
Abstract
It is suggested that the tumor suppressor p53 gene, classified as an interferon-stimulated gene, is implicated in the interferon (IFN)-mediated innate immunity against viruses. This study aimed to examine the transcriptional response of the p53 gene to hepatitis C virus (HCV) infection and IFN-based therapy in chronic hepatitis C (CHC) patients. The study included 65 CHC patients (HCV genotype 1), treated with pegylated IFN-α and ribavirin, and 51 healthy individuals. p53 gene expression was quantified by real-time polymerase chain reaction in peripheral blood mononuclear cells (PBMCs). Analyses were performed before and at weeks 4 and 12 of treatment. p53 gene expression was significantly upregulated in CHC patients compared with healthy controls and at week 4 of therapy. No significant differences in p53 mRNA expression between rapid virologic responders, complete early virologic responders, and nonresponders were observed. No significant correlation was found between p53 gene expression and viral load. The results obtained indicate that HCV infection and IFN-based treatment induces p53 gene transcription in PBMCs. The p53 gene may therefore play a role in HCV infection but is not directly involved in treatment-induced HCV elimination. Moreover, variations in p53 gene expression do not determine on-treatment response in patients with chronic HCV genotype 1 infection.
Introduction
H
Factors contributing to the host's defense against HCV are still poorly understood. Type I IFNs (α/β) released from infected cells establish an antiviral state by binding to specific receptors. This results in activation of the Janus kinase and signal transducer and activator (JAK-STAT) pathway and formation of the IFN-stimulated gene factor 3 (ISGF3) complex. Next, ISGF3 binds to the IFN-stimulated response elements (ISREs) leading to transcriptional induction of IFN-stimulated genes (ISGs) (35,40,43).
Recently, the tumor suppressor p53 gene widely known as “the guardian of the genome,” mediating cell cycle arrest and apoptosis (46), has been classified as an ISG due to the presence of ISREs in its promoter (44). Takaoka et al. (44) showed that p53 is induced in response to viral infection as a downstream transcriptional target of IFN-α/β, indicating a new link between the IFN-α/β signaling pathway and p53 responses in antiviral defense and tumor suppression. The transcriptional role of p53 in IFN-mediated antiviral immunity has been confirmed in vitro and in infected mice (30). In turn, Dharel et al. (7) showed significantly higher levels of HCV RNA replication and viral protein expression, reduced effectiveness of IFN-α treatment in inhibiting HCV RNA replication, and attenuated induction of ISGs in p53-knocked-down HCV-replication cell lines, demonstrating that p53 could have a crucial role in the cellular innate defense against HCV.
Moreover, HCV proteins (NS5A, NS3, core protein) can interact with p53 (6,18,31,39). Lan et al. (18) showed that NS5A protein suppress p53-mediated transcriptional transactivation and apoptosis during HCV infection, which may contribute to hepatocarcinogenesis. Sabri et al. (39) analyzed the interaction of NS3 and NS5A proteins with p53, and they concluded that these HCV proteins play important roles in hepatocarcinogenesis by downregulating expression of the p53 gene. Deng et al. (6) found that NS3 protein plays an important role in hepatocarcinogenesis and inhibits p53 function in an NS3 sequence-dependent manner. Ostuka et al. (31) showed that core protein augmented the transcriptional activity of p53 and increased the expression of p21(waf1) protein, which is a major target of p53.
The aim of this study was to examine the transcriptional response of the p53 gene to HCV infection and to PEG-IFN-α and ribavirin treatment in patients with genotype 1 chronic HCV infection. For this purpose, p53 gene expression was analyzed in healthy individuals and in CHC patients before as well as at weeks 4 and 12 of treatment in combination with viral load analysis.
Materials and Methods
Patients
Sixty-five patients diagnosed with CHC and 51 sex- and age-matched healthy blood donors were included in the study. All patients and healthy donors were of Polish Caucasian origin (Wielkopolska region). They were infected with HCV genotype 1 and were treatment naïve. The exclusion criteria included hepatitis B virus co-infection, human immunodeficiency virus co-infection, coexistent autoimmune liver diseases, or other coexistent chronic liver diseases. Patients received a course of the standard stable doses of PEG-IFN-α2a (PEGASYS®, Roche; 180 or 135 μg per week; n = 29) or PEG-INF-α2b (PEGINTRON®, Schering-Plough; 1.5 μg/kg of body mass per week; n = 36) combined with a weight-based dose of ribavirin (COPEGUS®, Roche, or REBETOL®, Schering-Plough; 1,000 mg per day if body weight was <75 kg or 1,200 mg per day if body weight ≥75 kg). Blood samples were obtained before (day of treatment initiation) as well as at weeks 4 and 12 of treatment.
Written informed consent was obtained from each patient. The study was approved by the local ethnical committee of the University of Medical Sciences in Poznań, Poland (no. 650/12). All experiments were carried put in compliance with the relevant laws and guidelines in accordance with the ethical standards of the Declaration of Helsinki.
Demographic, clinical, and laboratory data of all enrolled patients are summarized in Table 1.
All patients were of Polish Caucasian origin (Wielkopolska region).
Liver biopsy and **Fibroscan (Matavir scoring) were carried out no earlier than 18 months before treatment initiation.
RVR, rapid virologic response; cEVR, complete rapid virologic response; NR, nonresponse; ALT, alanine transaminase; AST, aspartate transaminase; GGT, gamma-glutamyl transpeptidase; AFP, alpha-fetoprotein.
Peripheral blood mononuclear cell preparation, RNA extraction, and cDNA synthesis
Peripheral blood mononuclear cells (PBMCs) were isolated from 5 mL of venous EDTA blood by Histopaque®-1077 (Sigma-Aldrich, St. Louis, MO) gradient centrifugation (1.077 g/mL), washed twice with phosphate-buffered saline, and counted. All samples were processed on the same day or the day after blood collection. Total cellular RNA was extracted from 1 × 106 PBMCs using an Invisorb® Spin Cell RNA Mini Kit (Stratec Molecular, Berlin, Germany) following the manufacturer's instructions. Immediately after isolation, RNA was reverse transcribed into cDNA using M-MLV Reverse Transcriptase, RNase H Minus, Point Mutant (M-MLV RT (H–)) and Random Primers (Promega, Madison, WI). The synthesized cDNA samples were stored at −20°C until use.
p53 gene expression quantification
The mRNA expression level of the p53 gene was determined by real-time quantitative polymerase chain reaction (PCR). β-actin (ACTB) was selected for normalization as the most suitable reference gene, on the basic of its PCR efficiency and low variability under study conditions.
The following p53 gene (GeneBank accession no. NM_000546.5) primers and TaqMan probe were designed: 5′-ATT TGC GTG TGG AGT ATT TGG ATG A-3′ (forward primer), 5′-GTA GTG GAT GGT GGT ACA GTC AGA-3′ (reverse primer), 5′-FAM-CTC AGG CGG CTC ATA GGG CAC C-TAMRA-3′ (p53probe). ACTB primers and fluorescent probe oligonucleotide were described previously (17). The PCR conditions were as follows: one cycle at 95°C for 3 min, followed by 45 cycles at 95°C for 15 sec, 59°C for 15 sec, and 72°C for 30 sec. Reactions were run in triplicate on the iQ5-I-Cycler (Bio-Rad), and the mean threshold cycle (Ct) value for each sample was analyzed when SD < 0.5.
HCV genotyping and HCV RNA quantification
Viral RNA was extracted from the serum using an Invisorb® Spin Virus RNA Mini Kit (Stratec Molecular). HCV genotyping was performed using a VERSANT® HCV Genotype 2.0 Assay (LiPA; Siemens Medical Solution Diagnostics, Tarrytown, NY), according to the manufacturer's protocol. Serum HCV RNA qualitative and quantitative detections were performed by reverse transcription PCR technique using a GeneProof HCV PCR Kit (GeneProof, Brno, Czech Republic; limit of detection 36.173 IU/mL).
Data analysis
p53 gene expression was expressed in relative quantification (RQ = 2−ddCt = 2−[(Ct p53 − Ct ACTB)patient − (Ct p53 − Ct ACTB)calibrator], pooled cDNA from healthy controls was used as a calibrator). The fold change between weeks of treatment and baseline was calculated using the formula: RQ = 2−ddCt = 2(Ct p53 − Ct ACTB)week − (Ct p53 − Ct ACTB)baseline. The results were given as median and range. The Mann–Whitney U-test was used to compare p53 gene expression or viral loads between independent groups. The Wilcoxon matched-pair signed-rank test was performed for comparisons between weeks of treatment within groups. Correlation analysis was conducted using Spearman's rank correlation. All calculations were performed using SPSS Statistics for Windows v22.0 software (IBM Corp., Armonk, NY). Results were considered statistically significant at p < 0.05.
Results
p53 mRNA expression in PBMCs from CHC patients and healthy individuals
In order to examine whether the p53 gene is upregulated in response to HCV infection, p53 gene expression was analyzed in patients with CHC and in healthy donors.
Median p53 gene expression values for HCV-infected patients and healthy individuals were 1.50 (range 0.25–5.77) and 1.05 (range 0.25–3.58), respectively. It was found that the baseline pretreatment level of p53 mRNA expression was significantly upregulated in CHC patients compared with healthy controls—significant 1.43-fold upregulation, up to 22.98-fold when the highest expression level noted in CHC group and the lowest value in control group were compared (p = 0.0028; Fig. 1).
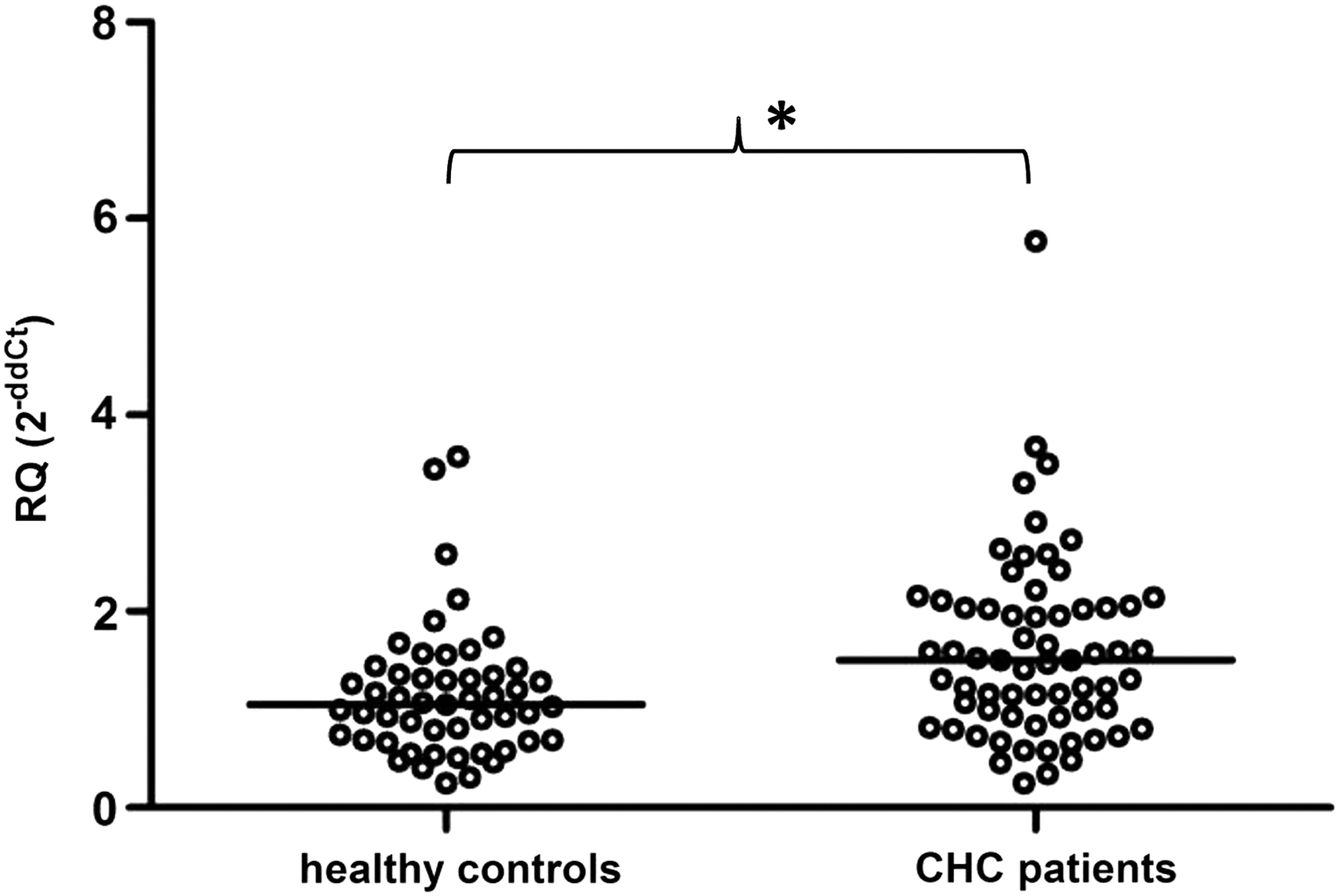
p53 gene expression in peripheral blood mononuclear cells (PBMCs) from healthy blood donors and 65 chronic hepatitis C (CHC) patients prior to the initiation of pegylated interferon alpha (PEG-IFN-α) and ribavirin treatment. p53 gene expression was significantly upregulated in patients (n = 65) compared with healthy controls (n = 51; p = 0.0028). The p53 mRNA expression level was calculated using the formula: RQ = 2−ddCt = 2−[(Ct p53 − Ct ACTB)patient − (Ct p53 − Ct ACTB)calibrator], as calibrator pooled cDNA from healthy controls was used. Single cases are reported. Median p53 gene mRNA values are indicated for each group. *p < 0.05.
p53 gene expression in CHC patients during treatment with PEG-IFN-α and ribavirin
To assess whether exogenous interferon affects p53 mRNA expression in PBMCs from HCV-infected patients, levels of p53 gene expression at week 0 (baseline; prior to the initiation of treatment), week 4, and week 12 of IFN-based therapy were compared (Fig. 2).
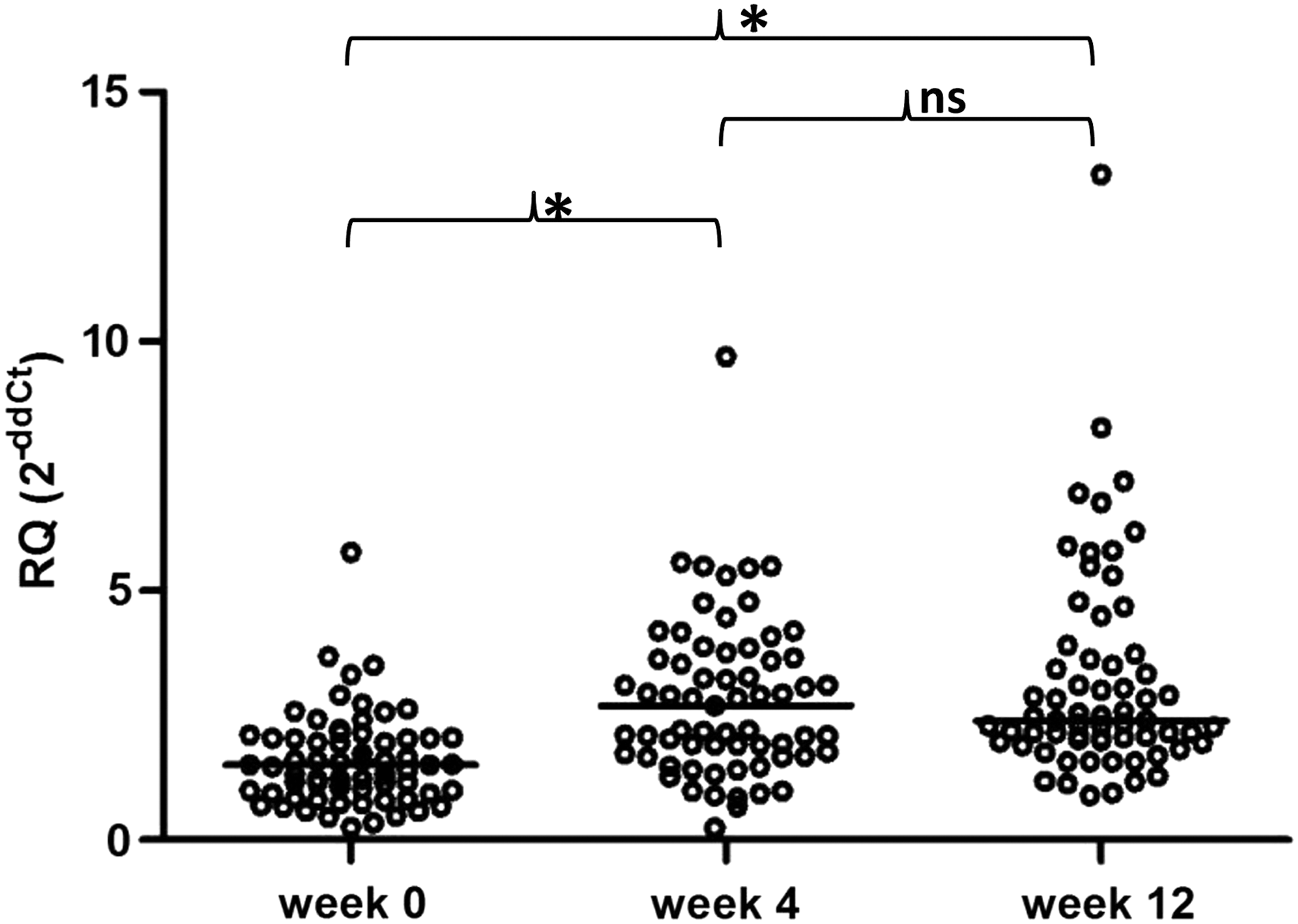
p53 gene expression in PBMCs from CHC patients prior to the initiation and during PEG-IFN-α and ribavirin treatment. p53 gene expression was significantly upregulated at weeks 4 and 12 of therapy compared with the baseline (p < 0.0001). No significant difference in p53 gene expression between weeks 4 and 12 was observed. The p53 mRNA expression level was calculated using the formula: RQ = 2−ddCt = 2−[(Ct p53 − Ct ACTB)patient − (Ct p53 − Ct ACTB)calibrator], as calibrator pooled cDNA from healthy controls was used. Single cases are reported. Median p53 gene mRNA values are indicated for each group. *p < 0.05; n.s., not significant.
p53 gene expression level was found to be significantly upregulated at week 4 (median 2.69; range 0.25–9.70) compared with the baseline level (median 1.50; range 0.25–5.77; p < 0.0001). The median increase (fold change) was 1.73-fold (range 0.96–7.62). To examine whether pretreatment p53 mRNA expression may determine the extent of p53 gene induction, the baseline level was correlated with this fold increase, but no significant association was found.
Moreover, at week 12 of treatment, p53 mRNA expression showed a significant increase compared with the baseline (p < 0.0001; week 12 median 2.39; range 0.89–13.34). The median increase was 1.64-fold (range 0.95–9.85). A significant difference in p53 gene expression between week 4 and week 12 was not observed.
p53 gene expression and response to IFN-based therapy in CHC patients
A significant increase of p53 gene expression at week 4 of treatment prompted an analysis of whether p53 gene expression is associated with a response to anti-HCV therapy (Fig. 3).
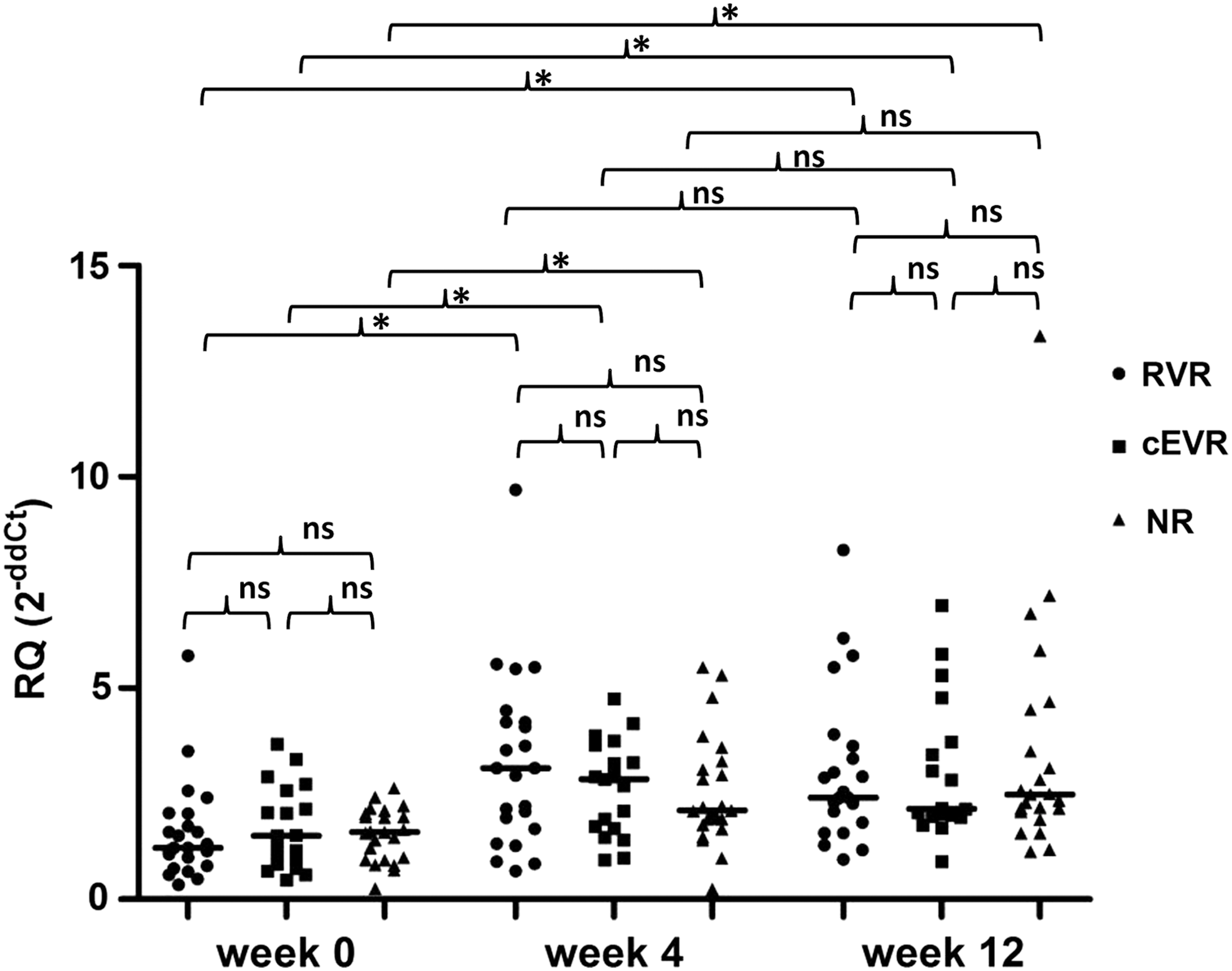
p53 gene expression at baseline, week 4, and week 12 of PEG-IFN-α and ribavirin therapy in three response groups of CHC patients. No significant differences in pretreatment level (week 0) of p53 expression between the rapid virologic response (RVR; n = 23), complete early virologic response (cEVR; n = 19), and nonresponse (NR; n = 23) groups were found. Moreover, there were no significant differences between RVRs, cEVRs, and NRs in p53 gene expression at week 4 and at week 12 of treatment. In each group, significant upregulation was observed at week 4 compared with the baseline (p < 0.0001) and at week 12 compared with the baseline (p = 0.0004, p = 0.0007, and p = 0.0001, for RVRs, cEVRs, and NRs, respectively). No significant changes in p53 mRNA expression were observed between weeks 4 and 12 in any group. The p53 mRNA expression level was calculated using the formula: RQ = 2−ddCt = 2−[(Ct p53 − Ct ACTB)patient − (Ct p53 − Ct ACTB)calibrator], as calibrator pooled cDNA from healthy controls was used. Single cases are reported. RVRs, cEVRs, and NRs are represented by circles, squares, and triangles, respectively. Median p53 gene mRNA values are indicated for each group. *p < 0.05; n.s., not significant.
Patients were classified into three groups based on on-treatment serum viral load kinetics. Among the 65 CHC patients, 23 (35.4%) had an undetectable level of HCV RNA at week 4 of treatment and were classified into the rapid virologic response (RVR) group. Among the others who were HCV-RNA positive at week 4 of treatment, 19 (29.2%) showed an undetectable viral RNA level at week 12 and were classified as complete early virologic responders (cEVRs), while 23 (35.4%) showed a detectable HCV-RNA level and were classified into the nonresponse (NR) group. The NR group consisted of six partial early virologic responders (pEVRs) who had at least a 2-log decline in viral load in serum at week 12 of therapy compared with their baseline level, as well as 17 patients who showed either no decline or a decline of less than 2-log (null response group). There were no significant differences in baseline serum HCV-RNA level between these three response groups (median 6.42 (0.15–220.00) × 104 IU/mL; 6.18 (0.14–91.00) × 104 IU/mL; 6.50 (0.11–42.00) × 104 IU/mL in the RVR, cEVR, and NR groups, respectively). Dividing patients into these groups, according to on-treatment kinetics of serum HCV-RNA, enabled changes in p53 gene expression and viral load to be tracked precisely and in parallel between baseline, week 4, and week 12 of anti-HCV therapy.
Median values of baseline p53 expression in the RVR, cEVR, and NR groups were as follows: 1.22 (0.35–5.77), 1.50 (0.46–3.68), and 1.59 (0.25–2.64). No significant differences in the pretreatment level of p53 mRNA expression was found between these three groups. Moreover, there were no statistically significant differences between RVRs, cEVRs, and NRs when p53 gene expression was analyzed at week 4—medians: 3.11 (0.67–9.70), 2.85 (0.93–4.75), and 2.11 (0.24–5.50) for RVRs, cEVRs, and NRs, respectively—as well as at week 12 of treatment—medians: 2.41 (0.94–8.27), 2.14 (0.89–6.96), 2.48 (1.13–13.34) for RVRs, cEVRs and NRs, respectively.
Next, changes in p53 gene expression during treatment in each response group were analyzed. RVRs, cEVRs, and NRs showed significant upregulation of p53 gene expression at week 4 compared with week 0 (p < 0.0001 for RVRs and NRs; p = 0.0003 for cEVRs) and at week 12 compared with week 0 (p = 0.0004, p = 0.0007, and p = 0.0001 for RVRs, cEVRs, and NRs, respectively). Furthermore, no significant changes were observed in p53 mRNA expression between week 12 and week 4 in the studied groups.
Considering an increase in p53 gene expression at week 4 relative to week 0, it was found that there are no significant differences between response groups—median increases: 1.95-fold (1.08–7.62), 1.32-fold (0.97–4.08), and 1.729-fold (0.96–2.48) for RVRs, cEVRs, and NRs, respectively.
Additionally, the association between p53 gene expression level and HCV viral load was analyzed. It was found that pretreatment serum viral load is not related to baseline p53 gene expression and that there is no statistically significant correlation between p53 expression and HCV-RNA level at week 4 or at week 12 of treatment. Moreover, it was observed that changes in expression of p53 gene (fold change) are not associated with corresponding changes in HCV RNA levels.
Discussion
The present study aimed to examine the transcriptional response of the p53 gene to HCV infection and to PEG-IFN-α and ribavirin therapy in patients chronically infected with HCV genotype 1. To the authors' knowledge, it is the second study to analyze changes in p53 gene mRNA expression during IFN-α and ribavirin therapy in CHC patients, and the first to examine these changes in patients chronically infected with HCV genotype 1.
In this study, baseline p53 mRNA expression level was significantly higher in the PBMCs of CHC patients compared with healthy controls. This finding is consistent with results of a previous study of patients with chronic HCV genotype 4 infection (4 patients; 12 controls) conducted by Hamdi et al. (15). Upregulation of p53 expression, as an interferon-stimulated gene, in studied patients may be explained by increased production of endogenous IFN during HCV infection. It has been repeatedly found that the endogenous type 1 IFN antiviral effector pathway is activated in chronically HCV-infected individuals prior to administration of therapeutic IFN-α (3,8,13,14,25,33,42). In the study conducted by Loguerico et al. (22), p53 protein was found to be overexpressed in 7/40 (17.5%) liver biopsy samples. It was observed that p53 overexpression can occur, even in the early stages of HCV-related liver disease. Importantly, they found the change in p53 expression to be specifically related to HCV infection but not to HBV infection, as p53 overexpression was not observed in HBV patients.
In the present study, p53 mRNA expression was also found to be significantly upregulated after initiation of anti-HCV treatment (week 4) compared with the baseline level, which further confirms that IFN-α affects p53 gene expression in PBMCs of CHC patients. Furthermore, the increase in p53 expression may be amplified by ribavirin. It was found that ribavirin may stabilize and activate p53 through the ERK1/2 pathway, which may contribute to the enhanced anti-HCV response to IFN-α and ribavirin (21).
Upregulation of p53 gene expression under the influence of endogenous and exogenous IFN suggests the role of this gene in mediating interferon's antiviral effects against HCV. In vitro analysis has shown that the tumor suppressor gene p53 is involved in the host's defense against HCV by modulating interferon signaling (7). Moreover, Hamdi et al. (15) indicated the existence of an association between p53 gene mRNA expression and the expression of genes coding the myxovirus-resistance protein A (MxA) and double-stranded RNA (dsRNA)-activated protein kinase (PKR)—two ISGs that play an important role in antiviral defense. They observed that high p53 gene mRNA expression is accompanied by upregulated MxA gene and PKR gene mRNA expression, whereas low p53 gene mRNA expression is not. Their study supports the previous in vitro research regarding a possible contribution of p53 to the host's antiviral response (7,30,44).
The effect of CHC therapy on p53 expression was measured at the protein level in liver biopsy specimens. Panasiuk et al. (32) showed that IFN-α2b treatment of CHC patients caused an increase in the percentage of p53-positive hepatocytes and the intensity of p53 protein accumulation. Moreover, they concluded that increased p53 protein expression may result from the effect of the increase in regeneration or apoptosis of HCV-infected hepatocytes, and may be considered as positive prognostic marker of the treatment effectiveness and HCV elimination.
Establishment of ISG expression as a useful molecular marker of response to antiviral treatment in CHC patients is a matter of debate. It is suggested that nonresponsiveness to IFN-based treatment in CHC patients is associated with pretreatment upregulation of ISGs such as MxA or PKR (13,24,42). Thus, CHC patients who show a strong induction of ISGs prior to treatment do not achieve a virologic response. In addition, compared with responders, these patients were not able to increase the transcription of ISGs further after the administration of exogenous IFN-α (41). Nonetheless, there are some analyses in which the above-mentioned associations were not observed (25,28).
In the present study, the pretreatment level of p53 gene expression was not associated with the response to anti-HCV IFN-based treatment. Moreover, it was observed that the baseline p53 gene expression level did not determine the extent of its induction after administration of exogenous IFN-α. In addition, no differences in p53 mRNA level at baseline, week 4, or week 12 of treatment were observed between the response groups. Hence, it can be concluded that p53 gene expression level in the PBMCs of CHC patients does not allow individuals with RVR, cEVR and NR to be distinguished, and thus it is not useful in predicting on-treatment response to anti-HCV therapy. Furthermore, no statistically significant correlation was found between p53 mRNA expression and serum HCV RNA level. These results confirm the lack of direct association between p53 gene expression in PBMCs and treatment-induced HCV elimination.
Similarly, Hamdi et al. (15) demonstrated that the baseline p53 gene expression level does not allow the response to treatment with PEG-IFN-α and ribavirin to be predicted in patients with chronic HCV genotype 4 infection. The authors showed no significant difference in pretreatment p53 expression between EVR patients (decrease in HCV RNA levels by at least 2 log10 at week 12 of treatment, n = 23) and non-EVR patients (no decline or a decline of less than 2 log10 in basal viral load at week 12 of therapy, n = 7) (15). In the present study, when patients were divided in the same way as in Hamdi et al.'s study (15), their results were confirmed (data not shown). Additionally, the p53 expression level was compared between RVRs (strong positive predictor for SVR) (9,37) and non-RVRs, as well as between NRs (strong negative predictor for SVR) (5) and non-NRs to confirm the lack of usefulness of p53 expression in predicting the response to anti-HCV therapy (data not shown). The association between p53 expression and the response to anti-HCV treatment was analyzed by Loguercio et al. (22) at the protein level in liver samples. In this study, a different response to IFN-α was observed between p53-positive and p53-negative patients (4/33 p53-negative patients achieved SVR; all seven p53-negative patients were NRs). However, the study only included a small number of cases. Therefore, firm conclusions on this observation cannot be drawn.
Some factors that may influence the obtained results need to be discussed. It cannot be excluded that HCV genotype and ethnicity—two factors that are associated with variable treatment response to PEG-IFN-α and ribavirin treatment in CHC patients (11,38)—may affect the p53 gene mRNA expression level. Little is known about differences in ISG expression among CHC patients infected with different HCV genotypes, and to the authors' knowledge, there are no studies that compare p53 gene mRNA expression among CHC patients infected with different HCV genotypes. In a study conducted on chimpanzees, it was found that animals infected with HCV genotype 3a exhibited a reduced level of intrahepatic ISGs expression compared with animals infected with HCV genotype 1. However, the study included only one animal with HCV genotype 3a. Therefore, results must be interpreted with caution (2). Furthermore, in the study conducted by Sarasin-Filipowicz et al. (41), it was observed that the ISG expression level, measured in pretreatment liver biopsies, was higher in CHC patients with HCV genotypes 1 and 4 compared with genotypes 2 and 3 and higher in non-rapid responses compared with rapid responders, independently from the HCV genotype. They concluded that this relationship provides an explanation for the poor response to IFN therapy in patients infected with HCV genotypes 1 and 4. It cannot be excluded that similar associations would be observed in the case of the p53 gene.
In addition, it can be considered that patients infected with different HCV genotypes may exhibit different response to ISGs, including p53. As an example, the response to the action of OAS, one of the ISGs, differs in patients with different HCV genotypes (HCV genotypes 1a and 1b compared with 2a, 2b, 3a, and 3b have a lower frequency of UA and UU dinucleotides, after which RNase L, stimulated by the protein product of OAS, cleaves RNAs) (16). It can only be assumed that the effect on p53 gene antiviral action may be different between patients with HCV genotype 1 and other HCV genotypes (especially with HCV genotypes 2 and 3 of confirmed higher IFN-responsiveness). Perhaps patients infected with HCV genotypes 2 or 3 could have a lower pretreatment level of p53 gene mRNA and exhibit a better response to p53, resulting in more efficient HCV elimination. However, the complexity of the response to IFN in HCV-infected patients does not allow firm conclusions to be drawn, even when the above-mentioned correlations are observed. In vitro functional studies are needed to analyze further the association between p53 gene expression and HCV genotype.
Moreover, it is possible that the genetic diversity among populations may affect the mRNA changes of ISGs, including the p53 gene. It was found that that relationship between ISGs expression (2,5-OAS, MxA) and the response to IFN-based treatment differed by race/ethnicity (non-Hispanic black vs. non-Hispanic white American patients infected with HCV genotype 1) (23). Furthermore, some SNPs, differentially distributed among populations, may affect the mRNA changes of ISGs. For example, it was found that IFNL3 (IL28B) polymorphism (rs12979860), a critical predictor of treatment-induced HCV clearance, whose prevalence differs among ethnic groups (12), may affect mRNA expression level of ISGs (perhaps also p53 gene expression). It was observed that genotype CC of rs12979860, related with a higher rate of SVR in CHC patients, is associated with a lower level of intrahepatic ISG expression (45).
It cannot be excluded that the association between p53 mRNA level and the response to IFN-based therapy in CHC patients would be observed if examining p53 mRNA expression before week 4 of treatment. Analyses of p53 gene expression in a shorter time after administration of the IFN-α injection could allow sharp p53 mRNA enhancement to be recognized, because it is possible that after few weeks, there is a blurring of the differences between patients with different responses.
Moreover, perhaps it would be possible to observe some association between p53 expression and the response to anti-HCV therapy if p53 gene expression was examined in liver biopsy specimens. Whether the ISG expression pattern may differ between PBMCs and the liver is a matter of debate. It has been suggested that PBMCs might not be representative of what happens in infected hepatocytes and that predictors derived from PBMC transciptome may reflect the response of the host rather than processes dependent on viral factors (41). However, some authors have demonstrated a close correlation in ISG expression between PBMCs and the liver (1,13), and they even observed a significant induction of ISGs (MxA, PKR) in PBMCs but not in the liver (27). In the present study, we examined p53 gene expression in PBMCs because they are strong IFN producers (4) and even stronger than hepatocytes (29), as well as the fact that PBMCs have been shown to be a reservoir for HCV replication (34). Moreover, by the use of peripheral blood, which in contrast to liver specimens is readily available biological material, it was possible to keep track of changes in p53 gene expression during anti-HCV treatment.
In summary, this study has demonstrated that expression of the p53 gene is upregulated in PBMCs from CHC patients compared with healthy controls. Moreover, it has found that IFN-based treatment induces an increase of p53 gene transcription, but there is no association between p53 gene expression and HCV elimination during PEG-IFN-α and ribavirin treatment. On the basis of the obtained results, the p53 gene may play a role in HCV infection, but it is probably not directly involved in treatment-induced HCV elimination. Because the p53 gene transcriptional response to exogenous IFN-α was similar in responders and non-responders, it seems that variations in p53 gene expression in PBMCs do not determine the on-treatment anti-HCV response in patients with chronic HCV genotype 1 infection. Further studies with a larger number of patients are needed to confirm these findings.
Footnotes
Acknowledgments
This work was supported by The National Science Centre, Poland; grant no. UMO-2011/01/B/NZ6/04258.
Author Disclosure Statement
No competing financial interests exist.