
Abstract
Hepatitis B virus (HBV) is a leading cause of liver cirrhosis and hepatocellular carcinoma. The outcome of HBV infection is largely determined by the host immune response, with virus-specific cytotoxic T cells being able to mediate immunity against HBV as well as causing liver pathology. γδ T cells are reported to be depleted in patients with HBV-associated liver disease. However, it is not known if these cells control HBV infection in patients with asymptomatic chronic HBV infection. In this study, the frequencies, phenotypes, and interferon-γ production were examined by circulating γδ T cell subsets in a group of asymptomatic HBV carriers with low viral loads and little evidence of liver disease. It is shown that γδ T cells expressing Vδ1 and Vδ2 T cell receptors and effector-memory phenotypes are found at higher frequencies in these patients compared to controls. Vδ2 T cells from the patients expressed interferon-γ significantly more frequently than Vδ2 T cells from healthy donors in the absence of ex vivo stimulation. These data suggest that effector-memory IFN-γ-producing Vδ2 T cells may contribute to the control of HBV in patients with asymptomatic infection, without mediating liver pathology.
Introduction
H
Resolution of HBV infection is associated with strong, polyclonal, and multi-specific CD8+ cytotoxic T-lymphocyte (CTL) responses directed against multiple viral epitopes, while chronic HBV infection is characterized by lower numbers and lower potency of HBV-specific CTLs (38,43,46,55). Inefficient T cell priming by dendritic cells (21,40), immunomodulation by regulatory T cells (28,54), and clonal exhaustion due to upregulation of inhibitory receptors, such as PD-1 (27,59), have been implicated as factors that contribute to the inadequate T cell responses. Moreover, CD8+ CTL-mediated cytotoxicity is strongly implicated in HBV-related liver damage but does not appear to play a major role in eliminating the virus, while IFN-γ produced by virus-specific CTLs and natural killer (NK) cells are thought to mediate clearance of HBV by interfering with viral replication and by recruiting other antigen-nonspecific effector cells (13,31,36,38,55). Therefore, the ideal immune response against HBV must control viral replication but limit hepatocyte cytotoxicity and immune-mediated liver damage.
Roles for γδ T cells in antiviral immune responses have been reported for cytomegalovirus (17,32), Epstein–Barr virus (18), human immunodeficiency virus (HIV) (45), and herpes simplex virus (39) infections. γδ T cells have also been implicated in immune responses to hepatitis C virus (HCV) and are thought to play a role in liver injury associated with the virus (41,56). Sing et al. (50) reported that γδ T cells were expanded in the blood of patients with HBV infection who seroconverted. Subsequently, Chen et al. (10,11) reported that Vδ2 T cells, the most abundant subset of γδ T cells in human blood and liver (37), are depleted in patients with chronic HBV infection and in patients who develop HBV-associated acute-on-chronic liver failure (HBV-ACLF). In contrast, Vδ1 T cells, the most abundant γδ T cell subset in the intestine, were expanded in the blood of patients with HBV-ACLF. γδ from HBV-ACLF patients exhibited enhanced cytotoxicity and inflammatory cytokine production compared to their counterparts in chronic HBV patients and healthy controls, suggesting that γδ T cells play a role in liver injury in HBV-ACLF (10).
In the present study, the potential role of γδ T cell subsets in immunity against HBV in the absence of liver injury was investigated by studying a cohort of patients with persistent HBV infection (HBsAg-positive) but low viral burden (<20,000 copies/mL) and no evidence of liver disease (alanine aminotransferase [ALT] <70 IU/mL). This patient cohort can be considered as having an efficient immune response against HBV, which is under sufficient regulatory control and does not cause significant pathology but fails to eliminate the virus completely (34). The frequencies, differentiation status, and IFN-γ production of the Vδ1+ and Vδ2+ subsets of γδ T cells were examined to assess, for the first time, their potential roles in controlled asymptomatic HBV infection.
Materials and Methods
Subjects
Sixty-two consecutive patients (32 male; age range 18–60 years) persistently infected with HBV, attending the Hepatology Outpatient clinic at St. James Hospital, Dublin, were studied. The patients were ethnically diverse and comprised 27 Africans, 25 Caucasians, and 10 Asians. ALT levels ranged from 8 to 143 IU/mL, with 50 patients having ALT <40 IU/mL and 12 patients having higher ALT levels >40 IU/mL. Liver biopsy was performed for those patients with high ALT (>70 IU/mL) relating to HBV. Blood samples for the study were obtained at one time point for each patient, and no patients in the cohort were receiving treatment at the time of sample collection. The viral load was measured at the time of blood collection and varied from 7 copies/mL to 4.5×108 copies/mL, but was <20,000 copies/mL in the majority of patients. For the analysis, 100,000 copies/mL was chosen as the cutoff to distinguish between higher and lower viral load, with only five patients in the cohort having such a high viral burden. All patients were HBV surface antigen (HBsAg) positive and HBeAg negative. There was no clinical or sonographic evidence of cirrhosis or portal hypertension for patients in the study cohort, and all patients were negative for HIV and HCV antibody.
The control population consisted of 66 healthy donors obtained as buffy coat packs from the Irish Blood Transfusion Service. The demographics of the control subjects were mostly unknown. Therefore, a cohort consisting of 13 African, 14 Caucasian, and 9 Asian healthy uninfected control subjects attending the Genitourinary and Infectious Diseases Clinic, St. James's Hospital, Dublin, was also studied. Ethical approval for the study was obtained from the Research Ethics Committees of St. James Hospital and Adelaide and Meath Hospital incorporating the National Children's Hospital, Dublin, and informed consent was obtained from all donors.
Antibodies and flow cytometry
Peripheral blood mononuclear cells (PBMC) were prepared from blood samples by density gradient centrifugation over Lymphoprep (Nycomed, Oslo, Norway). Freshly isolated PBMC were stained with monoclonal antibodies (mAbs) specific for human CD3, Vδ1, Vδ2, CD45RA, and CD27, obtained from BD Biosciences (Oxford, United Kingdom), Immunotools (Friesoythe, Germany), eBioscience (Hatfield, United Kingdom), and R&D Systems (Abingdon, United Kingdom). Cells were analyzed using a FACSCalibur flow cytometer and CellQuest software (BD Biosciences).
Investigation of IFN-γ-production by Vδ2 T cells in HBV
Freshly isolated PBMC were incubated for 4 h in medium alone or with 10 ng/mL phorbol myristate acetate and 1 μg/mL ionomycin (PMA/I), in the presence of 10 μg/mL brefeldin A (Sigma-Aldrich, Dublin, Ireland). Cells were then stained for surface expression of CD3 and Vδ2 and intracellular expression of IFN-γ. The frequencies of IFN-γ+ Vδ2 T cells were then detected by flow cytometry.
Statistical analyses
Statistical analysis was carried out using Prism GraphPad v5.0. Differences between groups were assessed using the Mann–Whitney U-test or unpaired t-test, where appropriate. p-Values of <0.05 were considered significant. p-Values were corrected for multiple testing (p c) by the Bonferroni method.
Results
Circulating Vδ1 and Vδ2 T cell frequencies are increased in asymptomatic patients with chronic HBV infection
PBMC were prepared from 20 HBV-infected subjects and from the buffy coat packs of 23 control subjects. Surface staining with PE-labeled anti-Vδ2 mAb and PE-Cy5-labeled anti-CD3 mAb was performed to identify Vδ2 T cells in the peripheral blood. The mean frequencies of circulating Vδ2 T cells were 3.6±0.4% in controls and 7.8±0.9% in HBV (p=0.0001; Fig. 1). PBMC from 23 HBV-infected subjects and from the buffy coat packs of 21 control subjects were indirectly surface stained with unconjugated anti-Vδ1 mAb (murine immunoglobulin [IgG]), followed by PE-labeled antimouse IgG, in combination with direct surface staining with PE-Cy5-labeled anti-CD3 mAb. Mean frequencies of circulating Vδ1 T cells were 0.8±0.2% in controls and 1.2±0.2% in HBV patients (p=0.02; Fig. 1). These data show significant expansions in both the Vδ1+ and Vδ2+ proportions of circulating γδ T cells in asymptomatic patients with HBV infection compared to uninfected control subjects.
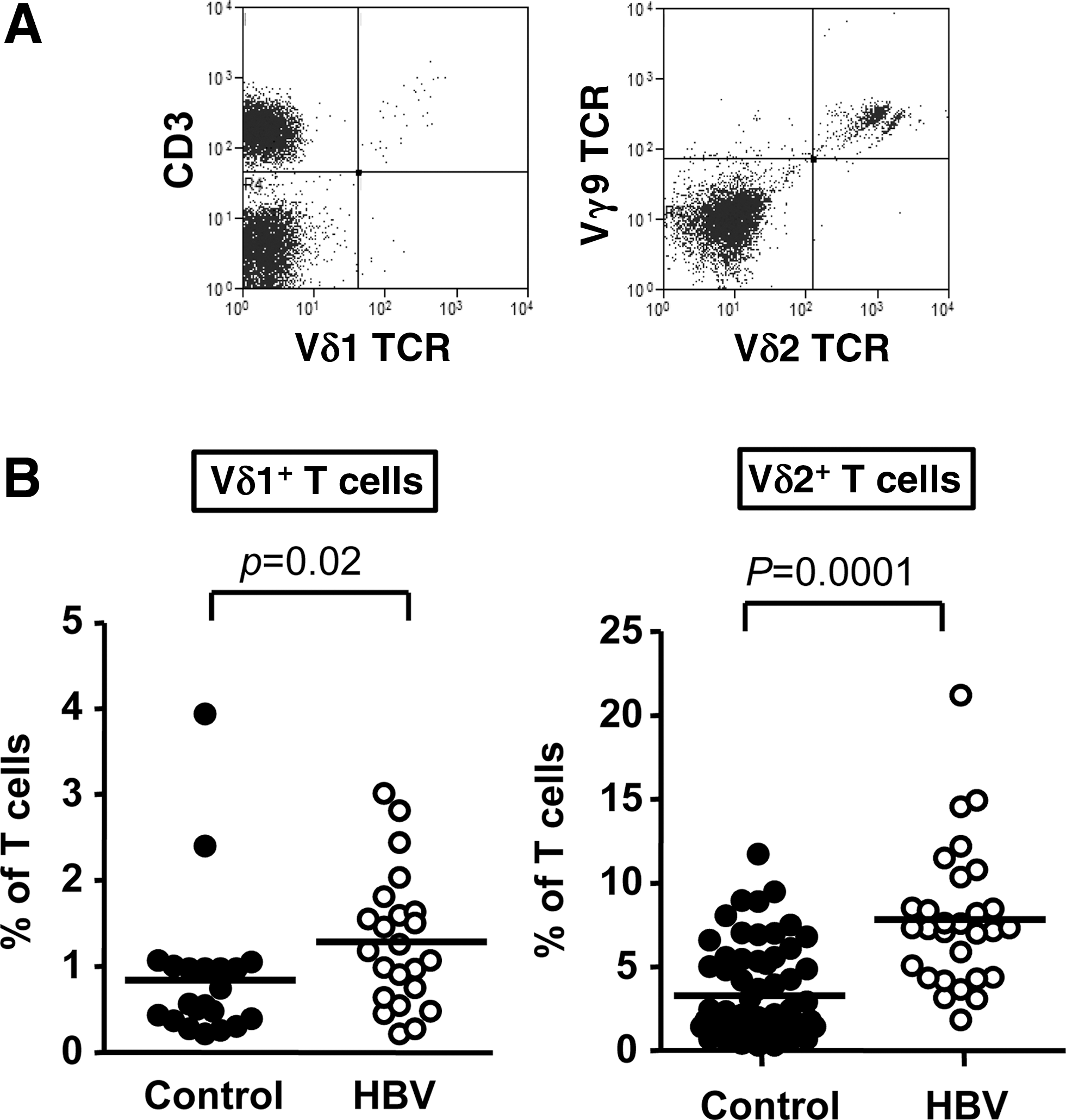
γδ T cell subset frequencies are higher in the peripheral blood of patients with chronic hepatitis B virus (HBV) infection compared to controls. (
Frequencies of circulating Vδ2 and Vδ1 T cells are similar in African, Caucasian, and Asian healthy control subjects
The patient cohort consisted of individuals of African, Caucasian, and Asian origin. Therefore, blood samples were obtained from healthy controls that were demographically matched to the patient cohort in an effort to identify differences in Vδ1 and Vδ2 T cell frequencies that relate to race and not HBV infection. Blood samples were taken from 13 African, 14 Caucasian, and 9 Asian healthy donors. PBMC preparation and surface staining were performed as above, and the frequencies of Vδ1 and Vδ2 T cells were quantified. The mean frequencies of circulating Vδ1 and Vδ2 T cells were similar in African, Caucasian, and Asian control subjects (Fig. 2). Although Vδ1 T cells were found at slightly higher frequencies in Asians, these data suggest that the frequencies of circulating Vδ1 and Vδ2 T cells do not significantly differ between people of the three ethnicities investigated in this study and, therefore, that the higher frequencies of Vδ1+ and Vδ2+ T cells in HBV patients (Fig. 1) are not biased by the demographics of the patient group.
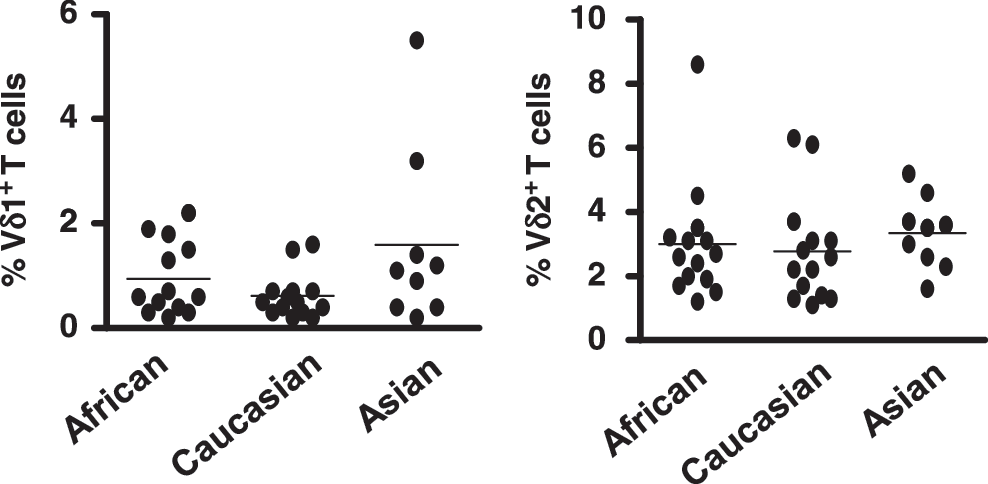
Frequencies of circulating Vδ1 and Vδ2 T cells are similar in African, Caucasian, and Asian healthy control subjects. Frequencies of circulating Vδ1 (left) and Vδ2 T cells (right), as percentages of total T cells, in 13 African, 14 Caucasian, and 9 Asian healthy control subjects. Horizontal bars show means.
The frequencies of circulating IFN-γ-producing Vδ2 T cells are higher in HBV patients than in healthy control subjects
Freshly isolated PBMC from 10 HBV-infected subjects and 18 control subjects were incubated for 4 h in medium alone or with PMA/I, in the presence of brefeldin A. Cells were then stained for surface expression of Vδ2 and CD3 and intracellular expression of IFN-γ to identify IFN-γ-producing Vδ2 T cells. The frequencies of IFN-γ-producing Vδ2 T cells were calculated as percentages of the Vδ2 T cells and as percentages of the total T cells. In the absence of stimulation, the mean frequencies of IFN-γ-producing Vδ2 T cells, as a percentage of total Vδ2 T cells, were 2.7±0.7% in controls and 6.7±1.4% in HBV patients (p=0.01; Fig. 3). After PMA/I stimulation, the mean frequencies of IFN-γ-producing Vδ2 T cells were similar in HBV patients and uninfected controls (p=0.6; Fig. 3). In the absence of stimulation, the mean frequencies of IFN-γ-producing Vδ2 T cells, as a percentage of total T cells, were 0.06±0.02% in controls and 0.4±0.2% in HBV-infected subjects (p=0.005; Fig. 5), and these differences were not observed following PMA/I stimulation (p=0.2; Fig. 5). Thus, in the absence of ex vivo stimulation, Vδ2 T cells from the HBV-infected individuals more likely produce IFN-γ than Vδ2 T cells from uninfected controls.
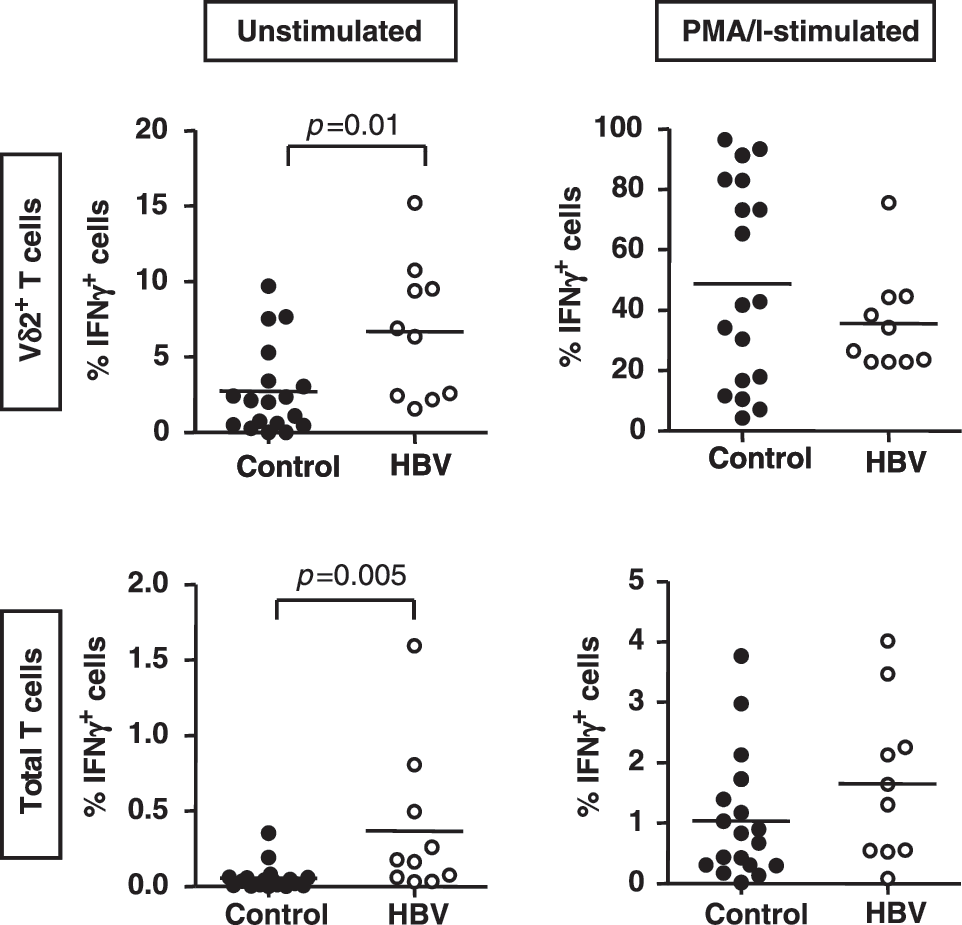
Frequencies of circulating IFN-γ-producing Vδ2 T cells are higher in HBV-infected patients than in control subjects. Scatterplots showing the frequencies of IFN-γ-producing Vδ2 T cells as a percentage of Vδ2 T cells (top) and as a percentage of total T cells (bottom), following incubation in medium alone (left) and following incubation in medium conditioned with PMA/I (right) in 18 uninfected control subjects (filled circles) and 10 HBV-infected patients (unfilled circles). Horizontal bars show means.
Vδ1 and Vδ2 T cells from patients with HBV infection predominantly display memory phenotypes
PBMC from 17 HBV-infected subjects and from the buffy coat packs of 21 control subjects were stained with mAbs specific for Vδ1, CD3, CD45RA, and CD27. The scheme described by Dieli (19) was then used to identify the naïve, central memory (TCM), effector memory (TEM), and terminally differentiated (TEMRA) Vδ1 T cells (Fig. 4A). The frequencies of naïve Vδ1 T cells, as a proportion of the total Vδ1 T cell population, were significantly lower in HBV patients compared with controls (p=0.0002; Fig. 4B), while the frequencies of TCM, TEM, and TEMRA Vδ1 T cells were significantly higher (p=0.02, 0.004, and 0.02; Fig. 4B). These data show that all memory subsets of Vδ1 T cells are expanded in HBV infection while naïve Vδ1 T cells are significantly lower, suggesting that Vδ1 T cells are actively involved in the immune control of HBV.
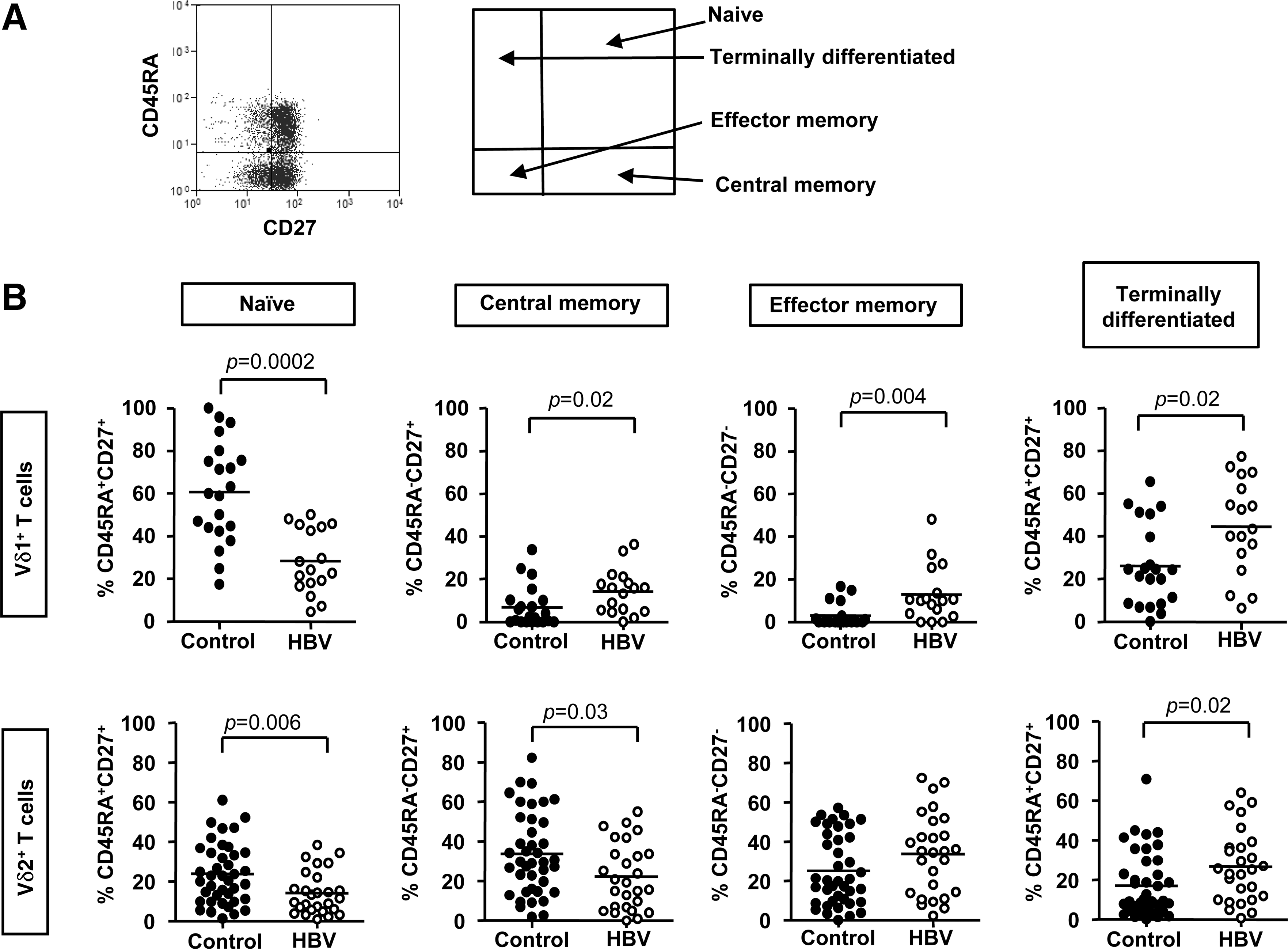
Circulating effector-memory Vδ1 and Vδ2 T cells are expanded in HBV infection while naïve Vδ1 and Vδ2 T cells are reduced. (
Further phenotypic studies were performed to ascertain whether the frequencies of naïve, TCM, TEM, and TEMRA Vδ2 T cells were altered in HBV infection. PBMC from 27 HBV-infected subjects and from 40 control subjects were surface stained for Vδ2, CD3, CD45RA, and CD27 expression. The frequencies of naïve and TCM Vδ2 T cells, as a proportion of the total Vδ2 T cell population, were significantly lower in HBV patients, compared with controls (p=0.006 and 0.03; Fig. 4B), while the frequencies of TEM Vδ2 T cells were similar (p=0.1; Fig. 4B). The frequencies of TEMRA Vδ2 T cells were significantly higher in HBV patients (p=0.02; Fig. 4B). These data suggest that effector Vδ2 T cells are also expanded in order to facilitate the control of HBV infection.
The frequencies of circulating Vδ2 T cells do not correlate with viral load, serum ALT, or age in HBV but are slightly higher in female subjects
Circulating Vδ1 and Vδ2 T cell frequencies in groups of HBV patients within the study cohort were compared based on clinical parameters. There were no significant differences in the frequencies of Vδ1 and Vδ2 T cells between patients with low and high viral loads, low and high ALT, or in patients aged 19–35 years compared to patients aged 35–55 years (Fig. 5). There were significantly higher frequencies of Vδ2 T cells in HBV-infected females compared with HBV-infected males, while the frequencies of Vδ1 T cells were similar (p=0.02; Fig. 5). Spearman correlation tests confirmed that there were no linear correlations between the frequencies of Vδ1 and Vδ2 T cells and viral load, ALT, or age. These data suggest that while the frequencies of circulating Vδ2 T cells are slightly higher in female HBV-infected subjects than in their male counterparts, the frequencies of Vδ1 and Vδ2 T cells do not significantly correlate with the clinical parameters investigated here.
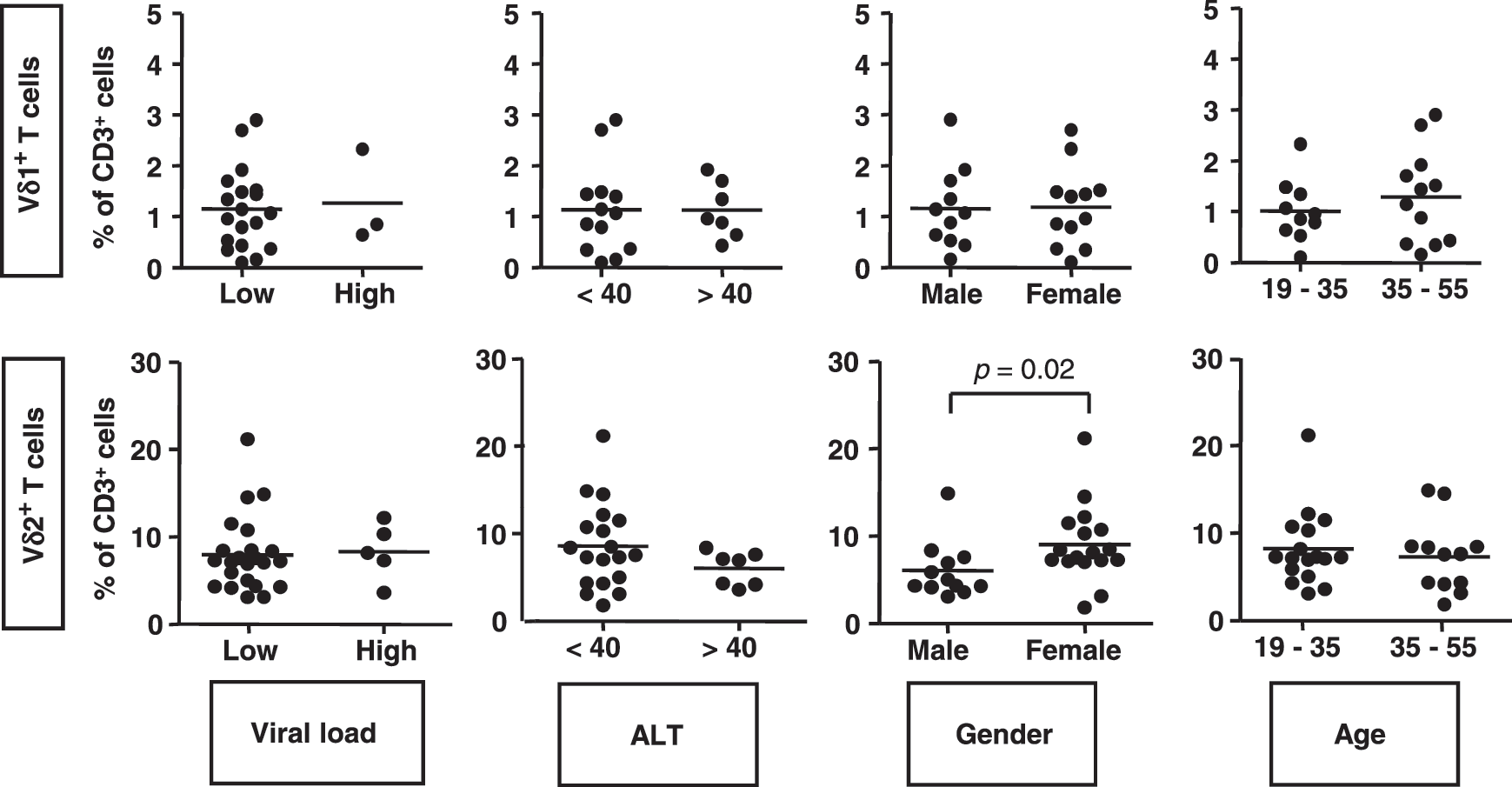
Frequencies of circulating Vδ1 and Vδ2 T cells in HBV patient subsets based on viral load, ALT, sex, and age. Scatterplots showing frequencies of Vδ1 (top) and Vδ2 (bottom) T cells in male and female HBV-infected patients with viral load between both 10 and 100,000 copies/mL (low) or between 100,000 and 5×108 copies/mL (high), with ALT <40 IU/mL (<40) or >40 IU/mL (>40) and aged between 19 and 35 years or 35 and 55 years. Horizontal bars show means. ALT, alanine aminotransferase.
Discussion
Vδ2 T cells are the predominant γδ T cell subset in human blood and are capable of producing IFN-γ and TNF-α (23,24), killing target cells (3,22,23), inducing activation and maturation of neutrophils (15,16), monocytes (24), B-cells (7,9), and dendritic cells (14,23,35), and presenting antigen to conventional CD4+ and CD8+ T cells (8). Their multifunctional capacity makes them ideal candidates for immunotherapy, and they are already the focus of several clinical trials (5,12,20,42,51). Vδ1 T cells are mainly found at mucosal surfaces and can exhibit immunostimulatory (26,29) and immunoregulatory (6) functions and are also under consideration as therapeutic targets (49).
Previous studies have revealed that Vδ2 T cell frequencies are decreased in the peripheral blood of chronic HBV patients with liver disease (11) and with HBV-ACLF (10), whereas Vδ1 T cells are expanded in the blood of HBV-ACLF patients and exhibit enhanced cytotoxicity and cytokine production compared to Vδ1 T cells from healthy controls. A cohort of asymptomatic, HBeAg-negative patients with persistent HBV infection with little evidence of liver disease was studied to provide an insight into the possible roles of γδ T cells in the immune control of HBV infection without significant liver injury. IFN-γ-producing Vδ1 and Vδ2 T cells with effector memory phenotypes are found at higher frequencies in the peripheral blood of these patients compared to controls. This suggests that these γδ T cell subsets are expanded in the circulation of patients, although the possibility that their increased frequencies are the result of a contraction of other T cell subsets cannot be excluded. Future studies are required to determine if Vδ1 and Vδ2 T cells are also expanded in the patients' livers. However, one study (44) has demonstrated that the numbers of peripheral and intrahepatic lymphocyte subtypes correlate closely with each other. Therefore, it is likely that effector memory Vδ1 and Vδ2 T cells may contribute to the immune control of HBV infection without causing liver pathology. The finding in the present study contrasts with the depletions of these cells in patients with HBV-associated liver disease and HBV-ACLF (10,11), which may facilitate liver damage. A role for Vδ2 T cells in immunity against HCV was reported by Agrati et al. (1), while the same group has provided evidence that Vδ1 T cells contribute to liver damage in patients with HCV infection (2).
As well as being expanded, the present study has revealed that greater proportions of Vδ1 and Vδ2 T cells from asymptomatic HBV patients displayed effector memory phenotypes compared to Vδ1 and Vδ2 T cells from uninfected control subjects, while the proportions of naïve Vδ1 and Vδ2 T cells were substantially lower. This suggests that Vδ1 and Vδ2 T cells are actively involved in immunity against HBV. Previous studies have shown that circulating HBV-specific CD8+ T cells from patients with acute HBV infection predominantly express memory T cell phenotypes, suggesting that they are also actively involved in immunity against HBV (52,57). It was also found that in the absence of ex vivo stimulation, the frequencies of IFN-γ-expressing Vδ2 T cells were higher in the HBV-infected patients compared to healthy controls. This suggests that Vδ2 T cells may control HBV infection and prevent immune-mediated damage by a mechanism that involves IFN-γ-mediated viral clearance. A similar role for subsets of NK cells in IFN-γ-mediated control of HBV without liver-damaging cytotoxicity has been proposed (13,48,60). The elevated frequencies of circulating Vδ1 T cells with effector memory phenotypes, although not as significant as those observed for Vδ2 T cells, suggest that these cells also play a role in the antiviral immune response against HBV, possibly via the production of IFN-γ and TNF-α (52). Since Vδ1 T cells have previously been implicated in the pathogenesis of HCV infection and arthritis, they might require strict regulation in asymptomatic HBV infection (2,4). However, they have also previously been shown to regulate inflammatory responses of CD8+ T cells in the small intestine via the suppression of IFN-γ, granzyme-B, and NKG2D expression (6). Further work is required to determine whether this γδ T cell subset plays a predominant antiviral or regulatory role in the control of HBV infection.
In summary, a previous study has provided evidence that failure to clear HBV infection is associated with depletions of Vδ2 T cells and reduced IFN-γ expression, with the greatest depletions observed in patients with the highest viral loads (11). The data in the present study support this hypothesis, with higher frequencies of IFN-γ-producing, effector memory Vδ2 T cells being found in a cohort of asymptomatic HBV patients with low viral loads. These findings implicate Vδ2 T cells as key players in the control of HBV replication, most likely via the production of IFN-γ. Since Vδ2 T cells have already been identified as targets for immunotherapy and they are already the focus of several clinical trials (5,12,20,42,51), it is proposed that they might be used as the basis for future immunotherapies to treat HBV patients with persistent symptomatic infection.
Footnotes
Acknowledgments
This study was supported by grants from the Irish Health Research Board and Science Foundation Ireland.
Author Disclosure Statement
No competing financial interests exist.