
Abstract
Porcine circovirus type 2 (PCV2) is associated with a number of diseases and syndromes, collectively referred to as porcine circovirus-associated disease. The main objective of this study was to define some in vitro correlates of protection after injection of inactivated PCV2 vaccines with a defined antigen mass. Twelve pigs were vaccinated with three different doses of inactivated, whole-virus antigen (211–844 ng), while four animals were injected with a commercial vaccine (positive control) and four other pigs were mock-vaccinated with phosphate-buffered saline (PBS) in the same oil emulsion. Four weeks later, they were intranasally challenged with 2 × 105 TCID50 of a PCV2a strain. Antibody was measured in blood and oral fluids by enzyme-linked immunosorbent assay (ELISA) and a neutralization assay. PCV2 was quantified in serum by real-time polymerase chain reaction for ORF2 gene. PCV2-specific cell-mediated responses were investigated by an IFN-γ release assay in whole blood, IFN-γ ELISPOT, and lymphocyte proliferation (Ki-67 and BrDU assays). All the vaccines under study but mock provided complete or incomplete protection from PCV2 infection in terms of post-challenge viremia. Serum antibody titers (ELISA and neutralizing) after vaccination were not correlated with protection, as opposed to the early neutralizing antibody levels of vaccinated pigs at day 7 after infection. Cell-mediated immune parameters showed a good correlation with vaccine efficacy. In particular, the IFN-γ release assay at 3 weeks after vaccination was an effective marker for predicting protection. All control pigs always tested negative in assays of cell-mediated immunity. Our results outline in vitro testing procedures toward reduced animal usage in the control of PCV2 vaccine batches.
Introduction
P
Methods
Cells and virus
PCV2 (Bio Bank Veterinary Resources, IZSLER, code VIR RE RSCIC 151) was propagated in circovirus-free PK15c28 cells (porcine kidney cells, IZSLER cell bank code BS CL 179). Cells were cultured in Minimum Essential Medium (MEM) supplemented with 5% fetal bovine serum (FBS), antibiotics (100 IU/mL penicillin and 100 μg/mL streptomycin final) and 2 mM L-glutamine until confluence. PK15c28 cells were trypsinized and infected with PCV2 at a multiplicity of infection of 0.1 for 30 min at 37°C in 5% CO2. Cells were then transferred into cell culture flasks for 4 days until confluence. PCV2-infected cells were lysed by two freeze–thaw cycles at −20°C and clarified by centrifugation in a JA-20 rotor (Beckman Coulter) at 7740 g for 15 min at 4°C. Fifty percent virus infectivity titers were determined by direct immunofluorescence in PK15c28 cells grown in 96-well microtiter plates. Briefly, 1.5 × 104 cells/well (50 μL) were inoculated with 10-fold virus dilutions (50 μL/well) and grown at 37°C in 5% CO2 for 24 h. Then, 100 μL/well of complete medium supplemented with 1000 IU/mL of swine recombinant IFN-γ (AbD Serotec) was added to semi-confluent monolayers. After 3 days of culture, the medium was discarded, monolayers were washed once with PBS, fixed with 80% acetone, stained with an anti-PCV2 polyclonal antiserum conjugated to fluorescein isothiocyanate (VMRD, Pullman, cat CJ-F-PCV2), and examined for fluorescent foci by ultraviolet (UV) microscopy. A cryolisate of PK15c28 cells (mock virus) was set up by freezing (−80°C) and thawing twice a cell suspension in the same medium employed for PCV2 virus propagation. After clarification, mock virus was frozen in aliquots at −80°C and used in the assays reported hereunder at a dilution corresponding to the cell concentration of the PCV2 stocks.
Virus inactivation, concentration, and pelleting
Clarified (12520 g, 10 min) PCV2 (titer: 105 TCID50/mL) was treated with beta-propiolactone (Sigma-Aldrich, cat P5648) twice in 3 days (0.05% final, 4°C). Then, the inactivated viral suspension was concentrated 20× through a 10,000 MWCO Hydrosart Vivaflow 200 tangential flow module (Sartorius Stedim Biotech GmbH) and pelleted in a JA-20 rotor (Beckman Coulter) at 48400 g for 16 h at 4°C. The pellet was resuspended overnight at 4°C in sterile PBS to obtain further 40-, 20-, and 10-fold concentrations, respectively. The recovery of PCV2 was confirmed by electron microscopy, real-time polymerase chain reaction (PCR) for the ORF2 gene, and ELISA. PCV2 virion concentrations were calculated by sucrose gradient analysis and UV spectroscopy, as described elsewhere (37). The concentration of PCV2 amounted to 1,954, 977, and 489 ng/mL for 40-, 20-, and 10-fold concentrated antigen, respectively.
Vaccine formulation
The above-mentioned inactivated PCV2 suspensions were mixed with Circovac adjuvant (Merial) at an antigen/adjuvant ratio of 1:2.31. A mock vaccine was also prepared by using sterile PBS instead of inactivated antigen at the same ratio. PCV2 and mock vaccines were kept at 4°C until use.
Experimental design
All animal experiments were conducted at IZSLER, Brescia, in compliance with the institution's Ethical Committee for Animal Experimentation. The treatment, housing, and husbandry conditions conformed to the European Union Guidelines. Animal care and procedures were in accordance with the guidelines of the Good Laboratory Practices (GLP) under the supervision of the Ethical Committee for Animal Experimentation and the Italian National Veterinary Service.
A pilot study was carried out first on four, 30-day old, Goland hybrid piglets to determine the possible influence of residual maternally derived antibody (MDA) titers on PCV2 infection. Then, the vaccine study was carried out on 20 Goland hybrid piglets (three litters) of a herd located in Brescia Province, Italy. Weaned, 30-day-old piglets were transferred to IZSLER isolation units and clinically inspected. A first blood sample was collected from each animal on the arrival day to measure MDA titers to PCV2. After 3 days, pigs were randomly allocated to five groups of four pigs each (all of them including at least one pig from each litter). As a result, the mean MDA titers were not significantly different between groups. Nine days after the arrival, groups 1–3 were immunized intramuscularly with 0.5 mL of three different doses of the experimental vaccine (see Table 1), while group 4 was treated with 0.5 mL of Circovac (lot L392813, Merial S.A.S., unknown whole virus mass concentration) according to the manufacturer's protocol. Mock vaccine (0.5 mL) was administered to a control group (group 5) housed separately from the others. At 66 days of age (27 days after PCV2 and mock vaccination, 0 days post challenge [dpc]), all pigs were challenged intranasally with 4 mL of a suspension containing 105.3 TCID50 of the above-mentioned, homologous PCV2 strain. Blood was taken in heparinized vacuum tubes or tubes without anticoagulant both before (days −27 and −7) and after challenge (dpc 7, 14, 21, 24, 29, and 36). Oral fluids from vaccinated and control groups were collected at the same times by a cotton rope.
Phosphate-buffered saline in Circovac adjuvant at the same ratio as in Circovac.
Total and neutralizing anti-PCV2 antibodies
Total PCV2-specific antibody was measured in serum by a competitive ELISA based on monoclonal antibodies to PCV2 ORF2, as previously described (3,32). Titers were expressed as the log2 of the highest antibody dilution inducing 50% inhibition of the optical density (OD) value corresponding to unreacted PCV2 antigen. Neutralizing antibodies in sera and oral fluids were measured by immunofluorescent staining as described by Fort et al. (15), with minor modifications. Briefly, 25 μL of heat-inactivated serum (56°C, 30 min) were serially twofold diluted in 96-well, round-bottomed serology plates from 1:2 to 1:32 in MEM supplemented with 5% FCS. Twenty-five microliters of PCV2 titering 100 focus-forming units were added to each well, and plates were incubated at 37°C, 5% CO2 for 1 h. Then, 1.5 × 104 PK15c28 cells/well were added in a volume of 50 μL. After 30 min incubation at 37°C in 5% CO2, the mixtures of cells, serum, and virus were transferred to a tissue culture 96-well plate and incubated for 24 h. Then, 100 μL/well of complete medium supplemented with 1000 IU/mL of swine recombinant IFN-γ (AbD Serotec, Bio-Rad) was added to semi-confluent monolayers. After 3 more days of culture, cells were washed, fixed, and stained as indicated above for PCV2 titration, and examined for fluorescence by UV microscopy. Cells with nuclear and/or cytoplasmic staining were considered positive, and fluorescent foci were counted in each well. The NA titer was calculated as the reciprocal of the highest serum dilution reducing the number of PCV2 fluorescent foci by 90% at least. Titers <2 were scored as 1 for data analyses. Oral fluids were considered Ab-positive if they significantly reduced (p < 0.05) the number of fluorescent foci, according to the statistical method described by Lorenz (26).
PCV2 DNA in serum
PCV2 DNA quantification was performed as previously described (30) on serum samples by real-time quantitative PCR performed after DNA extraction (DNeasy Blood and Tissue Kit, Qiagen). Results were expressed as PCV2 genome copies/mL of serum.
PCV2-specific IFN-γ release assay
Heparinized blood was distributed in triplicate in 1 mL aliquots in 24-well microtiter plates and incubated with 100 μL of PBS, beta-propiolactone-inactivated PCV2, and mock virus (cryolysate of PK15c28 cells), respectively. The amount of PCV2 inactivated antigen to be used in CMI assays was determined in preliminary Ag dose–response experiments on the basis of the proliferative response of peripheral blood mononuclear cells (PBMC) from PCV2-vaccinated animals (BrdU assay, see below). Plates were incubated for 18 h at 37°C in a humidified, 5% CO2 incubator. Then, plasma was carefully collected and tested for IFN-γ according to the ELISA procedure described in a previous study (10). Briefly, test results were evaluated on the basis of the IFN-γ response to PCV2 and mock antigen (a cryolysate of PK15c28 cells), respectively, and of the spontaneous IFN-γ release in vitro (PBS-stimulated plasma aliquot). Samples were considered as high positive if the OD corresponding to PCV2-stimulated whole blood cultures was higher than that of mock-stimulated and unstimulated cultures by 50 mOD at least, low positive in the 20–49 mOD range, and negative in the 1–19 mOD range.
PCV2-specific IFN-γ-SC
The numbers of PCV2-specific, IFN-γ-SC were determined on PBMC obtained at days 14, 21, and 36 after challenge by an ELISPOT assay. Briefly, 5 × 105 PBMC in 50 μL of RPMI 1640 medium supplemented with 10% heat-inactivated FBS were seeded into Multiscreen Filter plates (Millipore), precoated overnight (4°C) with an anti-porcine IFN-γ monoclonal antibody (5 μg/mL) (P2F6 clone, ThermoScientific). PBMC were incubated for 36 h at 37°C in a 5% CO2, with 50 μL/well of the same inactivated PCV2 and mock antigen, respectively, used in the IFN-γ release assay (see above), and also with Concanavalin-A (Sigma-Aldrich; 10 μg/μL final) as positive control and sterile PBS as negative control, respectively. Then, the ELISPOT assay was performed according to the procedure described in a previous study (10). Samples were scored positive when the number of IFN-γ SC in PBMC cultures incubated with inactivated PCV2 antigen was greater than that of cell cryolysate (mock) and PBS-treated PBMC cultures, representing the nonspecific and spontaneous IFN-γ response, respectively.
PCV2-specific proliferation
PCV2-specific expression of proliferation-related Ki-67 antigen using FITC Mouse Anti-Human Ki-67 Set (Becton Dickinson) was assessed at days −27, −7, 21, and 36 with respect to challenge. PBMC (4 × 106) in RPMI 1640 medium supplemented with 10% FBS were seeded in duplicate in 12-well plates in a volume of 1.5 mL. The wells were supplemented with 0.5 mL of inactivated PCV2 antigen or mock virus, and cells were incubated at 37°C, 5% CO2 for 6 days. Then, nonadherent PBMC were collected, pelleted, and fixed/permeabilized by vortexing and adding, drop by drop, 5 mL of cold, 75% ethanol to the cell pellet. After 2 h at −20°C, fixed/permeabilized PBMC were stained with the FITC Mouse Anti-Human Ki-67 Set and FITC control isotype according to the manufacturer's instructions. Stained cells were analyzed using a Guava EasyCyte HT flow cytometer (Millipore). Lymphocytes were gated on the basis of their forward and side scatter profile, and readings were stopped after acquisition of 10,000 events. Cells displaying fluorescence intensities above the upper limit of the negative control isotype distribution were considered positive.
At day −7 with respect to challenge, proliferation was also assessed using a colorimetric BrdU Cell Proliferation Kit (Millipore). One hundred microliters of PBMC (2 × 106/mL) in RPMI medium supplemented with 10% FBS were incubated in triplicate with 100 μL of inactivated PCV2 antigen, mock virus (both diluted 1:4 in complete medium), and sterile PBS in a 96-well plate at 37°C, 5% CO2 over 6 days. Cells were fixed and analyzed by ELISA for incorporated BrDU as indicated in the manufacturer's protocol.
Statistical analysis
A Kolmogorov–Smirnov normality test was conducted to check Gaussian distributions in the data sets. Then, the differences among the Ab titers at different time points were checked by one-way analysis of variance (ANOVA) for repeated measures in the data sets showing a normal distribution of the variable. Differences between groups before and after treatment (vaccination and viral challenge) were verified by two-way ANOVA for repeated measures (time × treatment). Pearson's correlation coefficient was used to assess potential relationships between the following parameters: viremia, NA and Ab titers, and PCV2-specific IFN-γ-SCs. The correlation between occurrence of viremia and results of the IFN-γ release assay was checked by Fisher's exact test. The significance threshold was set at p < 0.05 (Prism 5.01, GraphPad Software).
Results
Clinical evaluation
In both pilot and vaccination/challenge experiments, none of the pigs—vaccinated or nonvaccinated—developed clinical symptoms such as fever or weight loss that could be referred to PCVAD.
Pilot study
A preliminary experimental infection, according to the aforementioned challenge conditions, was carried out on piglets (n = 4) from the same herd selected for the provision of test animals. This experimental infection aimed to define a proper timing for the challenge infection in pigs born of PCV2-vaccinated sows. On the basis of the findings, piglets were infected at 60 days of age because of a substantial decay of MDA (NA titers <1:4), and the absence of active seroconversion to field PCV2 (data not shown). PCV2 viremia was detected in most pigs from dpc 16 onwards (mean number of genome copies: 80,923/mL). Then, viremia levels gradually decreased, PCV2 DNA being still detectable at dpc 40 in three subjects out of four (data not shown). Therefore, pigs of the above age and immune profile were shown to be fully susceptible to PCV2 infection despite the residual levels of MDA.
PCV2 viremia
All animals were PCV2-negative on the day of challenge. PCV2 DNA analyses confirmed the onset of viremia from dpc 14 or 21 onwards in all pigs belonging to the control group (group 5). On the contrary, all vaccinated animals were protected against PCV2 infection. Only a few of them showed low-titered, transient viremia (see Fig. 1).
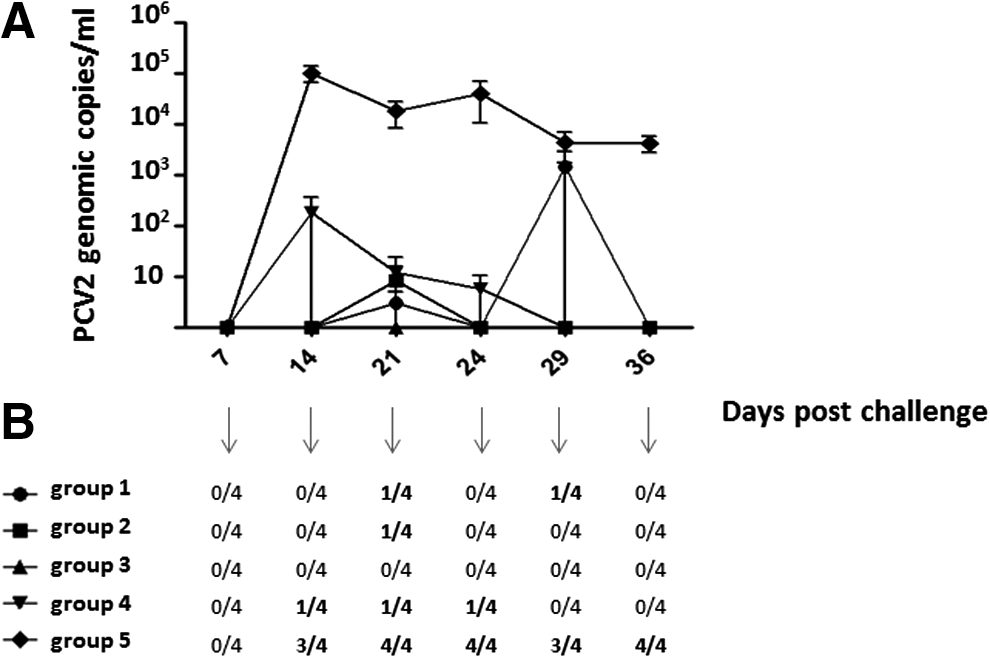
Porcine circovirus type 2 (PCV2) viremia. (
Antibody response to PCV2
At the beginning of the study, moderate MDA levels were still present (mean ELISA log2 titer at day −27 with respect to challenge: 9.60). As in the pilot study, MDA did not interfere with PCV2 infection in control, unvaccinated animals in terms of post-challenge viremia (see Fig. 1). The Ab responses were dissected with respect to both time-course and previous treatment (vaccination vs. nonvaccination). NA responses are summarized in Figure 2. There was no significant difference between groups 3 weeks after vaccination (day 7 before challenge), whereas at dpc 7 a significant booster effect (p < 0.05) on NA titers was observed in vaccinated animals, as opposed to the nonvaccinated ones. Then, at dpc 14, NA in control animals reached the same levels as in vaccinated pigs and declined thereafter. Yet, the peak of NA titers at dpc 14 coincided with viremia onset in control animals. Likewise, no correlation was found in any group between NA titers and PCV2 levels in blood.
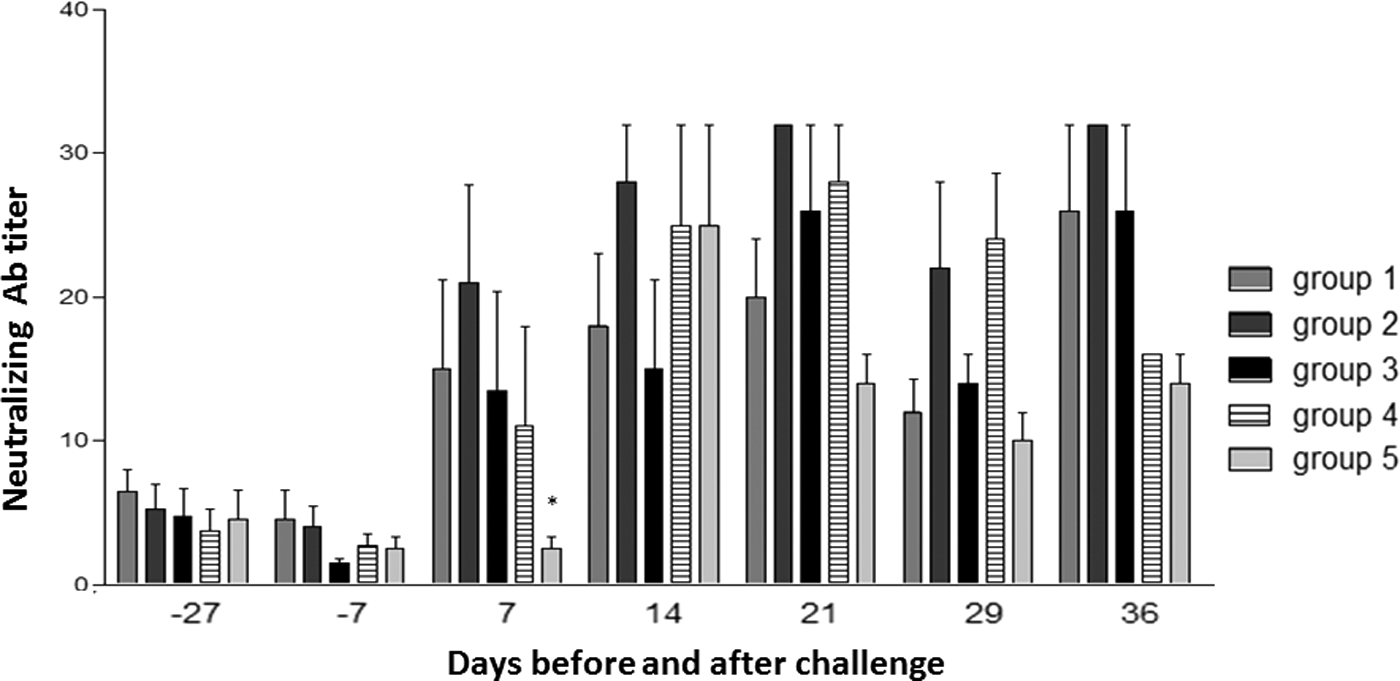
Mean titers of PCV2-specific neutralizing antibodies in sera. Results are shown as mean Ab titers ±1 standard deviation. The asterisk indicates a significant difference between vaccinated and nonvaccinated groups (p < 0.05).
After infection, ELISA antibodies had a different time-course (Fig. 3). As opposed to NA, there was a reduction of the serum Ab titers at dpc 7 (not significant) in all groups under study. From dpc 14 onwards, ELISA Ab levels increased in all groups and remained high until dpc 36 with slight fluctuations. No correlation was found between ELISA Ab titers and viremia levels in PCV2-infected pigs. Concerning NA titers in oral fluids, a slight Ab response was observed only at day −7 before challenge (NA titers <1:2 vs. 1:2 in nonvaccinated and vaccinated pigs, respectively; data not shown).
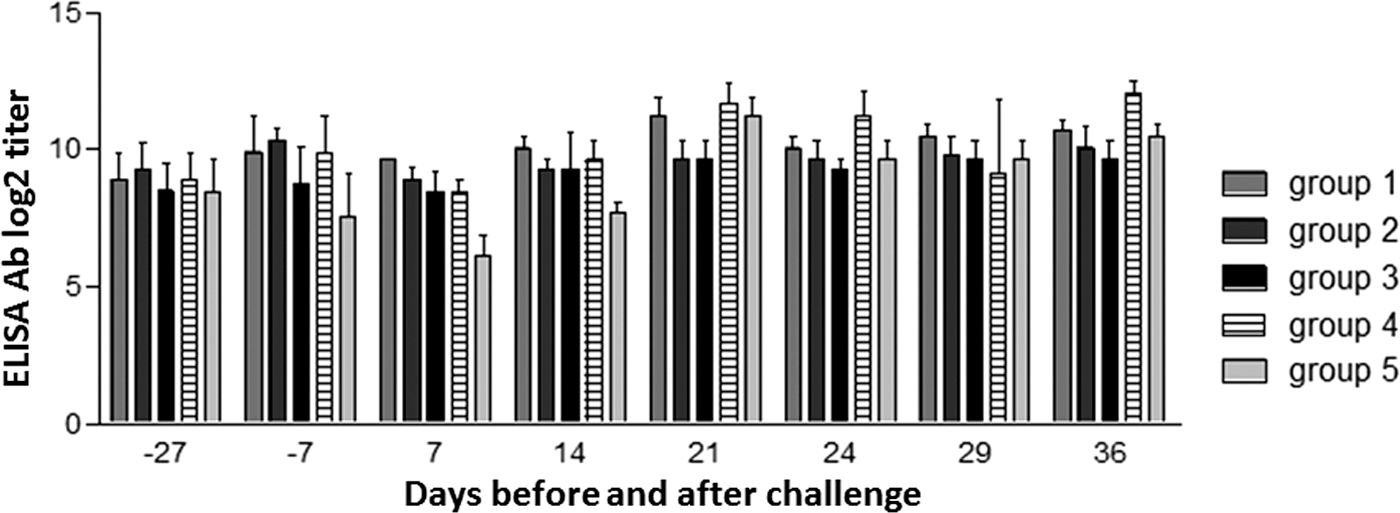
Log2-transformed group means of PCV2-specific enzyme-linked immunosorbent assay antibody titers in sera. Results are shown as mean ±1 standard deviation of the Ab titers.
PCV2-specific IFN-γ release
PCV2-specific IFN-γ release was analyzed in all the animals at days −27, −7, 14, 21, 29, and 36 with respect to challenge infection. Results are shown in Table 2. All pigs scored negative at the beginning of the experiment before vaccination. At day −7 with respect to challenge, the response of vaccinated subjects was clearly different from that of the control ones and was predictive of protection against viremia (p < 0.05 at dpc 14). In particular, group 3 showed higher and more prolonged positive responses than the other vaccinated groups did. Most important, the PCV2-specific IFN-γ release assay on whole blood was able to discriminate between different vaccinated groups. Pigs from group 5 (control group) remained negative at all time points, before and after infection.
ELISPOT positive samples are marked +.
Bold indicates PCV2 viremic serum samples.
PCV2, porcine circovirus type 2; Neg, IFN-γ negative; Low Pos, IFN-γ low positive; High pos, IFN-γ high positive; Tot, total number of test-positive subjects in each group at the indicated time points.
PCV2-specific IFN-γ-SC
IFN-γ-SC in PBMC were investigated by an ELISPOT assay in samples collected at dpc 14, 21, and 36. ELISPOT-positive samples are reported in Table 2. Although there was no clear correlation with the IFN-γ release assay in terms of response intensity, once again nonvaccinated animals tested negative at all times. As opposed to the whole-blood test, no vaccinated group prevailed over the others in the ELISPOT assay.
PCV2-specific proliferation of PBMC
PCV2-specific lymphocyte proliferation was investigated in samples collected at days −27, −7, 21, and 36 with respect to challenge by flow cytometry analyses of intracellular Ki-67 antigen expression. Since Ki-67 flow cytometry test profiles in mock virus-treated PBMC were always found to overlap with those of control isotype in PCV2-treated PBMC, the former were not included into the final analysis. Results are shown in Table 3. All samples tested negative at the beginning of the experiment (day −27), while both vaccination (day −7) and viral challenge (dpc 21) clearly induced a positive response (at least one test-positive pig in every vaccinated group). Interestingly, group 3 showed the greatest prevalence and intensity of positive reactions at any time point analyzed, in agreement with the response in the IFN-γ release assay. Samples taken at day −7 before challenge were also analyzed by a different proliferation assay, based on BrdU incorporation. In this case, only one pig tested positive (pig 4, group 1; data not shown).
High- and low-positive samples are highlighted using bold characters.
Neg, no specific proliferation; Low pos, low specific proliferation (4–10% Ki-67-positive cells); High pos, high specific proliferation (>10% Ki-67-positive cells); Tot, total number of test-positive subjects in each group at the indicated time points.
Discussion
The main goal of this work was to define correlates of protection induced by PCV2 vaccines with a precise Ag payload. This could be in fact a crucial factor underlying the establishment of the protective immunity induced by PCV2 vaccines. In order to determine the Ag mass of whole inactivated PCV2, a PCV2 purification and quantitation system was set up (37) and a PCV2 vaccination/challenge model was established to compare the immunizing effect of three different doses of inactivated, whole-virus PCV2 antigen in the same adjuvant. Under the experimental conditions, the three doses (211–844 ng) of the PCV2 inactivated antigen and the commercial Circovac vaccine were able to induce a protective immune response in pigs. Interestingly, there was no significant correlation between Ab levels at 3 weeks after vaccination, and height and time-course of PCV2 viremia: both ELISA and NA tests failed to predict susceptibility to infection. Also, there was no correlation with viremia resolution (no significant differences between Ab titers of vaccinated and control animals at dpc 29). The response to PCV2 vaccination and challenge in terms of ELISA Ab and NA was similar. The only difference between NA and ELISA Ab profiles was the priming effect of vaccination for the NA response observed at dpc 7, and lacking in control, nonvaccinated pigs. This might be an important difference. Early neutralizing antibody at dpc 7 might induce crucial effector functions (e.g., in vivo neutralization, ADCC) underlying a precocious curtail of PCV2 replication and inhibition of viremia. As opposed to NA, the ELISA tests showed a reduction of the Ab titers at dpc 7 that is possibly due to virus/Ab immunocomplexes sequestered at sites of primary virus replication after challenge. The observed difference between ELISA and NA assays at dpc 7 might be accounted for by the prevailing Ig isotypes and target specificities of the antibodies revealed by the two assays. Further studies are needed to clarify this crucial point. Apart from the early NA response after challenge, the serum Ab profiles could not be considered as predictive prechallenge markers of PCV2 vaccine potency. The seemingly discrepant results obtained in other studies (28,29) could be accounted for by pigs (commercial vs. SPF/germ-free animals), MDA titers at the time of vaccination, type, and timing of the performed Ab assays. In this respect, the choice of employing commercial pigs with MDA at the time of vaccination was dictated by the need for a vaccination protocol as close as possible to the current field scenario. A low-titered, mucosal NA response at day 20 after vaccination could possibly represent another feature of vaccine efficacy, but further investigations are needed to clarify its predictive role.
With regard to the CMI assays performed in this study, preliminary experiments showed that the use of mock virus (cryolysate of PK15c28 cells) as negative control was mandatory to obtain reliable results because of a significant overlapping of virus-specific and nonspecific immune signals. This phenomenon could be due to the recognition of stress antigens by NK and T cells (17). In particular, stress antigens are expressed on PK15 cells and other established pig cell lines of epithelial and endothelial origin (36), as also confirmed in the authors' laboratory (Supplementary Fig. S1; Supplementary Data are available online at
Two different IFN-γ assays were used in this study to investigate CMI in two extremely different contexts: ex vivo entire blood versus separated PBMC. This decision was adopted because the two substrates differ not only for their composition in cytokines and cells, but also for the presence of reactive oxygen metabolites in plasma, as a result of the constitutive oxidative stress in lean type, rapid growth pigs (4). As described elsewhere (23), oxidative stress alters the homeostasis and differentiation of T cells, favoring the polarization into Th2 phenotypes. Therefore, the results obtained from the whole blood IFN-γ release and ELISPOT assays should be considered as complementary approaches to the characterization of the immune response.
Altogether, the herein reported CMI parameters clearly discriminated vaccinated from control animals, and correlated with protection. In particular, the IFN-γ release assay in the recall response to PCV2 inactivated antigen was shown to be a very useful correlate of protection. Based on the results at 3 weeks after vaccination, this assay proved to be very informative with respect to the immunization status of the animals. The CMI tests also showed a tendency of the animals vaccinated with the lowest Ag dose (211 ng) to test positive more clearly than the other vaccinated groups, as though higher amount of inactivated antigen could somehow reduce the immune response. A correct interpretation of this finding and of the whole Ag dose/immune response curve would demand a further comparison between the 211 ng group and another group of animals receiving a much lower Ag dose. Such an antigen extinction test could help identify the minimum antigen dose corresponding to a complete protection of vaccinated pigs.
Most importantly, this work has provided some insights into the interaction of PCV2 with the host's immune system. While PCV2 infection induced immunosuppressive effects in control animals, positive responses in assays of CMI were observed in most vaccinated animals. In particular, PCV2 infection caused a dramatic downregulation of the in vitro IFN-γ response, which is in agreement with a suppressive regulation and/or homing of PCV2-specific responder T lymphocytes from blood to infected tissues. This is also in agreement with the lack of a PCV2-specific IFN-γ response in unvaccinated animals after infection. A partial resumption of the response in vaccinated animals could be appreciated at dpc 36 (see Table 2).
In agreement with previous observations about recovery of CD8+ T cells in blood of PCV2-infected pigs and resolution of viremia (8), vaccination efficacy was probably based on cellular immune mechanisms leading to a protective immunological environment (cytokine-releasing lymphocytes and the like). Such mechanisms could be complemented by effector functions associated with the early neutralizing Ab response of vaccinated pigs to PCV2 infection. Cellular immune mechanisms should be viewed in the framework of the “hidden memories” scenario, that is, the existence of long-lived effector memory T cells, prepositioned at sites of pathogen invasion or mobilized within hours to such sites from blood and marginated pools (27). These T cell populations could check virus replication at the very beginning, thus preventing or curtailing viremia. In this scenario, inactivated chimeric PCV1-2 and PCV2 whole virion vaccines could induce higher levels of PCV2-specific NA and IFN-γ-SC, which correlate with significantly lower viremia levels compared with subunit ORF2 vaccines (35).
Beyond the immune profile observed in the first weeks after vaccination, major issues about the interplay between PCV2 and the host's immune system still remain open. These should be viewed within a historical perspective. After PCV2 vaccines were introduced in pig farms worldwide, strong circumstantial evidence has highlighted their ability to reduce the prevalence of other viral and bacterial diseases on farm significantly (2,12,24). Also, the use of PCV2 vaccines has gone along with a dramatic decline of antibiotic usage in swine herds (5,16,21). These observations lead to two different hypotheses. The former implies that vaccine-induced immune responses check PCV2 infections, meant as a fundamental co-factor for the occurrence of diverse, full-blown diseases sustained by other microbial agents. The latter implies that inactivated PCV2 itself has general immunomodulation properties, associated to ORF2 capsid protein (13,22). This would be in agreement with the results obtained on icosahedral virus-like particles, in particular those consisting of VP60 protein of Rabbit Hemorrhagic Disease Virus (7).
Finally, a caveat should be expressed about the interpretation of the findings. In fact, using the results from a single study to predict a general phenomenon may have some risks. Therefore, it will be of great importance to reproduce these data in other laboratories, and also on ORF2-based and chimeric PCV1-PCV2 ORF2 vaccines. If the predictive potential should be confirmed by different laboratories, the IFN-γ release assay could be recognized as a practical tool to investigate PCV2 vaccine-induced immunity, and might be considered for PCV2 vaccine batch controls as an alternative to challenge infection of animals. This may represent an important step forward in accordance with the current 3Rs policies (18,20) in the framework of a “batch consistency” approach (9).
Footnotes
Acknowledgments
The authors thank C. Mantovani and D. Bonassi (IZSLER, Brescia, Italy) for their skillful technical assistance, and Dr. B. Boniotti (IZSLER, Brescia, Italy) for providing PCV2 viremia data. The Italian Ministry of Health funded this study with grant PRC2011001. The cooperation provided by Dr. G. Leotti and Dr. C. Charreyre (Merial SAS) is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.