
Abstract
Nowadays, adjuvant is still important for boosting immunity and improving resistance in animals. In order to boost the immunity of porcine circovirus type 2 (PCV2) DNA vaccine, CpG motifs were inserted. In this study, the dose–effect was studied, and the immunity of PCV2 DNA vaccines by recombinant open reading frame 2 (ORF2) gene and CpG motifs was evaluated. Three-week-old Changbai piglets were inoculated intramuscularly with 200 μg, 400 μg, and 800 μg DNA vaccines containing 14 and 18 CpG motifs, respectively. Average gain and rectum temperature were recorded everyday during the experiments. Blood was collected from the piglets after vaccination to detect the changes of specific antibodies, interleukin-2, and immune cells every week. Tissues were collected for histopathology and polymerase chain reaction. The results indicated that compared to those of the control piglets, all concentrations of two DNA vaccines could induce PCV2-specific antibodies. A cellular immunity test showed that PCV2-specific lymphocytes proliferated the number of TH, TC, and CD3+ positive T-cells raised in the blood of DNA vaccine immune groups. There was no distinct pathological damage and viremia occurring in pigs that were inoculated with DNA vaccines, but there was some minor pathological damage in the control group. The results demonstrated that CpG motifs as an adjuvant could boost the humoral and cellular immunity of pigs to PCV2, especially in terms of cellular immunity. Comparing two DNA vaccines that were constructed, the one containing 18 CpG motifs was more effective. This is the first report that CpG motifs as an adjuvant insert to the PCV2 DNA vaccine could boost immunity.
Introduction
P
Vaccination remains the single most valuable tool in the prevention of infectious disease. At present, most research has been carried out from the construction of chimeric viruses, subunit vaccines, recombinant vaccines, and genetic engineering vaccines. Many PCV2 vaccine approaches have been tested under experimental conditions in mice and/or pigs, including DNA vaccines (10,11,16,17,30), a chimeric baculovirus expressed cap protein (24), an ORF2 subunit vaccine (6), a pseudorabies virus expressing ORF1-ORF2 vector vaccine (15), an adeno-ORF2 expressing vector vaccine, and live or inactivated attenuated chimeric PCV2-1 vaccines (12). The most successful vaccine candidates were those based on the induction of an active immune response against the capsid protein of PCV2.
CpG motifs were dinucleotides flanked by two 5′ purines and two 3′ pyrimidines. These motifs could rapidly trigger an innate immune response, characterized by the production of interleukin-6 (IL-6), IL-12, and interferon-γ (IFN-γ). CpG motifs in the plasmid backbone of DNA vaccines play an important role in the induction of antigen-specific immunity (14,18 –21).
In this study, based on the earlier research in the authors' laboratory (Cheng KH, “Study on Series of DNA Vaccines Against Porcine Circovirus Type 2[D],”[master‘s thesis, QingDao Agricultural University, 2009]). DNA vaccines with better immunizing effect containing 14 and 18 CpG motifs were screened for further study to evaluate the immune enhancement effect of DNA vaccine.
Materials and Methods
Animals
Forty Changbai piglets aged 3 weeks old that tested negative for PCV2 antigen-antibody were selected randomly. The animal study proposal was approved by the Institutional Animal Care and Use Committee (IACUC) of the Shandong province. All experimental procedures involving the piglets were performed in accordance with the Regulations for the Administration of Affairs Concerning Experimental Animals approved by the State Council of People's Republic of China.
Viruses and vaccine
DNA vaccine plasmids 14CpG-pVAX1-ORF2 and 18CpG-pVAX1-ORF2 were constructed by Cheng Kaihui (8), PCV2 strain SD2 (TCID50=106.0) were saved by Shandong Key Laboratory of Animal Disease Control and Breeding. Mouse anti-pig CD4α and mouse anti-pig CD3ɛ were obtained from Southern Biotech. PE mouse anti-Pig CD8α and PE-Cy5 streptavidin were purchased from BD Pharmingen. The porcine interleukin-2 (IL-2) enzyme-linked immunosorbent assay kit (ELISA) was obtained from R&D. The CpG motifs sequences of eukaryotic vectors CpG-pVAX1-ORF2 are shown in Table 1.
Note: The underlined letters were the insertion CpG-ODN motif sequences; letters that are boldface but not underlined between the CpG-ODN motif are protective base.
CpG-ODN, CpG-oligodeoxynucleotide; ORF, open reading frame.
Animal vaccination
Forty 3-week-old piglets were randomly divided into eight groups (A–H), five piglets in each group. The piglets were vaccinated intramuscularly (i.m.) in the neck. Two weeks later, piglets were boosted individually. On day 42, four piglets, selected from groups A, B, E, and H individually, were euthanized. On day 58, animals were inoculated with 1.0 mL of minimal essential medium containing 106.0TCID50 of PCV2 strain SD2. On day 77, the remaining piglets were euthanized. A summary of the experimental design is shown in Table 2.
Physical signs studies
Body weight was recorded for all piglets at the time of first and second vaccinations, and at weekly intervals from the day of challenge onwards. Rectal temperatures were recorded every day.
Immunological assays
Assay of PCV2 detection
Two pairs of polymerase chain reaction (PCR) primers were designed using computer software (Primer Premier 5.0) according to the published sequence of the PCV2 in GenBank (HQ148879) to establish a fluorescent quantitative PCR assay for the detection of PCV2. One set of primers (YG1: 5′-TTA GTC TTC CAA TCA CGC TTC TGC-3′; YG2: 5′-ACG GAG TGA CCT GTC TAC TGC TG-3′) was designed to amplify a fragment of 198 bp from ORF2 of the PCV2 and the other set of primers (Y1: 5′-GCT GAT TTC TTT TGT TGT TTG GTT-3′; Y2: 5′-GAG TGG GCT CCA GTG CTG TTA T-3′) was designed to amplify a fragment of 196 bp from the ORF1 of PCV2. The amplification was carried out in a 25 μL reaction containing 1 μM of each forward and reverse primer, 12.5 μL of SYBR Green I Mix (TaKaRa), 2 μL DNA templates, and ddH2O up to 25 μL. The PCR conditions were as follows: 95°C for 10 min, followed by 30 cycles of amplification at 95°C for 10 sec, and 62°C for 10 sec. Melt curve analysis was performed at 95°C for 2 min, 60°C for 20 sec, and 95°C.
Assay of specific antibody against PCV2
Blood samples were collected upon the piglets' arrival at the research facility and weekly thereafter until necropsy. The serum was tested with the PCV2 ELISA kit (J.B.T.). Samples were considered positive if the calculated S/P ratio was ≥0.4.
Assay of IL-2 by sandwich ELISA
Blood samples were also tested by IL-2 ELISA kit (R&D). The absorbance of each well was read in a spectrophotometer at 450 nm. Serial dilutions of standard samples were used to quantify the contents, and served as a standard curve. Then IL-2 contents in the serum samples were calculated.
Assay of CD3, CD4, and CD8+ T-cells by flow cytometry
The lymphocytes of blood samples (106 cells) were resuspended in 1 mL Hank's Balanced Salt Mixture and incubated with 1 μL FITC-conjugated mouse anti-pig CD4α (Southern Biotech), 2 μL PE-CY5-conjugated CD3ɛ (BD Biosciences), and PE-conjugated mouse anti-pig CD8α (BD Biosciences) at a concentration of 1 mL per 106 cells in an ice bath. After 20 min incubation, the cells were washed and analyzed using flow cytometry (Beckman Coulter).
Pathological and histopathology studies
The tissues of the remaining piglets were collected on day 77 after being euthanized and processed using conventional histopathological methods. Briefly, the tissues were fixed in 10% neutral-buffered formalin solution, and were sectioned and stained with hematoxylin and eosin. Microscopic changes were determined by comparing the tissues of the challenged piglets with those of the control group.
Statistical analysis
All data are presented as means±SD with SPSS v13.0 software (SPSS, Inc.). Statistical analyses were performed by two-way analysis of variance (ANOVA) followed by S-N-K post hoc tests individually. The level of significance was set at p<0.05.
Results
Average daily gain
Body weight was recorded from all piglets on days 1 and 58, and average daily gain (ADG) was calculated. Compared to the control piglets (groups A and B), the ADG of the immunostimulated piglets increased. However, there was no significant difference among groups (p≥0.05). The weight gain was not altered by vaccination.
Temperatures of piglets
Rectal temperatures were recorded every day. All temperatures were <40.5°C. The vaccination is safe, the physiology of the piglets was stable, and the temperatures were not altered by vaccination.
Viremia
Each time, PCV2 was detected by quantitative fluorescence with two pairs of primers for ORF1 and ORF2. On day 42, four piglets, selected from groups A, B, E, and H individually, were euthanized, and no PCV2 genome was detected in any of their tissues, indicating that there was no PCV2 infection during the vaccination. Then, on day 58, animals were inoculated with 1.0 mL of essential medium containing 106.0TCID50 of PCV2 strain SD2. PCV2 could be detected in the blood of all piglets on day 63. On day 77, all piglets were euthanized, and PCV2 could be detected by quantitative fluorescence PCR in all selected tissues. PCV2 could be detected in the blood and all tissues simultaneously in groups A and B, but could not be detected in all blood simultaneously in groups C–H.
Amount of specific antibody
All the piglets seroconverted between 0 and 14 dpi (Fig. 1), and all groups decreased after 14 dpi except group H. The PCV2-antibody response of group H increased to its highest level at 28 dpi and was still higher than other groups during vaccination. At 58 dpi, inoculated with 1.0 mL of minimal essential medium containing 106.0TCID50 of PCV2 strain SD2, the antibody level was increased to a different degree due to the anamnestic response in the vaccinated groups and because virus challenge in the control groups and group H was kept at a higher level throughout the study.
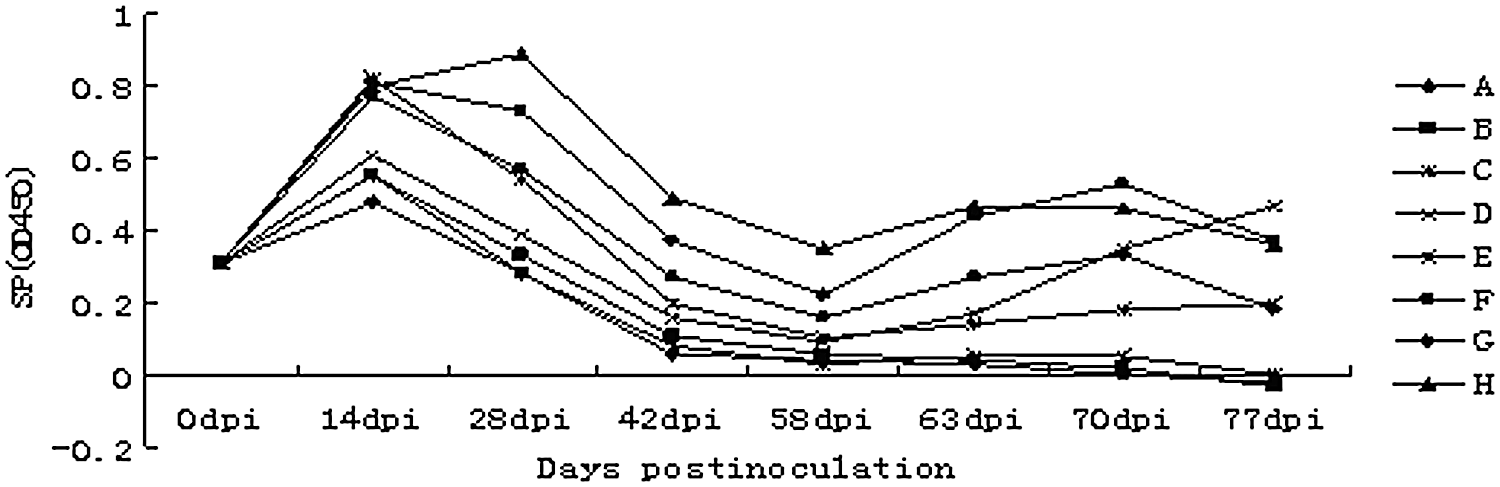
Porcine circovirus type 2 (PCV2)-antibody responses monitored by PCV2–ORF2 protein enzyme-linked immunosorbent assay. The positive cutoff was set at SP ≥0.4 where SP=(OD of sample – OD mean of negative control)/(OD mean of positive control – OD mean of negative control). The antibody of group H is higher than other groups throughout the time after vaccination. During the first week after injection, it remains higher and then gradually declines. OD, optical density.
Changes of IL-2 content in piglets
The IL-2 contents in the serum samples were calculated after serial dilutions of standard samples were used to serve as a standard curve. The IL-2 contents of the four groups (E, F, G, and H) were on the rise after 14 dpi (Fig. 2). After being injected with PCV2, the contents of most groups showed an upward trend in IL-2 levels, reached a peak at 70 dpi, and then gradually decreased. As shown in Figure 2, the immune effect of group H was better than the immune effect of the other groups.
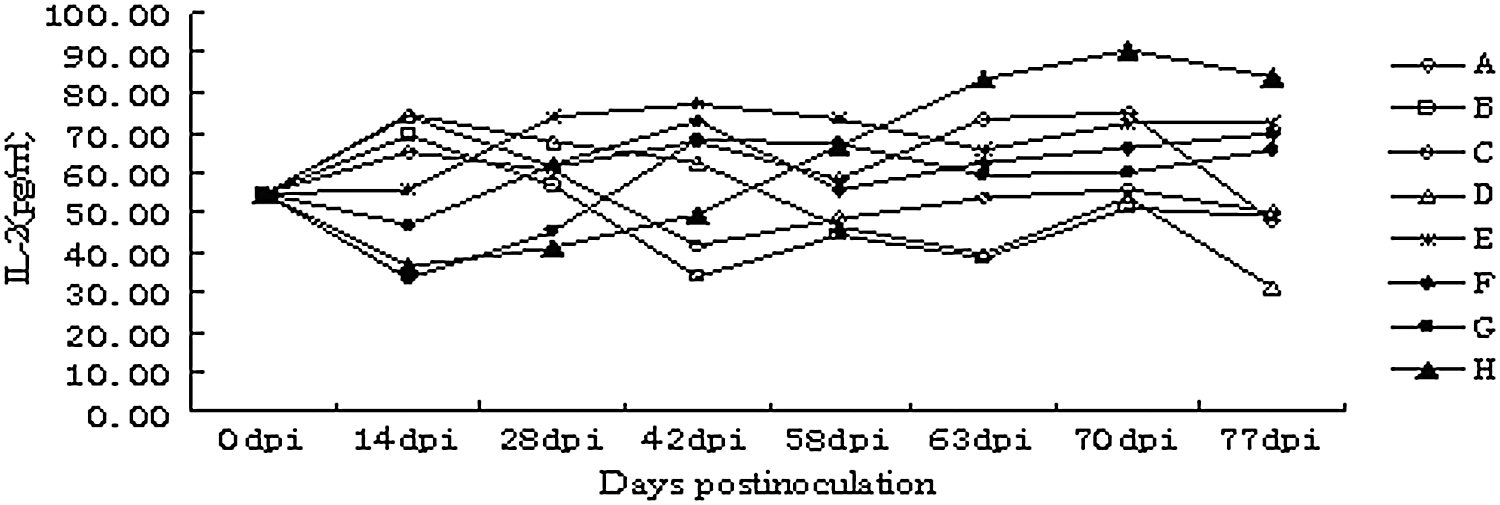
Level of interleukin-2 (IL-2) content in peripheral blood. On day 58, animals were inoculated with 1.0 mL of essential medium containing 106.0TCID50 of PCV2. The IL-2 level in the sera of group H differed significantly from the other groups since 58 days after injection (p<0.05), and then gradually declined.
Cellular immune responses
The lymphocytes in the blood were isolated at 2 and 3 weeks post-infection. CD3+, CD4+, and CD8+ T-cells were counted and analyzed using flow cytometry. Two weeks after injection, compared with the control groups, CD3+ T-cells, CD4+ TH cells, and CD8+ TC cells significantly increased in the blood of the treated groups C–H, which indicated that the vaccine with 14 and 18 CpG motifs can stimulate lymphopoiesis.
Clinical and pathological outcomes
To investigate the protective efficacy of different vaccine protocols, clinical symptoms and pathological changes were compared in the challenged piglets. No clinical signs or gross lesions were observed in the vaccinated groups. In contrast, predominant histological lesions in the control groups included depletion of lymphoid tissues and lymphohistiocytic to granulomatous inflammation in the lung, kidney, and lymphoid tissues (Fig. 3). Microscopic lesions included lymphoid depletion, necrotizing hepatitis, and mild necrotizing bronchiolitis (Fig. 4). Their alveolar was atrophied, and inguinal lymphadenopathy was observed, as well as gastric ulceration.
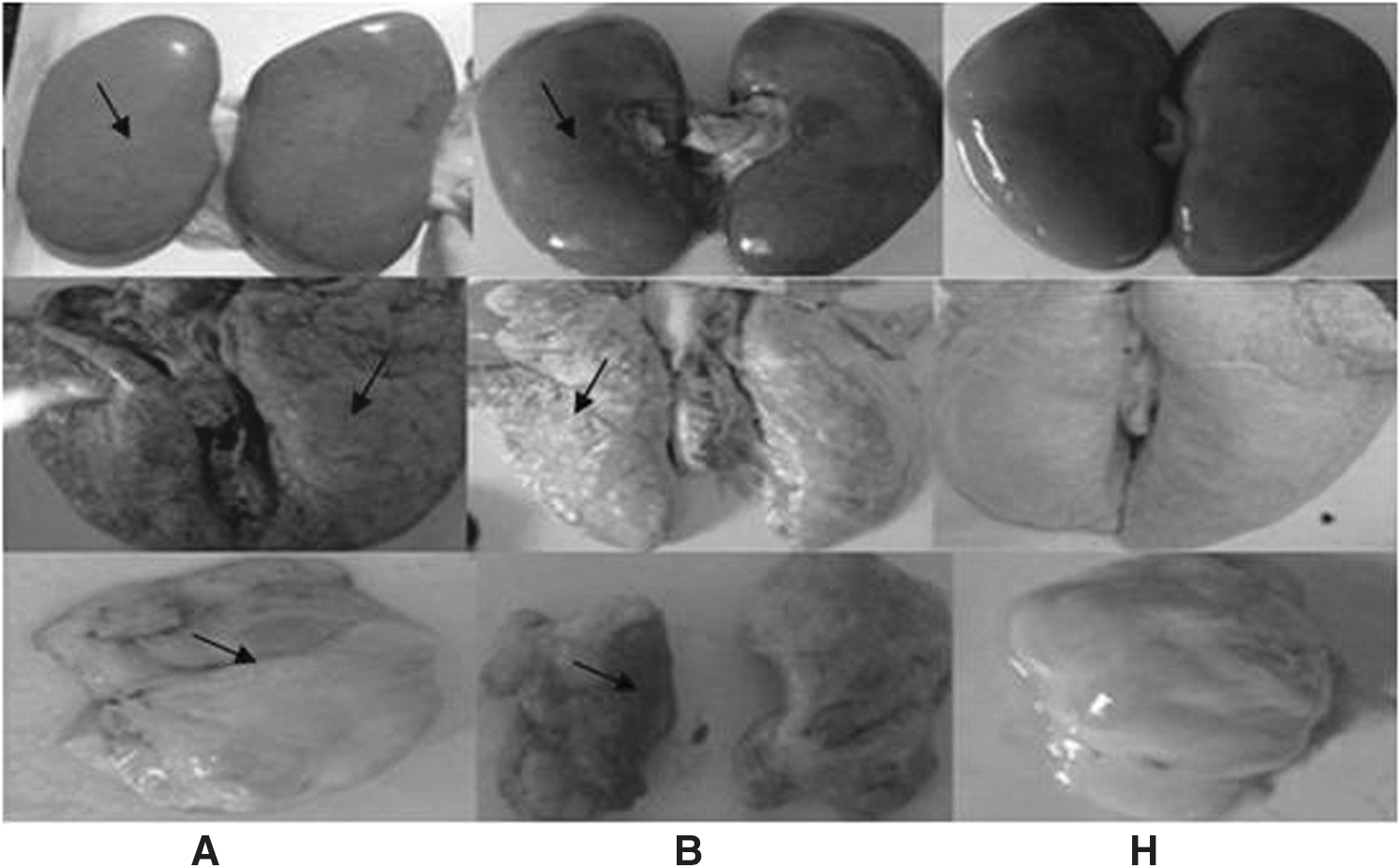
Pathological lesions of organs from control groups and immunostimulated groups. Groups
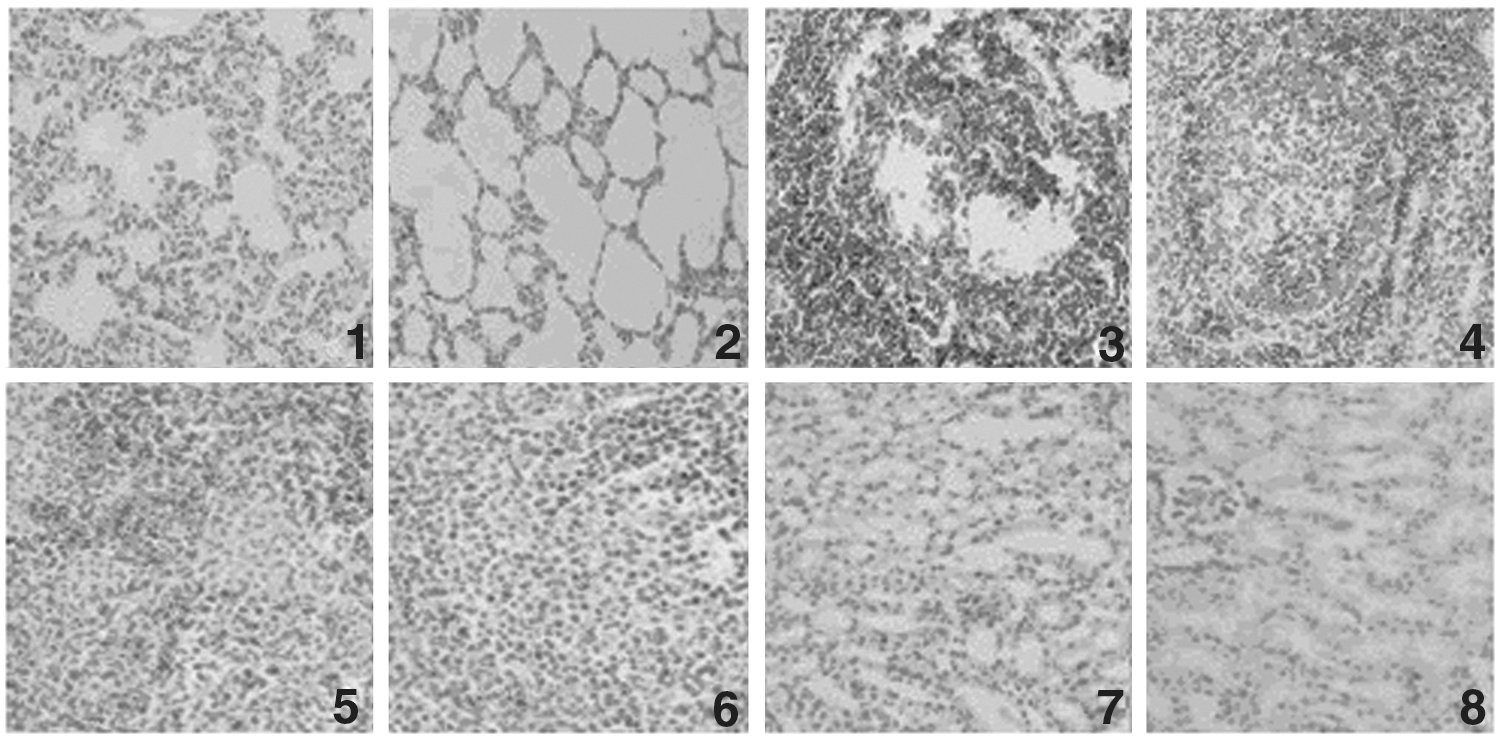
Microscopic sections of organs from control groups and immunostimulated groups (×50).
Discussion
The oligodeoxynucleotide (CpG-ODN) containing cytosine-guanine (CpG) motifs can, in the presence of antigens, enhance antigen-specific humoral and cellular immunity. However, in the absence of antigens, it can enhance nonspecific immunity (20,21). In the vaccine development process, the application of CpG-ODN adjuvant can induce specific immune cell differentiation and proliferation, and enhance the immunogenicity of the vaccine (14,18). In addition, this can induce a Th1-type immune response to increase humoral and cellular immunity (9). However, as CpG-ODN in vivo degrades by nuclease very quickly, high-dose CpG-ODN should be given to demonstrate its effectiveness, so the plasmid containing CpG-ODN can express for a long time in animal body to play its role in immune enhancement and immune regulation.
CpG motifs showed species specificity, and the length of CpG motifs between different species were not the same (31). It was reported that 5–20 additional CpG motifs were cloned into a pUC-derived plasmid, and the BALB/c mice were immunized with the plasmid. The results showed plasmids containing 20 CpG copies were the most effective immune enhancers both in vitro and in vivo (22).
Xu honglin inserted varieties of CpG motifs with immunostimulation to human and mice into plasmid backbone, and finally they had got plasmid contained 0, 9, and 26 CpG motifs separately. The results showed that IgM secretion in mice increased with the increase in CpG motifs, but the most IFN-γ content was generated by nine CpG motifs. These results suggest that the CpG motifs enhanced the immunogenicity of the DNA vaccines, but not the more, the better. Krieg's experiments showed DNA with the introduction of 50 CpG motifs could maintain the strongest CTL response but the antibodies induced were greatly reduced (23).
Based on earlier research of the immunogenicity effects of DNA vaccines containing series CpG motifs (from 1 to 24) in pigs, DNA vaccines were further studied on screening the better number of CpG motifs between 14 and 18.
Since PCV2 is an immunosuppressive disease, in the case of artificial reproduction, the results were different in different countries, which may be related to virulence, the age of the animal, and maternal antibodies. In previous studies, BALB/c or Kunming mice were used as infection models for PCV2 infection (17,25). Clinical PMWS had been reproduced in gnotobiotic pigs co-infected with PCV2 and porcine parvovirus (PPV) (4,13). However, no clinical PMWS was found in pigs infected with PCV2 alone (4,5). In the present study, 3-week-old Changbai piglets that tested negative for PCV2 antigen-antibody were selected for experiments. Through the experiment, the temperature of both the vaccination groups and the control groups fluctuated, but no piglets had a temperature >40.5°C. No groups had any significant ADG difference. The results showed that vaccination is safe. After PCV2 strain SD2 injection, the temperature of the control groups (A and B) was >40.5°C, while the temperatures of the vaccine groups (C–H) were still <40.5°C. There was also a significant ADG difference in the vaccine groups (C–H). Three weeks after PCV2 strain SD2 injection, although there were no clinical manifestations in the control group, lesions were found to varying degrees in the tissues (lung, kidney, and spleen). At the same time, viral DNA was detected in the blood and tissues of the control groups. In contrast, no visible lesions were detected in the tissues of groups C–H. Compared to the non-immunostimulated control piglets (A and B), the ADG of the immunostimulated pigs was increased, although there was no significant difference among groups (p≥0.05).
PCV2 is a virus that can affect the respiratory system and be transmitted oronasally. Before PCV2 injection, four piglets from four groups were tested, and no infection with wild strain of PCV2 was found. After injection, PCV2 could be detected in the blood and the viremia of the piglets. Three weeks later, the DNA vaccines neutralized the PCV2 virus, and the viremia disappeared.
In order to test the PCV2-specific antibody, an ELISA test was performed on the serum of these immunostimulated and non-immunostimulated control piglets to detect the PCV2-specific antibody. The results revealed that the two vaccines had good immunogenicity, without the influence of maternal antibody, and could quickly produce high antibody levels against viral infection. There was no PCV2-specific antibody detected in non-immunostimulated control piglets. The variation in antibody levels in this experiment was the same as the result of DNA vaccine and a chimeric vaccine (16,12).
IL-2 was identified based on its potent T-cell growth-factor activity and is widely considered to be a key cytokine in T-cell-dependent immune responses (27). In the present study, IL-2 concentration in the peripheral blood of experimental animals was detected and was found to increase after immunization, gradually return to normal levels, and then rapidly increased after injection. Early research showed that CD8+ T-cells play a crucial role in protecting against PCV2 infection and the protective immunity induced by the pORF2 plasmid is superior to that induced by the PCV2 Cap protein (32). In the FCM analysis, the proliferation of CD4+ and CD8+ lymphocytes after injection was tested. The CD4+/CD8+ ratio increased and then gradually returned to normal levels. In all of the above results, the immunoprotection of DNA vaccine containing 18 CpG motifs repeated was stronger than vaccine containing 14 CpG motifs repeated.
Conclusions
In summary, PCV2 DNA vaccines containing CpG motifs could build up resistibility and reduce immune organ damage by means of humoral immunity and cellular immunity. These vaccines decreased the incidence of PMWS effectively. Comparing the two DNA vaccines that were constructed, the one containing 18 CpG motifs was more effective. This is the first report that CpG motifs as an adjuvant insert to the PCV2 DNA vaccine could boost immunity.
Footnotes
Acknowledgments
The study was partly supported by the National Natural Science Funds (31201889, 31170146), Young Talents Training Program of Shandong Academy of Agricultural Sciences, Shandong Modern Agricultural Technology and Industry System.
Author Disclosure Statement
No competing financial interests exist.