
Abstract
The objective of this work was to evaluate the influence of human papillomavirus (HPV) vaccination on peripheral blood mononuclear cell (PBMC) proliferation and cytokine gene transcription. PBMCs isolated after HPV immunization were incubated with HPV vaccine, phytohemagglutinin, or buffer. Cell proliferation was assessed by MTT reduction assay. RNA was extracted from PBMCs, and the relative concentration of cytokine messenger RNA (mRNA) transcripts (IFN-β, IFN-γ, IL-12, TNF-α, IL-6, IL-17, or IL-10) relative to transcription of the β-actin gene was determined by real-time polymerase chain reaction. PBMC proliferation in response to HPV vaccine and PHA were greater than that observed in unstimulated cells (p<0.001). Cytokine mRNAs were upregulated in stimulated PBMC cultures. The median increase in vaccine-stimulated cultures was: IFN-β=334.4-fold; IL-12=46.33-fold; IFN-γ=12.64-fold; IL-6=9.07-fold; IL-17=7.33-fold; IL-10=6.47-fold; and TNF-α=2.36-fold. The IFN-β expression was significantly higher (p<0.05). Proliferative PBMC responses and multiple cytokine gene expression were detected in women who received the HPV vaccine.
Introduction
H
The induction of a pro-inflammatory immune response is necessary to break the tolerance induced by HPV (16). The successful initiation of pro-inflammatory signaling is a component of new treatments to promote an adequate immune response and disrupt the anti-inflammatory barrier triggered by HPV infection.
Therapeutic interventions with vaccines to induce an effective immune response have the potential to treat latent infection as well as clinically apparent lesions.
Two HPV L1 VLP vaccines have been developed: Cervarix®, a bivalent HPV 16, 18VLP vaccine from GlaxoSmithKline, and Gardasil®, a quadrivalent HPV 16/18/6/11 vaccine from Merck Vaccines. These products are to be delivered in a three-shot immunization schedule. They induce, at their peak after the third immunization at 6 months, high concentrations of neutralizing antibodies to L1, and virtually all subjects in the vaccine trials have seroconverted (17).
Strategies exist to enhance the therapeutic immune capability of HPV vaccination. Adjuvants enhance immunity to vaccines by a variety of mechanisms. Adjuvants have been traditionally used to increase the magnitude of an adaptive response to a vaccine, based on antibody titer and avidity or ability to prevent infection. A second role for adjuvants has become increasingly important: guiding the type of adaptive response to produce the most effective forms of immunity for each specific pathogen (9,15). Monophosphoryl lipid A (MPL)-based adjuvants (AS04) activate monocyte and myeloid dendritic cells. Such adjuvants induce CD4+ Th1 and low-level CD8+ T-lymphocyte responses (1).
The aim of this study was to evaluate the cell-mediated immune response to HPV after the bivalent HPV16/18 AS04-adjuvanted vaccination, assessing cell proliferation and quantifying cytokine mRNA expression.
Materials and Methods
Study population
To be eligible for the extension study, 30 subjects had to have received three doses of the HPV-16/18 AS04-adjuvanted vaccine (GlaxoSmithKline) intramuscularly into the deltoid at 0, 1, and 6 months. Blood specimens were collected for immunologic assays after 1 month following the last boost vaccination (month 7).
A similar assay was conducted using peripheral blood mononuclear cells (PBMCs) isolated from subjects who has not received the HPV-16/18 vaccine. This control group consisted of samples from the same individuals prior to the initial vaccination (month 0) to exclude the possibility of the vaccine adjuvant being responsible for activating the cell proliferation.
The proliferation from PBMCs was assessed in different experiments: (a) PBMCs from vaccinated subjects incubated in medium alone or medium plus vaccine or medium plus PHA, and (b) PBMCs from unvaccinated subjects incubated in medium plus vaccine (control).
Cell culture and MTT cell proliferation assay
PBMCs from 30 patients before and 1 month after the final HPV immunization were isolated by density gradient separation (Histopaque® 1077, Sigma-Aldrich). The cells were washed twice with 5 mL of 0.9% NaCl and resuspended in 4 mL of RPMI 1640 medium supplemented with L-glutamine, sodium bicarbonate, 1% antibiotic-antimycotic, and 10% heat-inactivated autologous serum (drawn before vaccination). Cells were counted with a hemocytometer, and 105 PBMC/well were incubated in sterile round-bottom plates for 72 h with medium, 5 μg/mL antigen (HPV L1 virus-like particle [VLP] HPV16/18 vaccine adjuvant AS04), or 2% v/v phytohemagglutinin M (PHA; Gibco®) in duplicate. The cells were incubated at 37°C in 5% CO2.
Cell proliferation was assessed by the MTT assay. For this procedure, 10 μL MTT (Sigma Aldrich; 5 mg/mL) was added to microtiter plate wells, and the plate was incubated for 4 h. The formazan crystals produced were dissolved by resuspension in 100 μL lysis buffer (10% SDS in 0.001 N HCl), and the optical density in the wells was obtained by a microplate spectrophotometer at 540 nm.
Relative quantification of cytokine mRNAs
For analysis of cytokine mRNA expression, cellular RNA was extracted from PBMCs using the RNA Isolation Mini RNAspin kit (GE Healthcare).
Relative quantification of cytokine mRNAs (IFN-β, IFN-γ, IL-12, TNF-α, IL-6, IL-17, or IL-10) (2,5,12,20) in comparison to transcripts of the β-actin gene (endogenous control) (19) was performed in real-time polymerase chain reaction (PCR) using a SuperScript® SYBR® Green III Platinum One-Step qRT-PCR kit (Invitrogen).
The reactions were performed in a final volume of 25 μL containing 10 ng RNA, 12.5 μL of Master Mix, forward and reverse primers (10 pmol) of each cytokine gene, and autoclaved Milli-Q water, under the following conditions: 55°C for 30 min, 95°C for 10 min, followed by 40 cycles at 95°C for 15 sec and at 60°C for 1 min.
The equipment used for amplification, detection, and quantification of samples was the 7500 Fast Real-Time PCR System (Applied Biosystems). The threshold cycle (CT) values obtained were used for the comparative analysis of mRNA expression of cytokines.
Statistical analysis
Data were entered into the GraphPad Prism software v6.0 (GraphPad Software Inc.), and submitted to one-way analysis of variance followed by Bonferroni's Multiple Comparison Test.
Results
The proliferation from PBMCs was assessed in PBMCs from vaccinated subjects incubated in medium alone or medium plus vaccine or medium plus PHA and PBMCs from unvaccinated subjects incubated in medium plus vaccine (control). The concentration of viable PBMCs from vaccinated subjects incubated with medium plus vaccine was higher (18.13%) than those incubated in medium alone and PBMCs from control subjects. This difference was highly significant (p<0.001; 95% CI 12.45–23.80).
Otherwise, the viable PBMCs from vaccinated subjects incubated with medium plus vaccine (18.13%) and incubated with medium plus PHA (27.3%; 95% CI 21.23–33.36) were similar. Results are shown in Figure 1.
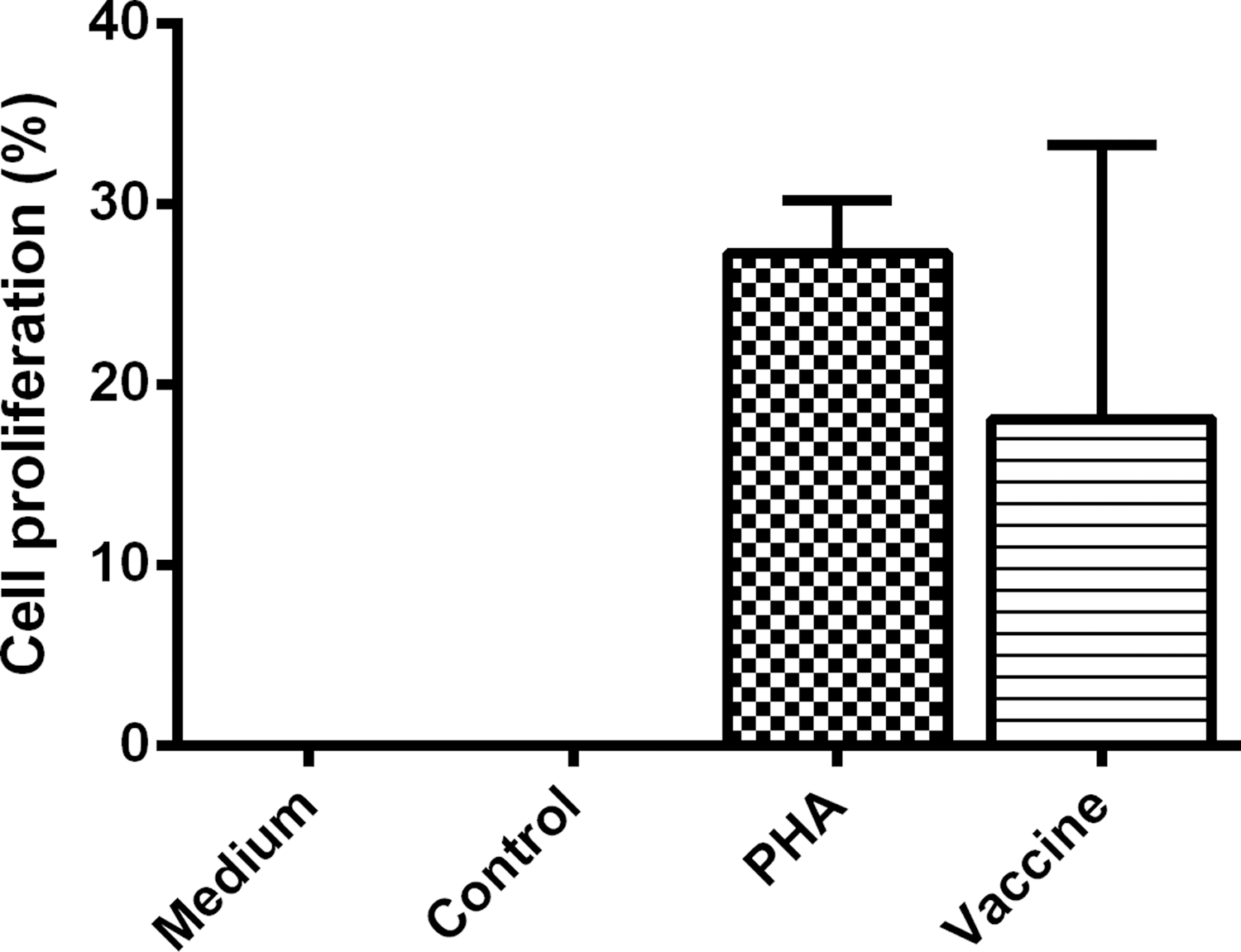
The proliferation from peripheral blood mononuclear cells (PBMCs) was assessed in PBMCs from vaccinated subjects incubated in medium alone or medium plus vaccine or medium plus PHA, and PBMCs from unvaccinated subjects incubated in medium plus vaccine (control).
Seven different cytokine mRNAs were evaluated in vaccine HPV-treated PBMCs, obtained following vaccination of recipients. All cytokine mRNAs were upregulated in HPV vaccine-stimulated PBMC cultures compared to levels in unstimulated cultures. The median increase in the cytokine mRNAs were: IFN-β, 334.4-fold (95% CI 37.49–706.3); IL-12, 46.33-fold (95% CI 4.29–96.96); IFN-γ, 12.64-fold (95% CI 2.44–27.72); IL-6, 9.07-fold (95% CI 1.64–16.5); IL-17, 7.33-fold (95% CI 0.73–15.40); IL-10, 6.47-fold (95% CI 0.84–12.09); TNF-α, 2.36-fold (95% CI 0.69–4.03; Fig. 2). In the PBMCs from women who were stimulated with the HPV vaccine, IFN-β mRNA expression was significantly higher than mRNAs for IFN-γ, TNF-α, IL-6, IL-10, and IL-17 (p<0.01). In the control group, no upregulation of any cytokine mRNAs was observed in stimulated PBMC cultures.
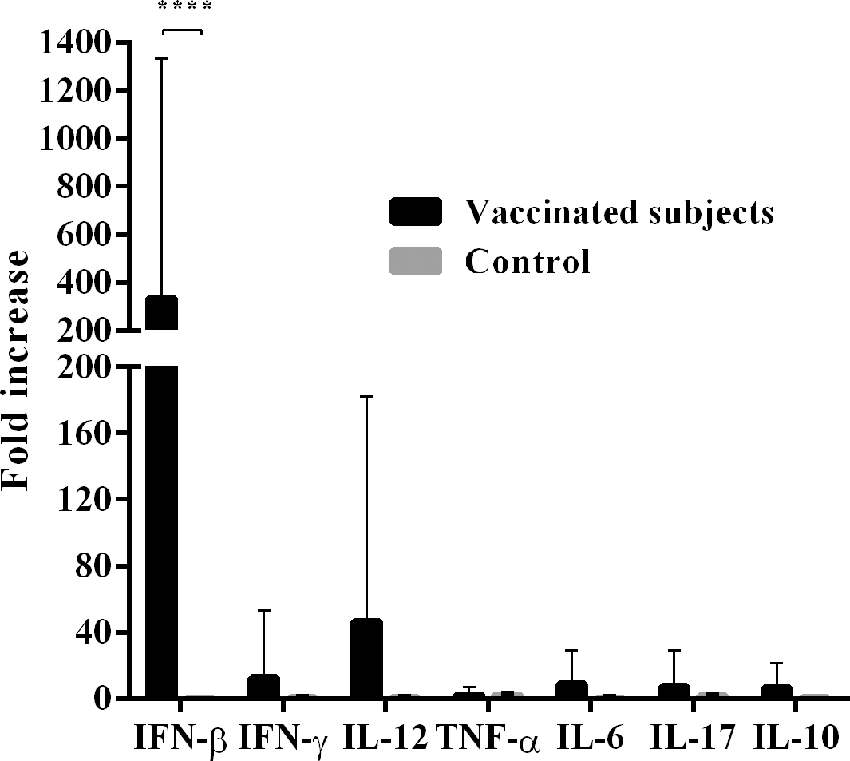
Cytokine mRNA expression in PBMC from HPV16/18 AS04-adjuvanted vaccinated women. The average increase of cytokine expression is shown on the vertical bar. For all analyses, p-values of <0.01 were considered statistically significant. HPV, human papillomavirus.
Discussion
In the present study, a proliferative PBMC response to the HPV-16 and HPV-18 vaccine was observed in immunized women. Previous studies found that a significant cellular mediated immune (CMI) response is induced following HPV vaccination (4,7,13,14).
This study compared samples from the same patient, stimulated by HPV vaccine or incubated in culture medium alone, and significant differences were observed in PBMC proliferation. The results reinforce previous studies (4,7,13,14) that show an increase in immune cell activity following vaccination. There is evidence that CMI responses play an important role in the control of HPV infections. A dysfunctional CMI response is likely to underlie the amplified risk that leads to HPV persistence and cervical cancer (3,8). Thus, these findings suggest that the increased PBMC proliferation represents a potential therapeutic effect of HPV vaccination.
The HPV16/18 AS04-adjuvanted vaccine contains the VLP antigens of HPV type 16 and 18 L1 proteins and uses the adjuvant AS04, which is a combination of aluminum hydroxide and MPL. The MPL formulated on alum (AS04) stimulates a polarized Th1 cell response (3). Therefore, it is very likely that the vaccine adjuvant may also have contributed to the cellular immune response observed in vaccinated women.
In addition, it was observed that PBMC-derived cytokine mRNA expression also increased in HPV vaccine-stimulated cultures from vaccinated women. Among the studied cytokines, IFN-β mRNA expression was significantly higher than mRNAs were for IFN-γ, TNF-α, IL-6, IL-10, and IL-17 (p<0.05). This suggests that immunization with the HPV vaccine may preferably stimulate an antiviral immune response, as evidenced by robust induction of IFN-β.
A possible association between deficits in production of cytokines and persistent HPV infections has been postulated too (18). Thus, various cytokines, including IFN-β, may play roles in checking the growth of HPV-infected cells, and viral persistence, disease progression, and/or malignant transformation could involve escape from these mechanisms.
In this context, it is possible that a post-vaccination activation of the CMI response is of crucial importance for achieving a therapeutic effect following HPV vaccination. Furthermore, potentially therapeutic HPV vaccines need to induce both arms of the CMI response, since T-helper cells (CD4+) help to augment the cytotoxic T-cell (CD8+) response (10,11).
An effort to enhance the CMI response must be addressed in future developments of therapeutic vaccines. Adjuvants such as AS04 have been used as a complementary tool to enhance antivaccine immunity. However, it is important to continue the development of other adjuvants that, working together with specific antigens, could lead to more effective HPV therapeutic vaccines, decreasing the morbidity and mortality of HPV-associated malignancies.
Footnotes
Acknowledgments
This investigation was funded by FAPESP (Fundação de Amparo a Pesquisa do Estado de São Paulo) under award number R56AI097146-01A1.
Author Disclosure Statement
No competing financial interests exist.