
Abstract
Dengue virus (DENV) causes a febrile disease, infecting around 50–100 million people annually. The relationship between DENV proteins and host cellular responses during infection is unclear. This study investigated the interaction of host protein disulfide isomerase (PDI) with DENV proteins and role of lipid rafts in viral immunopathogenesis. Host viral protein interactions were studied by co-immunoprecipitation and co-localization. It was found that PDI interacts with DENV nonstructural protein 1 (NS1) intracellularly as well as on the surface in the lipid raft domain. Disruption of this key interaction between PDI and NS1 could be an important therapeutic strategy to block DENV infection.
Introduction
D
Recently, certain hydrophobic regions of the NS1 dimer have been suggested to interact with the ER membrane (2). The viral replication assembly located along the ER membrane includes the viral nonstructural proteins. A hydrophobic protrusion has been identified on one face of the NS1 dimer, which has been suggested to interact with the viral NS4B protein of the replication complex, thereby being involved in organizing the viral replication complex (2). Moreover, NS1 has been reported to associate with the cellular lipid bilayer and remodels it to its benefit by forming lipoprotein nanoparticles. Eventually, hexameric NS1 proteolipid particles are formed, which are secreted from the infected cells and interact with the complement system (2).
Protein disulfide isomerase (PDI) is a host cellular chaperone with oxidoreductase activity (12,17). It is capable of oxidizing nascent proteins. PDI can identify misfolded proteins in the cell and direct their degradation via the unfolded protein response. PDI found on the platelet surface has been shown to be recognized by anti-DENV NS1 antibodies (5). Moreover, it has also been shown that PDI shares regions of homology with DENV NS1. PDI can cross-react with the anti-DENV NS1 antibodies and, as a consequence, may lead to a reduction in platelet aggregation. DENV entry into endothelial cells has been shown to be inhibited on silencing PDI, indicating that PDI is involved in viral entry into the host cell (27). Studies have also shown that PDI co-localizes with cell surface lipid rafts along with DENV envelope protein, leading to the activation of cell surface integrins (β1 and β3), which has direct implications in aiding the virus to gain entry into the cell (27,34). In addition, PDI also has a role in viral replication within the host cell (6,16).
Very little is known about the relationship between the DENV structural and nonstructural proteins and cellular responses during infection. In a previous study, it was reported that PDI was upregulated during DENV infection (22). The present study is the sequel of the same, to find out the interaction between viral proteins and PDI. The data clearly indicates that host protein PDI interacts with DENV NS1 protein intracellularly, as well as on the surface in the lipid raft domain. Therefore, disruption of this key interaction between PDI and NS1 could be an important therapeutic strategy that could block DENV infection.
Materials and Methods
Cells and virus
Human monocytic cell line THP-1 was obtained from the National Centre for Cell Science (Pune, India) and maintained in RPMI (Sigma) supplemented with 10% fetal bovine serum. Dengue virus type-2 (DENV) New Guinea C strain grown in C6/36 insect cell line with viral titer 106 pfu/mL was obtained from the International Centre for Genetic Engineering and Biotechnology (New Delhi, India).
Immunoblotting
Whole cell extract was prepared from THP-1 cells either mock or DENV infected with or without Bacitracin treatment, as per standard protocol. Lysates containing 40 μg protein were separated on 12% sodium dodecyl sulfate-polyacrylamide gels following the method described by Laemmli et al. (18) and transferred to polyvinylidene difluoride (PVDF) membranes, as described by Towbin et al. (26). The membranes were incubated for 1 h with 3% bovine serum albumin in Tris-buffered saline (TBS; 0.1 M Tris HCl, pH 7.4, 0.9% NaCl) to block nonspecific binding followed by washing with TBST20 (0.1% Tween-20 in TBS) and incubated with primary antibody (Env, prM, C, and NS1 from Abcam; NS4B, NS3, and NS5 from GeneTex; PDI from Thermo Scientific; and β-actin from Sigma). Subsequently, the membranes were washed three times for 10 min with TBST20 and incubated with secondary antibody horseradish peroxidase (HRP)–conjugated IgG (AbD Serotec) against the host primary antibody, respectively. The proteins were detected by chemiluminescence (Sigma).
Co-immunoprecipitation
Briefly, 15 μg of PDI-specific monoclonal antibody (Thermo Scientific) was coupled to 50 μL of AminoLink plus amine-reactive resin (aldehyde-activated beaded agarose) prior to the addition of 400 μg of crude cellular lysate to the resin beads. Co-immunoprecipitation (Co-IP) was carried out using a Pierce Co-IP Kit (Thermo Scientific) in accordance with the manufacturer's protocol. Immunoblotting was done as described above to probe PDI protein (bait) and DENV protein(s) (prey) such as DENV Env, prM, NS1, and NS4B (Abcam) Capsid, NS3, and NS5 (GeneTex).
Immunofluorescence staining
Surface staining for lipid rafts and PDI
THP-1 cells (2×106 cells/mL) were either mock (cell culture medium treated) or DENV infected at MOI-3 for 24 h. For lipid raft staining on the cell surface, harvested cells were first stained for lipid rafts using lipid raft labeling kit (Invitrogen), as per the manufacturer's protocol. This was followed by PDI staining using PDI Antibody (Thermo Scientific) at 1:200 dilution for 1 h, and antimouse IgG-PE secondary antibody (BD Biosciences) at 1:500 dilution for 1 h on ice, and prepared for immunofluorescence imaging using a BX51 Fluorescence Microscope (Olympus).
Surface staining for PDI and NS1
Mock- and DENV-infected cells were harvested and washed once. Cells were stained with PDI as described above. Cells were also incubated at 1:200 dilution with DENV NS1-FITC (Biorbyt) conjugated antibody and prepared for immunofluorescence imaging using a fluorescence microscope.
Intracellular staining for PDI and NS1
For intracellular staining of PDI and NS1 proteins using a fluorescence microscope, harvested cells were permeabilized after fixation (using cytofix buffer for 15 min in the dark followed by washing and permeabilization using cytoperm buffer for 10 min and washing with 1×phosphate-buffered saline at room temperature) and then immunostained with PDI and NS1 as described above. Cells were also nuclear stained using Hoechst (Sigma-Aldrich) at a final concentration of 0.05 μg/mL.
Cell-binding assay
THP-1 cells at a concentration of 2×106 cells/mL were treated with either PDI inhibitor Bacitracin (100 μg/mL) or cholesterol inhibitor MβCD (30 mM), and cultured overnight. Inhibitor treated or untreated cells were infected with DENV at MOI-3 for 80 min on ice for viral adsorption. Cells were washed with cold media to remove unbound virus and fixed using 200 μL cytofix buffer (eBioscience) by incubating the cells for 15 min at room temperature. Immunostaining for DENV Env protein was done using anti DENV ENV antibody raised in mouse (Abcam) at 1:200 dilution, followed by antimouse IgG-FITC secondary antibody (Jackson ImmunoResearch) at 1:200 dilution, for detecting the cell surface bound virus by flow cytometry using BD FACSCalibur using CellQuest Pro software.
Statistical analysis
The results are expressed as mean±standard error of the mean, and all the statistical comparisons were carried out using Student's t-test for repeated measurements wherever applicable. Experiments were repeated at least three times. A significance level of p<0.05 was set.
Results
Identification of PDI-interacting protein(s) by Co-IP
In order to determine which of the DENV protein(s) interact with the host protein PDI in DENV infected cells, Co-IP of mock and DENV-infected cell lysates was performed using specific antibodies against human PDI. Presence of PDI and viral protein(s) in the whole cell lysates (Input) of uninfected and infected cells was confirmed by immunoblotting in order to rule out any false positive signals. The bait–prey complex captured in the Co-IP column was subjected to simultaneous immunoblotting analysis for PDI and one of the seven DENV proteins. This was repeated at least three times with each of the seven DENV proteins (Env, prM, capsid, NS1, NS3, NS4B, and NS5) along with PDI. Immunoprecipitation of PDI mock cell lysate did not show co-precipitation of DENV proteins, whereas, in DENV-infected cell lysate, the viral NS1 protein was found to co-precipitate with PDI, as shown by immunoblot analysis (Fig. 1). However, the other DENV proteins did not show interaction with PDI by Co-IP.
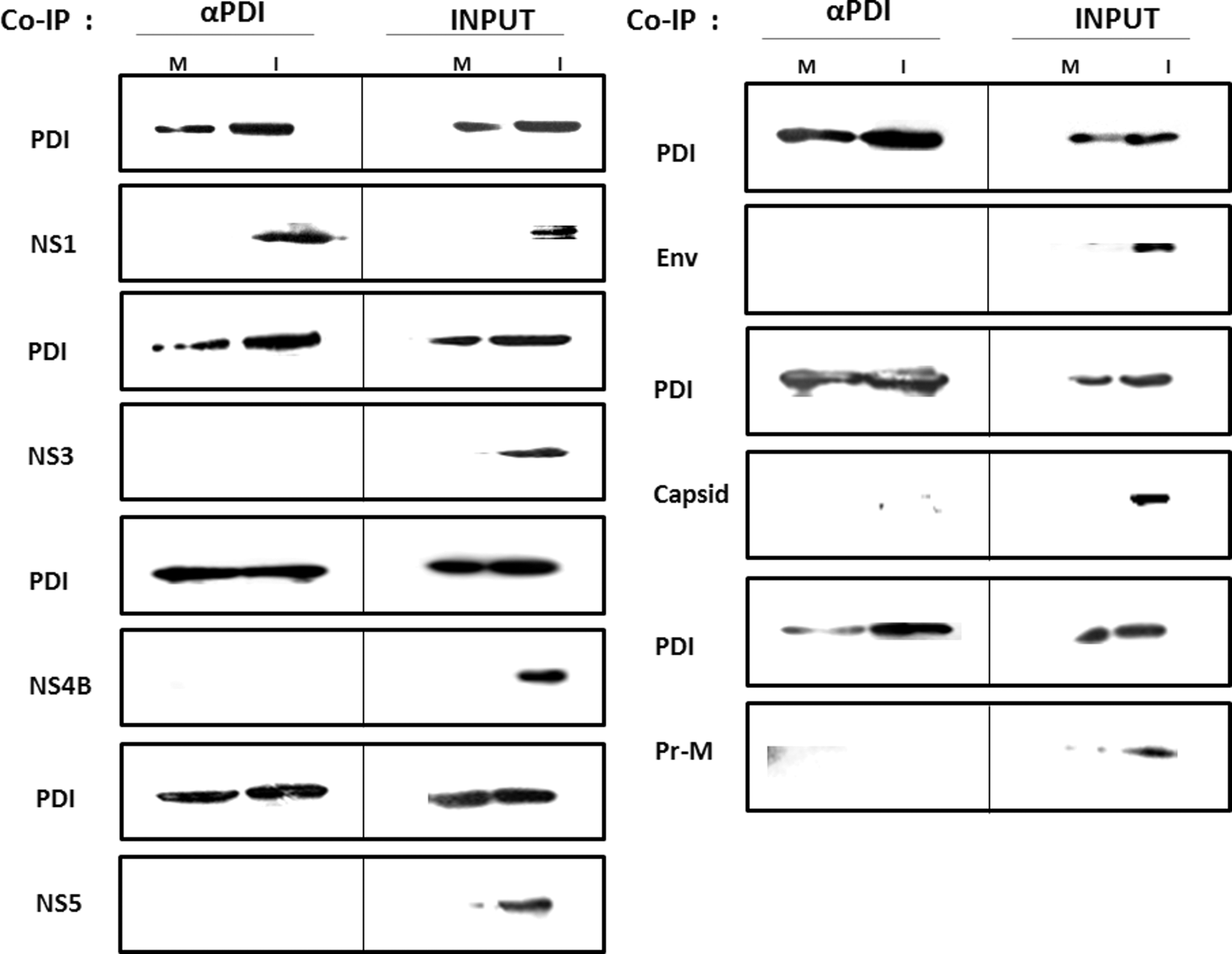
Co-immunoprecipitation (Co-IP) and immunoblotting. The results are shown of the screening of protein disulfide isomerase (PDI) against seven dengue virus (DENV) proteins: NS1, NS3, NS4B, NS5, Env, Capsid, and prM. αPDI, DENV proteins interacting with PDI were subjected to immunoblotting for both PDI and DENV protein simultaneously. M, Co-IP eluted proteins from mock cell lysate. I, Co-IP eluted proteins from DEN-infected cell lysate. INPUT: whole cell lysate of M-mock cells, I-DENV infected cells, 48 h post-infection.
PDI localized on lipid rafts interacts with the viral NS1 protein on host cell surface
The regulation of PDI expression, localized in lipid rafts, during DENV infection in THP-1 cells was investigated. PDI was found to co-localize with lipid rafts on the cell surface of both mock (C and Merge; Fig. 2A) and DENV-infected cells (CV and Merge; Fig. 2A). However, their expression was more pronounced in the latter, indicating that DENV infection upregulated the expression of PDI and lipid rafts. Lipid raft expression was disrupted using the inhibitor Methyl β Cyclodextrin (30 mM MβCD), which showed decreased expression of lipid rafts as well as PDI on the cell surface (CI and Merge; Fig. 2A). When MβCD-treated cells were infected with DENV, 24 h post-infection, expression of PDI on the cell surface increased along with the lipid rafts (CIV and Merge; Fig. 2A). DENV-infected cells alone (CV and Merge) and MβCD-treated and DENV-infected cells (CIV and Merge) both showed a similar increase in the expression of lipid rafts and PDI. Taken together, these results indicate that the disruption of lipid rafts resulted in decreased expression of PDI on the cell surface. However, when MβCD-treated cells were infected with DENV, the expression of these lipid rafts increased, and PDI expression increased simultaneously.
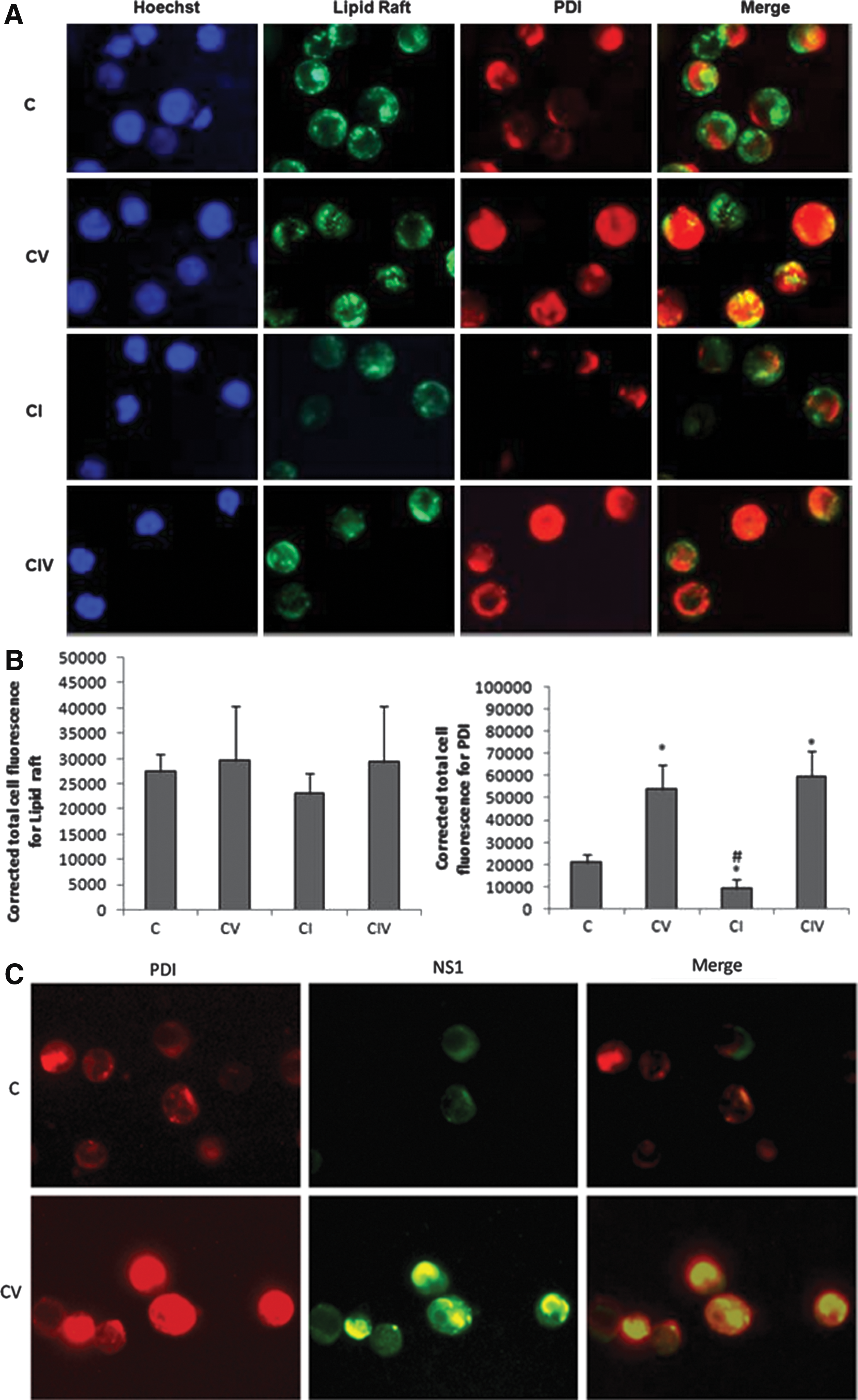
Furthermore, fluorescence image of mock- and DENV-infected cells immunostained for cell surface bound PDI and NS1 using PDI- PE labeled antibody and NS1- FITC labeled antibody were obtained. The FITC and PE channels were merged, and it was found that surface-bound PDI and NS1 co-localized in DENV-infected cells (CV and Merge; Fig. 2B). The mock cells (C and Merge; Fig. 2B) only showed PDI expression. Thus, taken together, these results suggest that DENV infection increases the expression of PDI localized on cell surface lipid rafts, and this PDI also interacts with the viral NS1 protein.
The role of lipid rafts in DENV infection and entry in THP-1 cells
Immunoblot analysis showed that the expression of DENV NS1 decreased on treating THP-1 cells with MβCD followed by DENV infection compared to DENV-infected cells alone (Fig. 3A). Moreover, cell-binding assay showed that the median fluorescence intensity of DENV-infected cells alone and MβCD-treated and DENV-infected cells was 71.69 and 35.87, respectively (Fig. 3B). Thus, it is reported that inhibition of lipid rafts decreased DENV cell surface binding in monocytes.
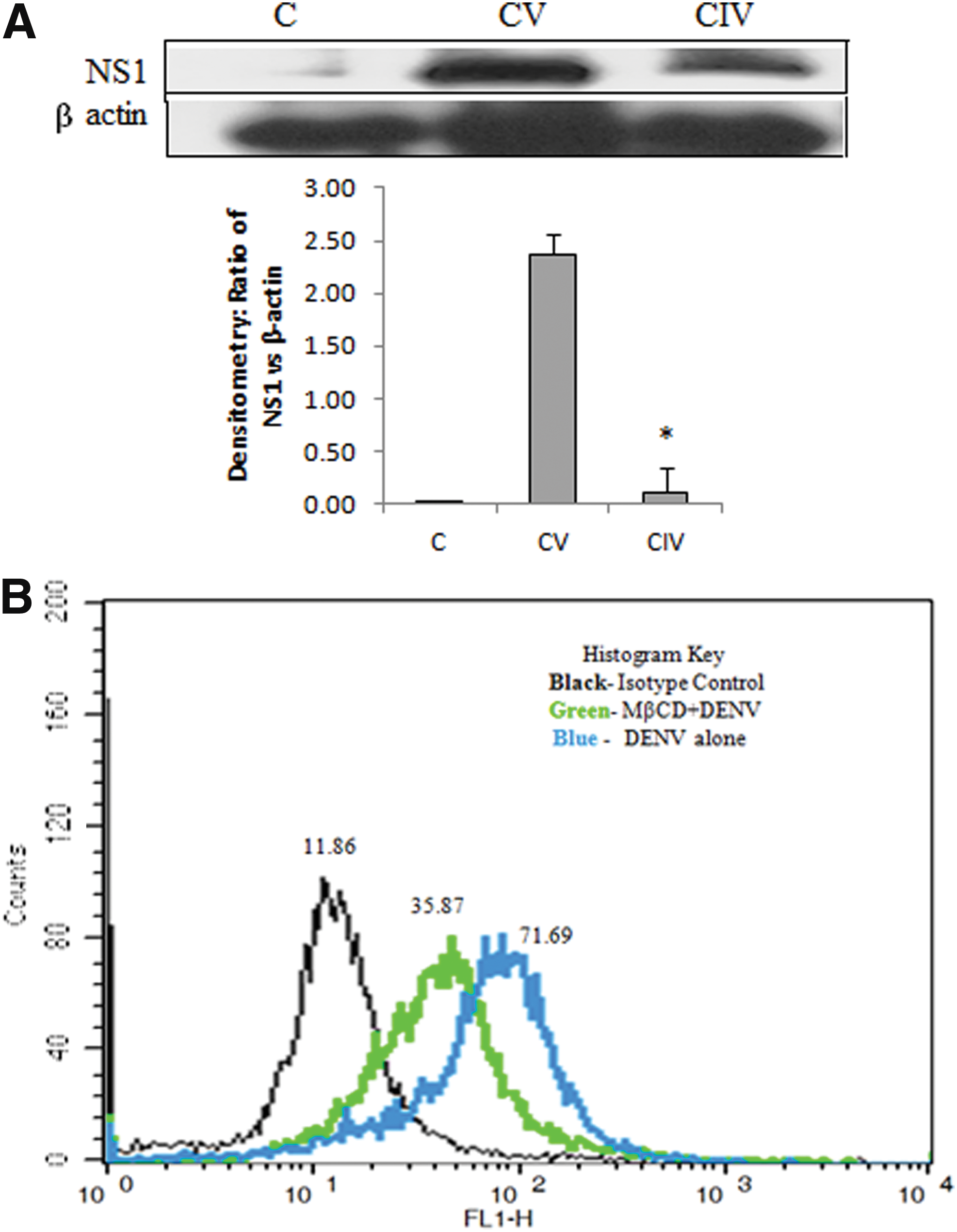
Role of PDI in DENV infection and entry in THP-1 cells
THP-1 cells were treated with the PDI inhibitor Bacitracin (100 μg/mL), and inhibition was confirmed by immunoblotting in the whole cell extract (Fig. 4). Moreover, the viral Env protein was inhibited in Bacitracin-treated and DENV-infected cells compared to DENV-infected cells alone. PDI thus plays an important role in DENV infection.
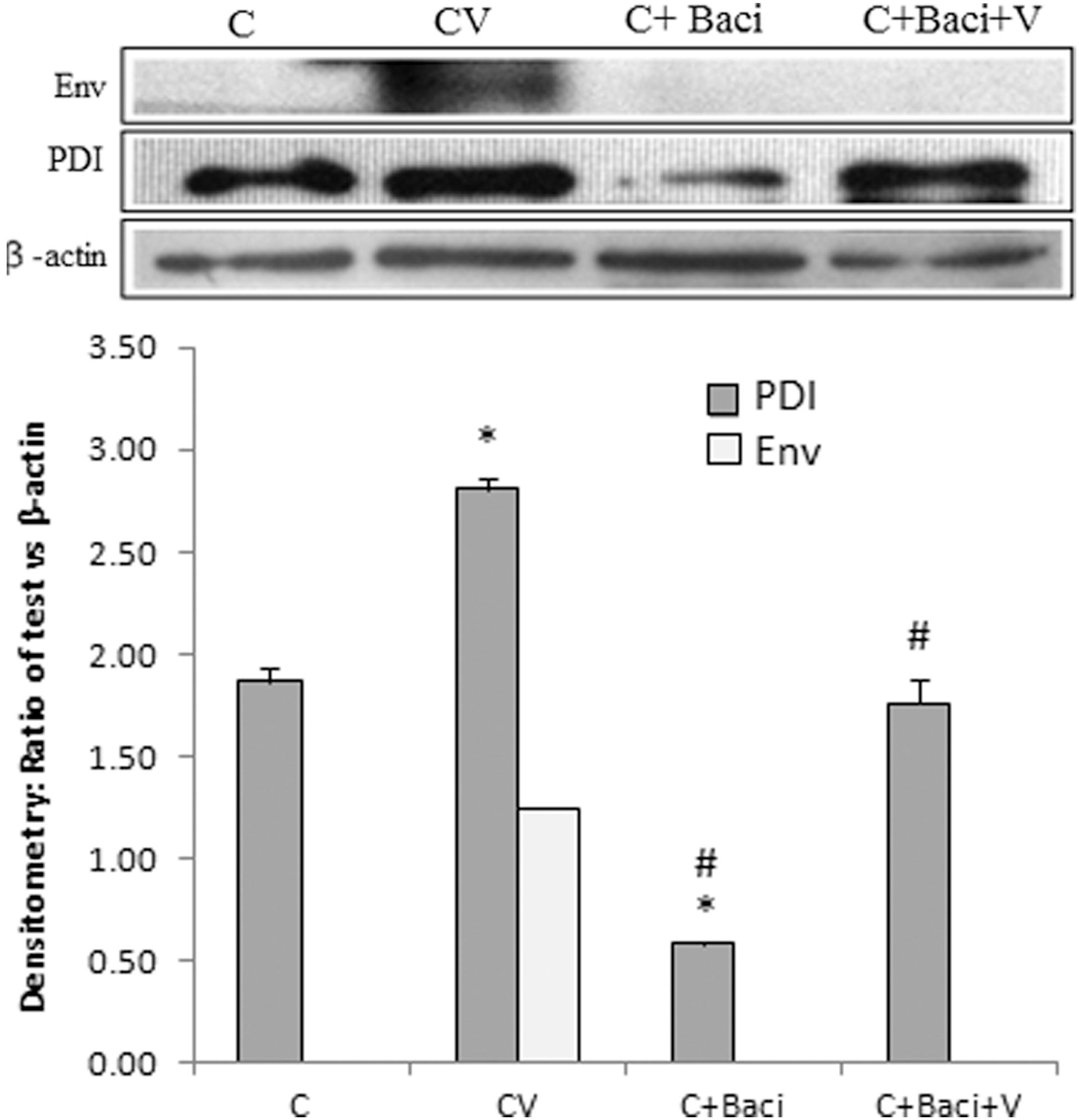
Immunoblot for Env and PDI: expression of Env was found to be inhibited in Bacitracin-treated and DENV-infected cell lysate, 48 h post-infection. Mock cells (C), DENV-infected (CV), Bacitracin-treated (C+Baci), and Bacitracin-treated and DENV-infected (C+Baci+V) cells. *p<0.05 vs. C; #p<0.05 vs. CV.
Immunofluorescence imaging of cells treated with Bacitracin showed decreased intracellular expression of PDI (CI-PDI Panel; Fig. 5). However, Bacitracin-treated and DENV-infected cells (CIV-PDI Panel; Fig. 5) showed decreased intracellular expression of PDI and NS1 compared to DENV-infected cells alone (CV-PDI Panel; Fig. 5). Thus, PDI was found to co-localize with NS1 within the cell and inhibition of PDI expression led to the inhibition of DENV NS1 expression. Moreover, cell-binding assay showed median fluorescence intensity 71.69 in DENV-infected cells, while Bacitracin-treated and DENV-infected cells showed an intensity of 38.20 (Fig. 6). Thus, it is reported that inhibition of PDI expression decreased DENV cell surface binding in monocytes.
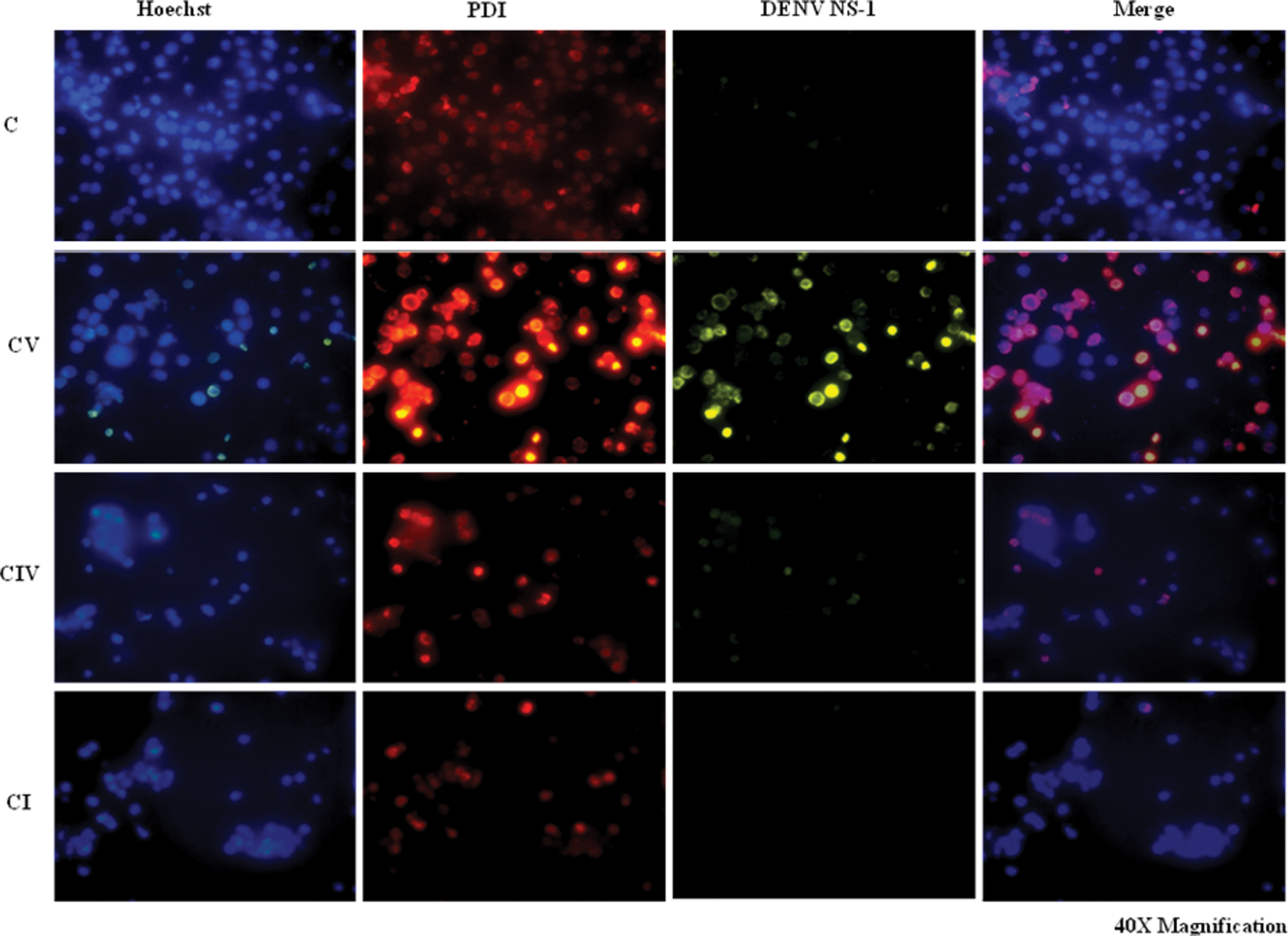
Intracellular co-localization of PDI and NS1: mock cells (C), DENV-infected (CV), Bacitracin-treated (CI), and Bacitracin-treated and DENV-infected (CIV) cells were immunostained for PDI (PE labeled), NS1 (FITC labeled) and Hoechst (nuclear stain, blue). DENV NS1 and PDI co-localize intracellularly. Color images available online at
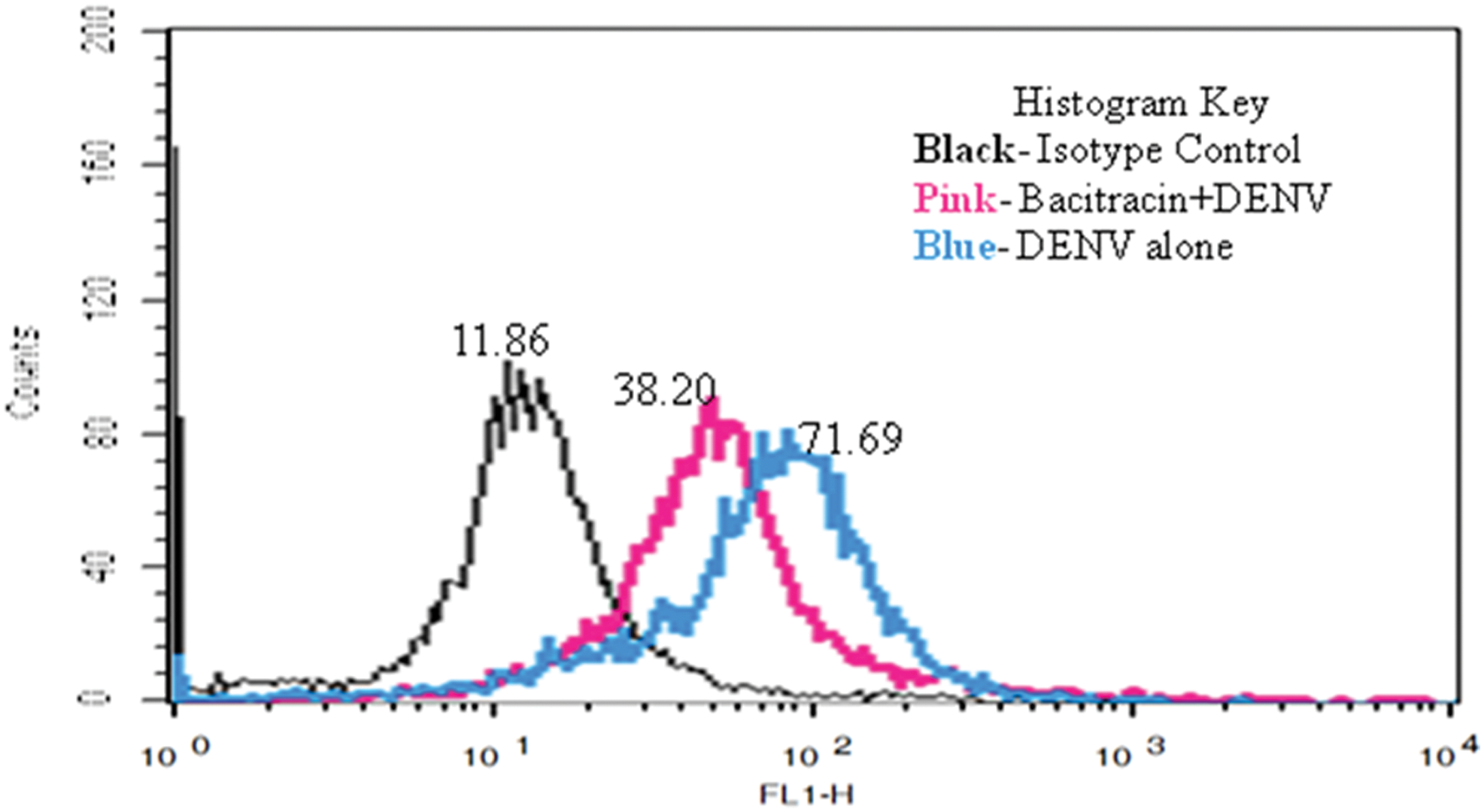
DENV binding to THP-1 cell surface after PDI Inhibition: mock- and DENV-infected cells with and without Bacitracin treatment were harvested, and immunostaining of cells was done using anti-DENV envelope primary antibody and IgG-FITC secondary antibody. Median fluorescence intensity of cells 80 min post-infection on ice has been shown. Color images available online at
Discussion
The present study identifies PDI as a key mediator of DENV infection in human monocytic cell line THP-1. Monocytes (MO)/macrophages (MΦ) are the prime targets of DENV, both in vivo and in vitro (11,12). The results of this study demonstrate that PDI interacts with the viral NS1 protein. The results confirm and provide further understanding of the previous findings that PDI, which is primarily involved in protein folding, is upregulated during DENV infection and is also proviral in nature (22). The viral NS1 protein can be found to be associated with intracellular membranes and the cell surface (10). PDI is localized in the lipid raft domain on the cell surface (3). It has been observed that PDI co-localizes with lipid rafts on the THP-1 cell surface. Disruption of these lipid rafts using MβCD decreased the expression of PDI along the cell surface. However, when MβCD-treated cells were infected with DENV, the expression of PDI increased along with lipid rafts. DENV NS1 protein has been shown to interact with the host cellular lipid bilayer, especially lipid rafts, and uses it to make secretory forms of NS1 for further signaling within the host (2,25). The results of this study show that DENV infection increases the expression not only of cell surface lipid rafts but also of the host chaperone PDI. These results are in line with others (27,34). Moreover, the cell surface PDI was found to co-localize with the viral NS1 protein. Taken together, the present study suggests that PDI localized on cell surface lipid rafts interacts with the viral NS1 protein during DENV infection.
Intracellular DENV protein NS1 expression was also decreased in MβCD-treated and DENV-infected cells. Moreover, MβCD treatment also decreased the DENV binding to the host cell surface. This suggests that cholesterol inhibition decreased DENV infection as a direct implication of inhibition of virus entry into the cells. Another study suggests that the cholesterol rich lipid-raft microdomains play an important role in the viral polyprotein processing and replication, since it was found that the viral NS3 and NS2B proteins interacted with the lipid raft marker caveolin (9). The presence of cholesterol is required for the activation of signaling molecules Jun NH(2)-terminal kinase (JNK) and p38 in human macrophages, which are important for establishing DENV infection in the host cells (4). Intact lipid rafts have also been shown to be necessary for mediating antibody-dependent enhancement of DENV infection in monocytic cells (29). Moreover, DENV has been shown to increase the intracellular cholesterol levels in Huh-7 cells early during infection by modulating the expression of low density lipoprotein receptor (31). DENV therefore was found to upregulate and use the lipid metabolism of the cells to its advantage, resulting in increased expression of lipid rafts and the PDI in its association. The present study is suggestive of the fact that if PDI or cholesterol inhibition is done for a transient period during DENV disease it may bring about a reduction in the viral load. Another strategy for a therapeutic target could be to inhibit the host PDI and DENV NS1 interaction for decreasing DENV multiplication.
Inhibition of PDI in DENV-infected cells led to the inhibition of the viral Env protein. Moreover, PDI has been shown to co-precipitate with DENV Env protein in C6/36 cells (32). Another consequence of this was that the viral NS1 protein was also inhibited, which was found to co-localize with PDI within the cell. Furthermore, PDI was inhibited using Bacitracin, which has been reported to enter the cells and has been found to associate with various cellular components, including caveolin, clathrin light chain, and LAMP1 using pulse-chase experiment (35). Bacitracin treatment decreased the binding of DENV to the host cell surface, indicating that PDI may have a role even in virus entry into the cell besides helping the virus to multiply within. However, DENV adopts different routes of entry in different types of cells (1,19,30). The DENV Env protein has been suggested to participate in viral entry into the host cell (13,24). Although it has been observed that PDI has a role in virus entry, DENV Env was not found to co-precipitate with PDI by co-immunoprecipitation. This may be because the Env protein is probably bound to some other host protein during viral entry or, in other words, the DENV Env could be in less proximity to PDI. These results suggest that PDI is a key cellular protein that helps the virus to multiply and also gain entry into the cell. Several studies have shown interactions or signaling by NS1 indirectly via PDI. The viral NS1 protein has been shown to mimic PDI, since anti-DENV NS1 antibodies were found to cross-react with PDI (3,4). Antibodies directed against DENV NS1 protein has been suggested to upregulate heme oxygenase 1 by interacting with PDI in the human umbilical vein endothelial cells (14). Taken together, this suggests that PDI inhibition may be an effective strategy to decrease DENV infection and the ensuing pathogenesis it causes. Taken together, the results of this study show that the host protein, PDI, has a vital role in DENV multiplication and its entry into the cell, and its inhibition could be an important strategy to decrease the viral load and in controlling DENV infection.
Footnotes
Acknowledgments
This project was supported by the Defence Institute of Physiology and Allied Sciences, Defence Research and Development Organization (DRDO), Ministry of Defence, India, in the form of TASK-177 and is gratefully acknowledged. D.D. received financial support in the form of INSPIRE fellowship from Department of Science and Technology, government of India.
Author Disclosure Statement:
No competing financial interests exist.