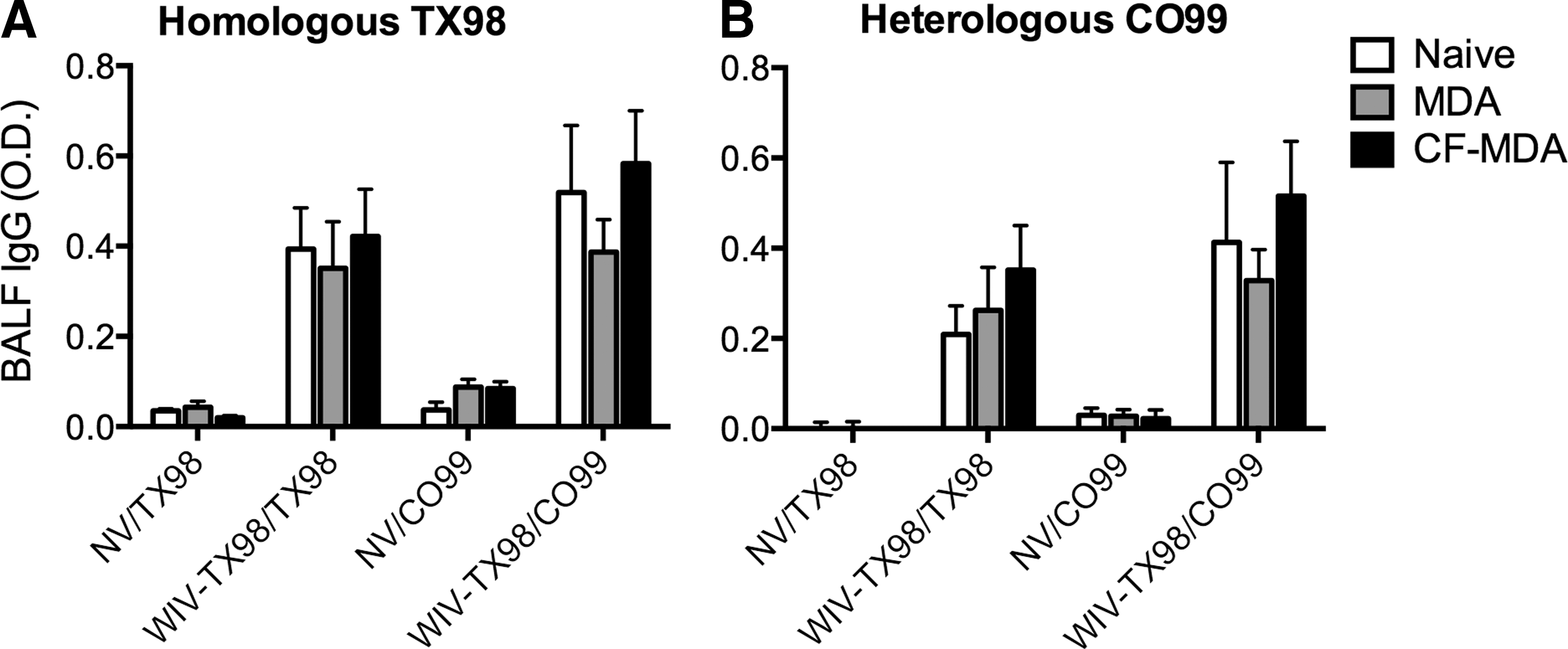
Influenza-specific IgG levels in the bronchoalveolar lavage fluid were not significantly different between pigs vaccinated under differing immune statuses. Levels of IgG in the bronchoalveolar lavage fluid (BALF) collected at 5 days postinfection were measured against
Discussion
The antigenic diversity of IAV strains currently circulating in North American swine is a major problem for pork producers because it makes controlling infection and disease with currently available commercial vaccines ineffective. While vaccinating sows may result in passively acquired MDI in piglets that can protect against matched homologous virus when adequate titers are achieved (1), the IAV strains in the vaccine and subsequent strains to which pigs are exposed can be antigenically distinct. Thus, MDI to IAV in swine production may be of limited benefit (5). The current study confirms previous reports that IAV-specific MDI in piglets interferes with efficacy of WIV vaccines administered to piglets (10,11,24) and even resulted in VAERD upon heterologous infection. Previously, our group described the occurrence of VAERD in pigs that were vaccinated with WIV in the presence of matched MDI and subsequently challenged with an antigenic variant of IAV (24). The component of MDI that altered the immunogenic nature of the WIV vaccine in piglets was the focus of the current study. Our results indicated that MDC were not involved in this process. Instead, MDA was the primary immune factor that interfered with vaccine efficacy.
Maternal immunization, for the passive transfer of protective immunity, is a strategy used to provide protection to neonatal animals, and has been shown to have success (reviewed in Crowe (6)). However, maternal vaccination for subsequent protection of the neonate can also significantly alter neonatal responses to vaccination. The component of maternal immunity (antibody or cells) that negatively impacts the neonate's primary response to vaccination has not been clearly defined. In pigs, antigen-specific MDC can be detected in pigs for at least 4 weeks following birth (14). Early work on IAV in mice includes reports in which IAV-specific lymphocytes significantly contributed to increased pneumonia and mortality following challenge, and this response was related to the priming antigen (live vs. inactivated virus) (4). These data led us to hypothesize that MDC, particularly IAV-specific lymphocytes derived from WIV vaccination in the mother, would impact the development of the neonate immune response to IAV vaccination that contributes to VAERD upon heterologous challenge. Our initial thoughts were that maternal memory B- or memory T-cells would alter the development of an active piglet response, possibly altering the cytokine milieu in which response developed and/or skewing the development of pig immunoglobulin to a specific isotype. However, our data indicate that transfer of IAV-specific antibody (MDA) alone altered vaccine immunogenicity such that lesions were exacerbated following heterologous challenge and only partial protection was provided following homologous challenge. Thus, we did not complete any additional studies on the effect maternal lymphocytes may play on the developing piglet immune response. Instead, how antibody bound to virus alters antigen-specificity of the piglet antibody response will be further explored.
Our data support prior reports that IAV-specific MDI alters primary immune responses in the neonate upon vaccination with WIV (Tables 2 and 3) (10,24). WIV-TX98 administered to piglets with WIV-TX98-specific MDI was partially protective, as pigs in the MDI/WIV-TX98/TX98 and CF-MDI/WIV-TX98/TX98 groups had pneumonia and pathologic changes consistent with NV/TX98 group (Fig. 1) but reduced viral titers in the BALF (Fig. 2). Serum HI titers to homologous challenge virus (TX98) in these groups were below the level considered protective (≤40). However, NV piglets with MDI still present at the time of TX98 challenge also had HI titers of a similar level (≤40), and these pigs were not protected against pathology or virus replication, suggesting a protective mechanism for WIV in MDI piglets other than induction of neutralizing antibodies. Piglets without IAV-specific MDI that were vaccinated with WIV-TX98 had a GM HI titer in the predicted protective range (≥40), and indeed, these pigs were protected against homologous TX98 challenge. These data reiterate the finding that vaccination in the presence of MDI significantly interferes with vaccine immunogenicity and subsequent efficacy. Thus, maternal vaccination as a disease control strategy may provide some protection in the neonate, but it is not advantageous when paired with vaccination of the neonate with the same vaccine when MDI is still present.
The amount of MDA circulating in the piglet at the time of WIV vaccination likely impacted neonatal vaccine efficacy and VAERD in vaccinated/challenged piglets. In our previous report, HI titers to homologous vaccine virus (TX98) in piglets with MDI at the time of vaccination were nearly 2.5-fold higher than in the current study. Subsequently, there were significant differences in the amount of IAV-specific IgG in the BALF between MDA+ and MDA− vaccinated pigs in the prior study (Vincent and others 2012) but not the current study (Table 3 and Fig. 3). Thus, the degree to which MDI altered the development of IAV-specific immunity in the piglet was dependent on the magnitude of IAV-specific antibody present at the time of piglet vaccination. This could be affected by the vaccine regimen in the sow and the amount of colostrum suckled. Regardless, in the current study, the immunogenicity of the WIV-TX98 vaccine was altered by MDI, and VAERD occurred following heterologous challenge of piglets that were vaccinated in the presence of MDI. These data indicate that the amount of MDA in piglet circulation does not have to be high to interfere with subsequent vaccine immunogenicity.
The finding that the virus titers in the BALF of the CF-MDI/WIV-TX98 group were not significantly increased over naive piglets given WIV-TX98 and then challenged with CO99 was unexpected (Fig. 2B). However, the MDI/WIV-TX98/CO99 group had a significant increase in BALF titers when compared to the WIV-TX98/CO99 group. This is consistent with our previous report. However, the virus titers in the BALF of all CO99 challenged pigs were lower than expected, with titers ranging from 101.8 to 104.3 TCID50/mL. In our previous study with the same virus, the average BALF virus titer following CO99 challenge was approximately 106 TCID50/mL. The challenge virus in the previous study was administered by the intratracheal route, and in the current study, intranasal challenge was used, likely explaining the differences in titers between the two studies.
We used ELISA to evaluate IAV-specific IgG in serum collected on the day of challenge. One consistent feature of VAERD appears to be the presence of cross-reactive but non-neutralizing serum antibody to the antigenically mismatched challenge virus (8,24). Given that CF did not prevent VAERD following WIV vaccination in the presence of MDI, it was likely that MDA, but not MDC, played a significant role in altering vaccine immunogenicity. In the current study, WIV-TX98 vaccination in the presence of MDI did not result in measurable serum cross-reactive HI titers to CO99 (Table 2). However, WIV-TX98 vaccination in the presence of MDI, regardless of CF, elicited production of IgG antibodies that were cross-reactive to CO99 virus as measured by a whole-virus ELISA (Table 3). IgG reactive to the CO99 virus was also detected in serum collected from MDI and CF-MDI groups that were not vaccinated. However, levels were increased in MDI/WIV-TX98 and CF-MDI/WIV-TX98 groups following vaccination. These data indicate that WIV-TX98 administration in the presence of MDI significantly increased IgG antibody levels that cross-reacted with CO99 virus, but not to epitopes that increased HI titers. The molecular and cellular mechanism in which MDA alters the presentation of WIV vaccine antigen is currently unknown. It is possible that MDA complexes with vaccine antigen and is then taken up by receptor-mediated phagocytosis, and this possibly alters MHC processing and presentation. B-cell receptor recognition of antigen complexed with antibody may alter FcR expression and subsequent activation, or MDA binding to vaccine antigen prevents B-cell recognition of antigen through epitope masking. This latter mechanism may drive antibody responses to non-neutralizing epitopes. Regardless of the mechanism, antibody against non-neutralizing epitopes develops upon WIV administration in the presence of MDA and is not protective. The intranasal administration of live-attenuated influenza virus (LAIV), even in the presence of matched MDI, is protective against homologous challenge and does not result in VAERD following heterologous challenge (12). LAIV vaccine immune priming is mainly at the respiratory mucosa and thus may not complex with MDA present in circulation. LAIV vaccines replicate primarily in the upper respiratory tract and would not come into contact with IgG from circulation that transudates into the lower respiratory tract but not upper. This may explain why LAIV vaccines do not prime a response resulting in VAERD, even in the presence of MDI. Current work in our laboratory is aimed at understanding how non-neutralizing but IAV-specific antibody complexed with virus alters antigen presentation and subsequent immune responses that contribute to VAERD.
BALF IgG profiles to TX98 virus following challenge did not mirror serum IgG profiles on the day of challenge. For instance, BALF IgG levels to TX98 in naïve pigs that were given WIV-TX98 were similar to IgG levels in MDI/WIV-TX98 and CF-MDI/WIV-TX98 groups (Fig. 3A). However, in the serum, there were significant differences in IgG specific to TX98 between these groups (Table 3). In the serum, residual MDA that reacted to both TX98 and CO99 was detected in NV pigs. Yet, antibody was not detected in the BALF of these pigs following challenge. This may be due to the reduced sensitivity of the assay when using BALF as a sample, as O.D. values in general were reduced in BALF samples. Additional work is required to determine if cross-reactive, non-neutralizing antibody is present in the lungs prior to challenge, and subsequently drives a pathologic response upon heterologous challenge.
Collectively, this work has implications for swine production systems and also for human health. Maternal vaccination for passive protection may be a method for protecting the neonatal animal at a vulnerable age. However, consideration of subsequent vaccination regimen and sequelae if infection with heterologous virus occurs should be considered. In the case of IAV, MDC do not appear to interfere with the immunogenicity of neonatal vaccination as it relates to WIV. However, MDA can significantly alter the neonatal immune response to IAV WIV vaccine and alter immunogenicity such that heterologous infection results in VAERD. Identifying the mechanism by which MDA alters immunity toward non-neutralizing epitopes may lead to strategies to circumvent the problem. Mucosal administration of LAIV vaccines has already been shown to be efficacious, even when delivered in the presence of MDA. However, a LAIV vaccine has yet to be commercially available for swine producers, and the vaccine in humans is not labeled for use in children younger than 18 months. Thus, additional solutions are needed to improve the efficacy of vaccines in neonatal humans and pigs.
Footnotes
Acknowledgments
We would like to thank Gwen Nordholm, Zahra Olson, Kim Driftmier, Michelle Harland, and Deb Adolphson for excellent technical assistance. We would also like to thank Jason Huegel and Brian Pottebaum for assistance with animal work. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendations or endorsement by the U.S. Department of Agriculture (USDA). USDA is an equal opportunity employer.
Author Disclosure Statement
No competing financial interests exist.