
Abstract
Human immunodeficiency virus (HIV)-specific CD8+ T cell dysfunction parallels disease progression; therefore, restoring potent HIV-specific CD8+ T cell responses is a key therapeutic goal. Certain CD8+ T cell peptide epitope variants, termed heteroclitic, enhance cytokine production by the HIV-specific CD8+ T cells of some individuals. In this study, we investigated whether heteroclitic peptides that enhance cytokine production by HIV-specific CD8+ T cells also reduce functional and phenotypic evidence of HIV-specific CD8+ T cell exhaustion in those instances. Twenty-four variant peptides of human histocompatibility-linked leukocyte antigen (HLA)-A2-restricted reference HIV peptide epitopes designated as A2-7; Nef 83→91, A2-8; Nef 135→143, A2-Gag; Gag 77→85 and A2-9; Gag 433→440 were synthesized with conservative and semiconservative amino acid substitutions at positions 3, 5, and 7 or 3, 5, and 8 of Gag 433→440. Variants that enhanced interferon-gamma (IFN-γ) and/or interleukin-2 (IL-2) production in enzyme-linked immunospot assays (29 cases overall) were subsequently tested by 7-day in vitro peptide stimulation for their effects on HIV-specific CD8+ T cell proliferation and programmed death-1 (PD-1) expression. Heteroclitic variants enhanced HIV-specific CD8+ T cell proliferation by >20% in 13/29 cases tested, reduced PD-1 expression on proliferating cells by 15–50% in 10 cases, and reduced PD-1 expression on proliferating cells by >50% in 3 cases. In five cases, the same heteroclitic peptide increased proliferation by >20% and reduced PD-1 expression by >15%. These data demonstrate that heteroclitic peptides can alter the magnitude and character of HIV-specific CD8+ cell responses relative to reference peptides and may have a unique immunotherapeutic value in therapeutic vaccines.
Introduction
E
Although viral persistence appears to drive antigen-specific CD8+ T cell impairment and increased PD-1 expression, viral clearance is insufficient to restore the effector functions of lymphocytic choriomeningitis virus-specific CD8+ T cells in a murine model of chronic infection and likewise, combination antiretroviral therapy (cART) fails to completely restore HIV-specific CD8+ T cell functions, despite effective long-term suppression of HIV replication (18,23,24,27). Therefore, some form of therapeutic intervention to reconstitute CD8+ T cell functions or induce stronger, more persistent HIV-specific CD8+ T cell responses is necessary as an alternative or complement to cART for HIV treatment and eradication strategies.
Heteroclitic peptides are sequence variants of native peptide epitopes that stimulate T cell responses in some way superior to those stimulated by the native epitope itself (8,31). Amino acid (aa) substitutions in heteroclitic peptides can enhance T cell responses by increasing the peptide-binding affinity for human histocompatibility-linked leukocyte antigens (HLAs) or by modifying interactions between the T cell receptor (TCR) and the peptide/HLA complex (3,22). These changes can have significant impacts in vivo as in experimental models, therapeutic vaccination with heteroclitic peptides activated CTL responses capable of eradicating tumor cells or virus-infected cells that persisted in the face of the immune response mounted against native viral or tumor-associated self-antigens (8,31). In the same manner, incorporation of heteroclitic peptides into prophylactic or therapeutic vaccines could potentially induce more effective T cell responses against chronic human pathogens and tumor antigens.
We previously showed that conservative or semiconservative aa variation at positions 3, 5, and 7, sites within HLA-class I-restricted peptide epitopes predicted to interact with the TCR, can generate heteroclitic HIV peptides that trigger stronger cytokine responses by HIV-specific CD8+ T cells cross-reactive with the reference peptide (10). However, we did not test the effects of these heteroclitic peptides on other important HIV-specific CD8+ T cell functions such as proliferation and differentiation. In this study, we investigated the effects of heteroclitic peptides on HIV-specific CD8+ T cell proliferation and on expression of PD-1, an inhibitory cell surface protein associated with T cell dysfunction and exhaustion, by comparing 7-day in vitro responses to stimulation with heteroclitic versus reference peptides.
Materials and Methods
Study subjects
This study was approved by the Newfoundland and Labrador Provincial Health Research Ethics Authority. Written informed consent for whole blood collection and immunological studies was obtained from all participants. Individuals infected with HIV-1 and apparently healthy non-HIV-infected volunteers were recruited from the Newfoundland and Labrador Provincial HIV clinic and from the Faculty of Medicine personnel, Memorial University of Newfoundland, St. John's, Canada, respectively. Most HIV-infected study subjects had their plasma HIV viral load suppressed with cART to below clinical detection limits at the time of participation.
Peripheral blood mononuclear cells (PBMC) were isolated by density gradient centrifugation with Ficoll-Paque (GE Healthcare Bio-Sciences, Piscataway, NJ) from fresh whole blood collected by venipuncture into vacutainer tubes containing acid-citrate-dextrose anticoagulant and diluted 1:2 with phosphate-buffered saline (PBS), pH 7.4. The isolated PBMC were used fresh or cryopreserved in a lymphocyte medium (RPMI 1640 supplemented with 10% fetal calf serum [FCS], 100 μg/mL streptomycin, 100 IU/mL penicillin, 2 mM
Heteroclitic peptide effects were only tested when cells stimulated with the reference and heteroclitic peptide(s) were either both cryopreserved or both freshly isolated from the same time point. Expression of HLA-A2 was determined with the anti-HLA-A2 monoclonal antibody (mAb) NFLD.M2 (a gift from Dr. S. Drover, Memorial University of Newfoundland) plus fluorescein isothiocyanate-conjugated goat anti-mouse IgG (Jackson Immunoresearch Labs, West Grove, PA) and confirmed with the Lambda Monoclonal Typing Tray Second HLA Class I, Lot #6A (One Lambda, Canoga, CA), as per the manufacturer's instructions (7).
Peptides
Subjects with CD8+ T cell responses against reference HIV peptides were identified as previously described (10). Four consensus clade B, immunodominant, HLA-A2-restricted, and optimally defined HIV peptide epitopes commonly recognized by HIV-infected subjects in our cohort were chosen as reference peptides and a series of six sequence variants were synthesized per peptide. Twenty-four variants were synthesized overall with either conservative or semiconservative aa substitutions at positions 3, 5, or 7 of peptides designated as A2-7; Nef 83→91, A2-8; Nef 135→143 and A2-Gag; Gag 77→85 or positions 3, 5, and 8 of A2-9; Gag 433→440 (Table 1). Synthetic HIV peptides 95% pure (EZBioLab, Carmel, IN) were dissolved in DMSO at 10 mg/mL, and stock solutions of 1 mg/mL made by dilution with unsupplemented RPMI 1640.
Amino acid changes from reference peptide sequence are in bold text.
aa, amino acid.
ELIspot assays
Enzyme-linked immunospot (ELIspot) plates (Millipore, Bedford, MA) were activated by treating with 35% ethanol, washed four times with PBS, and coated overnight at 4°C with 7.5 μg/mL IFN-γ mAb 1-D1K or 15 μg/mL IL-2 mAb IL-2-1/249 in PBS (Mabtech, Cincinnati, OH). Plates were blocked with PBS containing 1% FCS before dispensing 2×105 cells per well in the lymphocyte medium in duplicate for IFN-γ or 4×105 cells per well for IL-2 ELIspot, respectively. Peptides were added to 4 μg/mL. Unstimulated PBMC served as a negative control while cells stimulated with 4 μg/mL phytohemagglutinin (MP Biomedicals, Santa Ana, CA) served as a positive control. The ELIspot plates were incubated overnight at 37°C in a 5% CO2 incubator. IFN-γ and IL-2 spot-forming units (SFU) were detected by adding 100 μL/well of either 1 μg/mL biotinylated anti-IFN-γ mAb 7-B6-1 or anti-IL-2 mAb IL-2-II (Mabtech) and incubating for 2 h at room temperature. Plates were washed and 100 μL/well streptavidin-alkaline phosphatase (ALP) conjugate (Mabtech), diluted 1:1,000 in PBS with 0.5% FCS, was added for 1 h. Plates were washed again and 100 μL/well of a 1/100 diluted chromogenic ALP substrate (Bio-Rad Laboratories, Hercules, CA) in a color development solution was added. Color development was stopped after 30 min by rinsing plates with tap water. Plates were air-dried overnight and spots counted with an ImmunoScan ELIspot reader (Cellular Technology Ltd., Cleveland, OH). Wells producing at least twice the number of spots as observed in the background control wells and ≥50 IL-2 SFU/106 PBMC or ≥100 IFN-γ SFU/106 PBMC over background were considered to indicate positive responses to peptide stimulation.
Flow cytometry
Carboxyfluorescein succinimidyl ester (CFSE) was used to monitor CD8+ T cell division based on sequential halving of the fluorescence intensity of each successive daughter cell generation. For labeling, PBMC were incubated in the dark at 37°C for 10 min at 1×106 cells/mL in PBS supplemented with 5 mM ethylenediaminetetraacetic acid (Sigma-Aldrich) and 0.25 μM CFSE (Invitrogen). Cell labeling was quenched by adding five volumes of ice-cold lymphocyte medium followed by incubation on ice for 5 min. Cells were then washed three times with the lymphocyte medium, resuspended at 2×106 cells/100 μL lymphocyte medium, and stimulated for 1 h at 37°C with 100 μL of peptide at 200 μg/mL for a final peptide concentration of 100 μg/mL (∼100 μM). Cells were then resuspended at 1×106/mL in the lymphocyte medium and maintained in culture for 7 days in a 37°C, 5% CO2, humidified incubator.
In some cultures, the PD-1 blocking antibody (clone EH12.2H7; BioLegend, San Diego, CA) was added at 5 μg/mL. After harvesting, cells were stained with the anti-CD8-peridinyl chlorophyll protein (BW135/80; MiltenyiBiotec, San Diego, CA) and anti-PD-1-allophycocyanin, if anti-PD-1 was not used in the culture. Appropriate isotype controls were included. Cells were analyzed on a FACSCalibur™ Cell Analyzer (BD Biosciences, San Jose, CA) and results visualized by representation of CFSE intensity versus PD-1 expression on live CD8+ lymphocytes.
Statistical analyses
All statistical analyses were performed using the GraphPad Prism (version 5.0) statistical software package (Graph-Pad Software, San Diego, CA). A significant difference in the percentage of responding cells expressing PD-1 following heteroclitic peptide stimulation versus reference peptide was assessed with the Wilcoxon signed-rank test. This nonparametric test was used to compare paired values of the percentage of responding cells expressing PD-1 for each of the 29 cases where heteroclitic and reference peptide stimulation was carried out.
Results
Identification of heteroclitic peptide variants
We screened PBMC from 25 HLA-A2 expressing HIV-1-infected individuals by ELIspot assay for reactivity against one or more of the four peptide sets that were synthesized. A general summary of the number of positive responses is provided in Table 2. After background subtraction, variant peptides that stimulated ≥100 more IFN-γ SFU/106 PBMC or ≥50 more IL-2 SFU/106 PBMC than the corresponding reference peptides were considered heteroclitic, provided that the total SFU/106 PBMC was ≤1,000 for IFN-γ or ≤500 for IL-2. In the case of responses ≥1,000 IFN-γ SFU/106 PBMC or ≥500 IL-2 SFU/106 PBMC, variant peptides were considered heteroclitic if they stimulated ≥10% more IFN-γ or IL-2 SFU/106 PBMC than the reference peptide.
ELIspot, enzyme-linked immunospot;
We identified a total of 29 instances of the heteroclitic peptide activity: 6 for A2-7, 10 for A2-8, 6 for A2-Gag, and 7 for A2-9 (Table 3). For each of A2-8, A2-Gag, and A2-9, there were five different subjects where one or more of the variant peptides were heteroclitic, while there were three such subjects for A2-7. The heteroclitic nature of particular variant peptides was not generalizable, as the particular peptide variant or variants with the heteroclitic activity often differed between individuals responding against the same reference peptide. While the possibility of any one variant being heteroclitic in a particular instance was small, even with the limited set of variant peptides we synthesized (two aa variants at each of three sites), at least one of the six variant peptides/set had heteroclitic properties between 30% and 40% of the time. In addition, there were a number of cases where more than one variant peptide within the set increased cytokine secretion relative to the reference peptide, indicating that the heteroclitic peptide variant phenomenon itself is relatively common (Table 3).
Numbers in bold text denote cases where either cytokine production or proliferation was increased or PD-1 expression reduced by the indicated (heteroclitic) peptide relative to the reference peptide.
Percent proliferation represents the percentage of total CD8+ T lymphocytes that proliferated over 7 days of stimulation with the indicated peptide.
Percent PD-1 expression represents the percentage of the CD8+ T cells that proliferated over 7 days of stimulation with the indicated peptide that express PD-1.
HIV, human immunodeficiency virus; PBMC, peripheral blood mononuclear cells; PD-1, programmed death-1; SFU, spot-forming units.
Heteroclitic peptides enhance HIV-specific CD8+ T cell proliferation relative to reference peptides
One objective of this study was to determine if heteroclitic HIV peptides that augment HIV-specific CD8+ T cell cytokine production also enhance HIV-specific CD8+ T cell proliferation. To test this, PBMC from the same individual with whom the heteroclitic properties were identified by increased cytokine production in ELIspot assays were stimulated individually with the reference peptide and the peptide or peptides previously identified as heteroclitic. After 7 days of in vitro stimulation, the percentage of proliferating (CFSEdim) CD8+ T cells was measured by flow cytometry. Proliferation was considered to be enhanced if heteroclitic peptides stimulated a ≥20% increase in the percentage of CFSEdim CD8+ T cells relative to the percentage of CFSEdim CD8+ T cells stimulated by reference peptide.
By this criterion, heteroclitic peptides enhanced CD8+ T cell proliferation in 13/29 cases tested (representative examples are shown in Fig. 1C, D, F) and stimulated similar CD8+ T cell proliferation to the reference peptides in a further seven cases. Therefore, variant peptides identified as heteroclitic by their ability to increase cytokine production, stimulated superior or equivalent proliferation of HIV-specific CD8+ T cells compared to the reference HIV peptide epitope in 20/29 cases tested.

Comparison of HIV-specific CD8+ T cell proliferation and PD-1 expression stimulated by reference and heteroclitic peptides. After 7 days of stimulation with the peptides listed above the plot frames, live CD8+ cells were gated for analysis as in the upper left plot
Heteroclitic peptides reduce expression of PD-1 on proliferating HIV-specific CD8+ T cells relative to reference peptides
Expression of PD-1 on HIV-specific CD8+ T cells is associated with their functional impairment and often considered a sign of T cell exhaustion (23,29). In vivo, expression of PD-1 on HIV-specific T cells is related to both the plasma virus load and the viral epitope recognized by particular HIV-specific T cells. Clonotypically distinct T cells recognizing the same viral epitope can also differ in PD-1 expression, indicating that slight variations in TCR interactions can affect the level of PD-1 upregulation (14). To test whether stimulation with heteroclitic peptides affected the phenotype of HIV-specific CD8+ T cells and potentially recruited responder cells with less sign of functional impairment, we compared the percentage of proliferating CD8+ T cells expressing PD-1, following stimulation with reference versus heteroclitic HIV peptides.
The percentage of proliferating CD8+ T cells expressing PD-1 in response to heteroclitic peptide stimulation was reduced by >15% relative to reference peptide stimulation in 13/29 cases (representative examples are shown in Fig. 1D, F). To confirm that PD-1 affected CD8+ T cell dysfunction in our system as illustrated by reduced proliferation, we added a blocking antibody against PD-1 to in vitro cultures stimulated with either a reference peptide or variant peptide identified as heteroclitic. As shown in Figure 2, HIV-specific CD8+ T cell proliferation was increased in both cases by blocking PD-1, similar to previously reported findings (5).
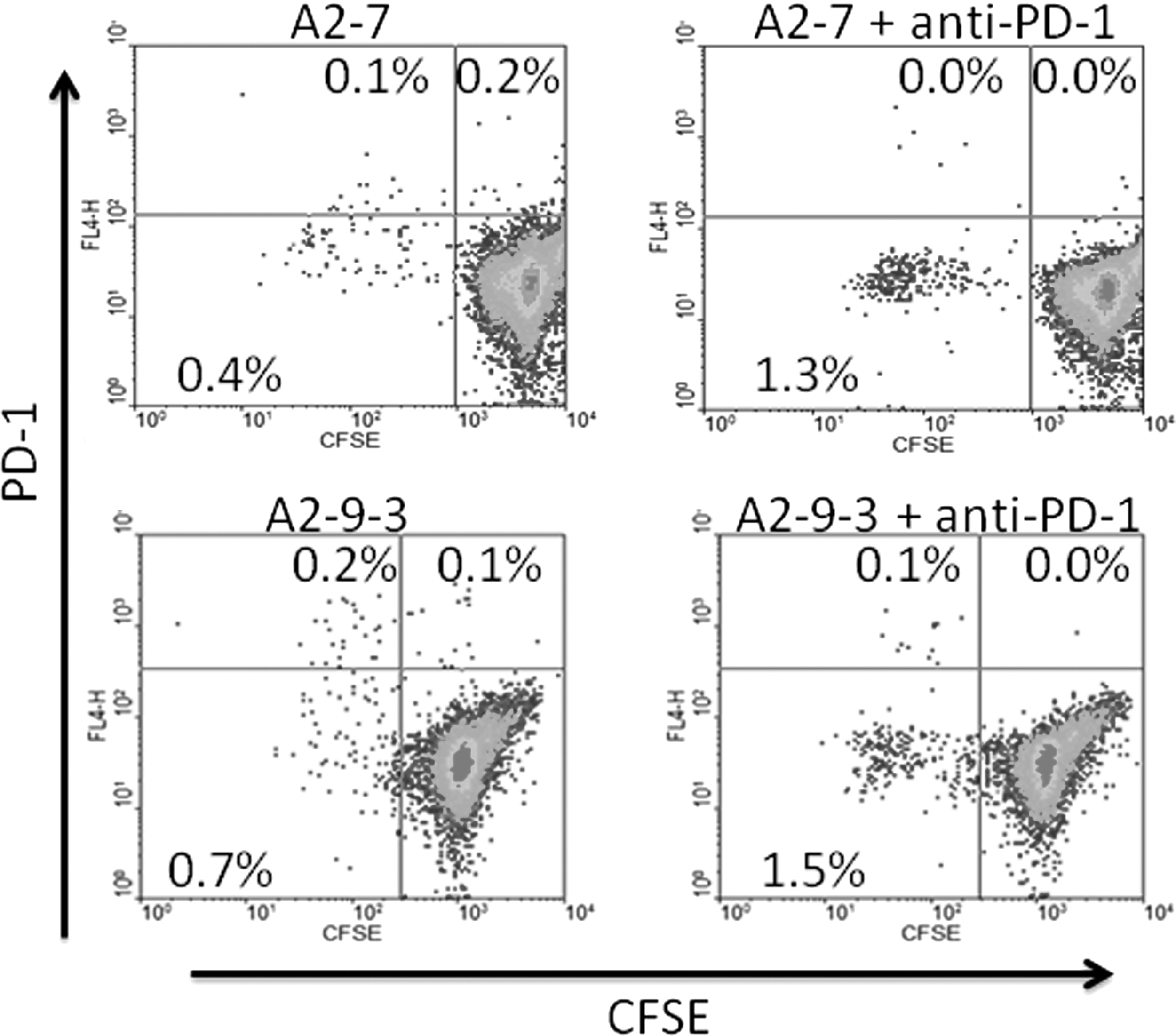
Effect of blocking PD-1 on proliferation of HIV-specific CD8+ T cells stimulated with reference or heteroclitic peptides. After 7 days of stimulation with peptides±anti-PD-1 as listed above the plot frames, CD8+ cells were gated for analysis as in Figure 1 and proliferating cells identified by plotting CFSE intensity versus PD-1 expression. Cells to the left of the vertical quadrant lines have undergone proliferation. PD-1 expression was not assessed in the cultures to which anti-PD-1 was added. In the examples shown, blocking PD-1 increased HIV-specific CD8+ T cell proliferation stimulated by reference peptide A2-7 and heteroclitic peptide A2-9-3 by 160% and 78%, respectively. Numbers in the plot quadrants represent the percentage of total gated cells in that quadrant.
The median percentage of proliferating CD8+ T cells expressing PD-1 following stimulation with reference HIV peptides was significantly higher (p=0.005, Wilcoxon signed-rank test) at 29% with interquartile range (IQR) 19.5–79.5% versus 25%, IQR 17.5–64.5% than following stimulation with heteroclitic peptides (Fig. 3A). The difference in PD-1 expression was small when compared across the entire 29 cases (29% vs. 25%), but there were a number of instances within the 13 cases of lower PD-1 expression where the effect was quite large (Table 3).
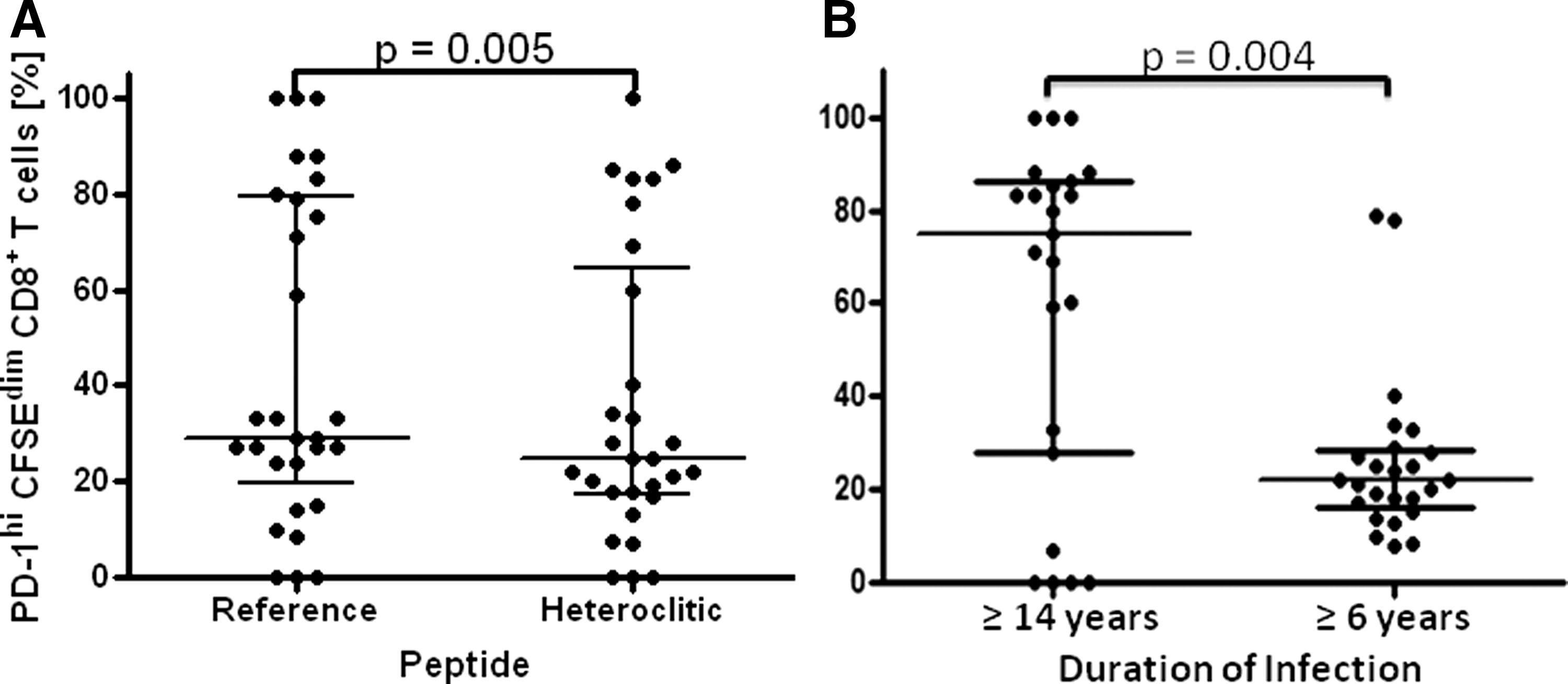
There appeared to be a more bimodal (>60% and <40% clusters) than normal distribution of percentages of proliferating CD8+ HIV-specific T cells that expressed PD-1 following stimulation with either reference or heteroclitic peptides. While many factors can affect PD-1 expression, the duration of infection recorded for responders to heteroclitic peptides also had a bimodal distribution around ≥6 and ≥17 years (Table 4). Therefore, we also compared median percentage of proliferating CD8+ T cells expressing PD-1 following stimulation with either heteroclitic or native peptides between the groups distinguished by duration of infection. The median percentage of proliferating CD8+ T cells expressing PD-1 was significantly higher in the group with a longer duration of infection (p=0.004, Mann–Whitney test) at 75%, IQR 28.0–86.0% versus 22%, IQR 16.0–28.5% (Fig. 3B).
Virus load obtained from clinical charts for the time of testing expressed as log10 copies HIV RNA/mL plasma.
Number of CD4+ and CD8+ T cells/μL peripheral blood at the time of testing.
CD4+ T cell nadir is the lowest recorded CD4+ T cell count from clinical charts.
Duration of infection is based on the earliest record of HIV-seropositive status.
Whether subjects were receiving HAART at time of testing.
HAART, highly active antiretroviral therapy.
Heteroclitic peptides did not increase proliferation of HIV-specific CD8+ T cells and reduce PD-1 expression in the same 13 cases (Table 3) and there was no significant correlation between enhancement of proliferation and reduction in PD-1 expression on CD8+ T cells stimulated with heteroclitic peptides (Fig. 4). However, stimulation of HIV-specific CD8+ T cells with heteroclitic peptides enhanced proliferation and reduced PD-1 expression in a total of 5/29 cases and either reduced PD-1 expression or enhanced proliferation in a total of 21/29 cases (Table 3). Thus, in the majority of cases, heteroclitic peptides identified by their ability to augment IFN-γ or IL-2 production by HIV-specific CD8+ T cells also had positive effects on either proliferation or differentiation of HIV-specific CD8+ T cells.
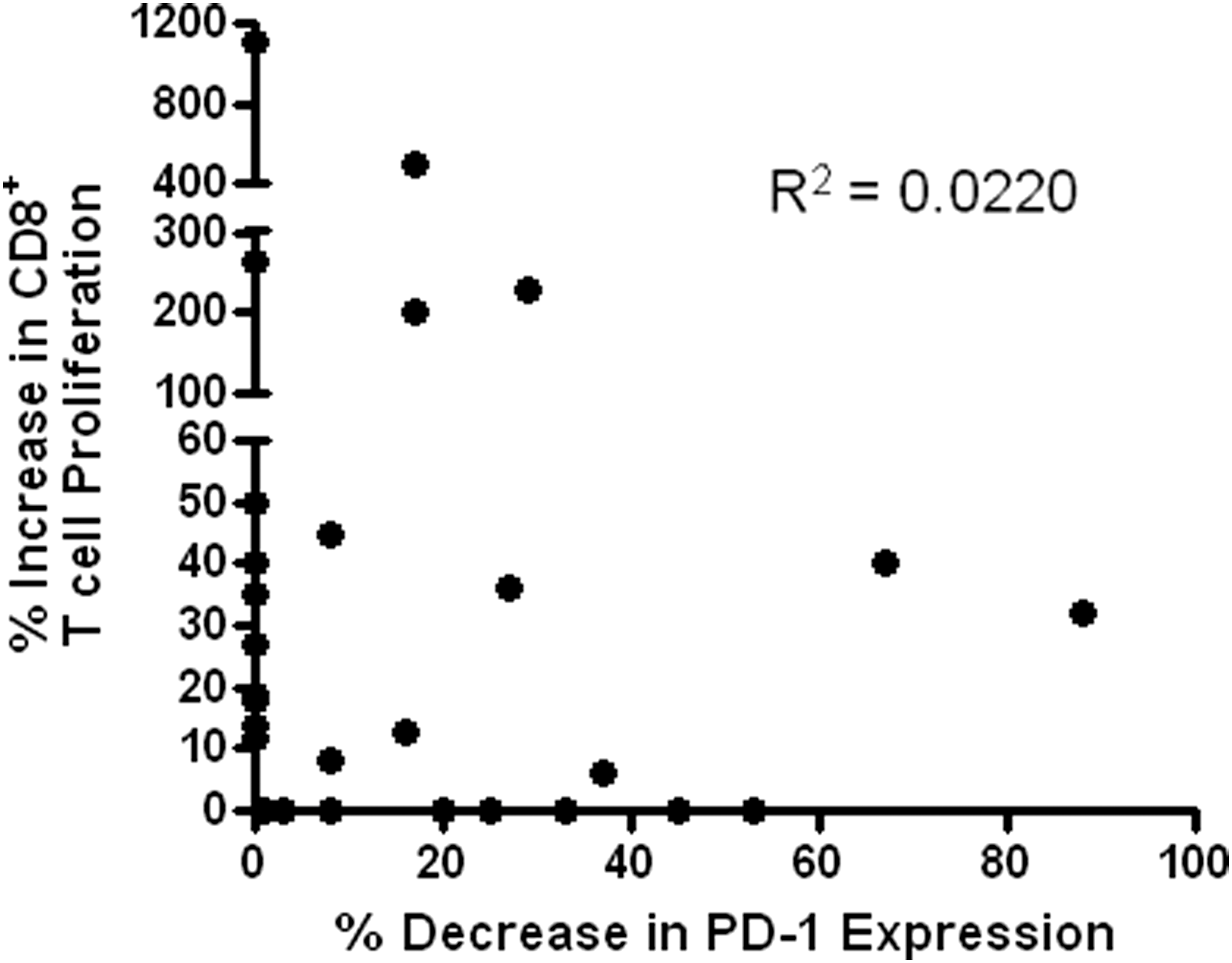
Scatter plot of increased proliferation versus reduced expression of PD-1 stimulated by heteroclitic peptides. Linear regression analysis indicated no significant correlation between the two effects as shown on the graph.
Discussion
The failure of HIV-specific CD8+ T cells to durably control viral replication reflects the combined effects of viral escape mechanisms and cellular immune dysfunction. The loss of T cell polyfunctionality, in terms of the range of cytokines produced and reduced capacity for proliferation, is closely associated with chronic infection, disease progression, and development of a dysfunctional or exhausted T cell phenotype. Depletion of CD4+ T cells, less overall IL-2 production and degradation of other forms of help may be partially responsible for reduced proliferation of HIV-specific CD8+ T cells in progressive HIV infection (1). However, some HIV-specific CD8+ T cells produce IL-2 themselves and can thereby drive their own proliferation (30).
Since we previously showed that heteroclitic peptide stimulation can enhance both IFN-γ and IL-2 production by HIV-specific CD8+ T cells, we followed up in this study by testing whether heteroclitic peptides that increase cytokine production also enhance HIV-specific CD8+ T cell proliferation and reduce the dysfunctional phenotype of responding cells.
The heteroclitic peptides used in this study were initially identified by their ability to augment IFN-γ or IL-2 production during overnight stimulation of PBMC in ELIspot assays. Proliferation of HIV-specific CD8+ T cells and their expression of markers associated with functional exhaustion on the proliferating cells were then compared following extended (7 day) in vitro stimulation of PBMC with heteroclitic versus reference peptides. Of the 29 cases where heteroclitic peptides enhanced either IFN-γ or IL-2 production, proliferation was increased in 13 cases and was roughly equivalent to that induced by the reference peptide in another 7 cases. In the same 29 cases, expression of the T cell exhaustion marker PD-1 was also reduced on proliferating HIV-specific CD8+ T cells in 13 cases. However, these were not the same 13 cases in which proliferation was increased and there was no significant correlation between increased proliferation and reduction in PD-1 expression.
Since T cell activation itself induces PD-1 expression, it is plausible that some peptides that increased CD8+ T cell proliferation might also result in a higher percentage of responder cells expressing PD-1. Conversely, our results and those of others suggest that the absence of PD-1 expression on proliferating cells enables more proliferation (5). In any case, we saw no significant positive or negative correlation between reduction in PD-1 expression and increased proliferation of HIV-specific CD8+ T cells stimulated by heteroclitic peptides.
Ideally, activation, proliferation, and evolution of the exhausted phenotype for CD8+ T cells stimulated by heteroclitic versus reference peptides should be monitored across multiple time points within the in vitro stimulation process, but cell numbers available for this study were limiting. Likewise, it would be informative to comparatively phenotype the circulating reference and heteroclitic peptide-reactive cells using tetramers or intracellular cytokine production, but the frequency of cells responding against individual peptides ex vivo is generally too low for accurate phenotyping by flow cytometry without an initial sorting step. This approach would potentially be applicable where exceptionally large responses are detected by ELIspot or through analysis of very large numbers of PBMC obtained by leukophoresis.
There were five cases in which heteroclitic peptides increased proliferation and reduced PD-1 expression by HIV-specific CD8+ T cells. These cases best illustrate the potential for heteroclitic peptides to affect differentiation of HIV-specific CD8+ T cells toward the polyfunctional status persistently exhibited by HIV-specific CD8+ T cells in the small minority of HIV-infected, ART-naive individuals referred to as elite controllers or long-term nonprogressors (29).
Other approaches to restoring or improving functionally impaired T cell responses have involved selectively blocking T cell inhibitory signaling pathways and/or providing exogenous costimulation (5,23,25). While we showed in this study that heteroclitic HIV peptides improved the character of HIV-specific CD8+ T cell responses in the absence of exogenous costimulatory molecules and without blocking inhibitory signaling pathways, the mechanism by which the heteroclitic peptides act is not clear. Our results suggest that heteroclitic peptides either selectively stimulate nonexhausted T cells with specificities related to the reference peptide or restore a subset of apparently exhausted HIV-specific CD8+ T cells to a functional or even polyfunctional state. If the heteroclitic peptides stimulate a different subset of CD8+ T cells than the reference peptides, we would expect reference and heteroclitic peptides together to produce an additive response in ELIspot assays. However, when we previously tested this possibility, adding reference and heteroclitic peptides together did not increase responses above those stimulated by the heteroclitic peptide alone (10).
The contextual variability in the heteroclitic activity suggests that multiple factors relate to the ability of a particular variant peptide to stimulate a superior response in a particular individual and that more than one mechanism may be involved. For example, the available T cell repertoire will be partially dictated by the entire complement of HLA molecules, not just the one actually presenting the peptide. In addition, individual histories of antigenic stimulation and disease progression will have differential effects on capacities of both the antigen-presenting and T cells of study subjects. This limits the ability to generalize the heteroclitic peptide activity for any particular peptide across individuals sharing the same HLA-class I antigen and, thus, identifying heteroclitic peptides universally or even generally active across a particular HLA type may not be a realistically achievable goal. However, developing a better understanding of what features favor the heteroclitic peptide activity in different settings through identifying and studying more cases should help guide work at the individual level to produce personalized heteroclitic peptides or peptide sets.
Rather than acting on a different set of T cells, activation signals generated by heteroclitic peptide stimulation could somehow bypass the PD-1 and other inhibitory signaling pathways to enhance proliferation and cytokine production even by those HIV-specific CD8+ T cells with phenotypic signs of dysfunction. Inhibitory signals delivered by PD-1 normally downregulate TCR signaling through direct dephosphorylation of intracellular signaling intermediates. The phosphatases (SHP-1 and SHP-2) associated with PD-1 dephosphorylate CD3ζ and prevent the phosphorylation of ZAP-70 and PKCθ (12). One possible explanation for enhanced proliferation of PD-1 expressing T cells in response to heteroclitic peptide stimulation, which has previously been reported, could be increased tyrosine phosphorylation of ZAP-70 and TCRζ chains in CTL stimulated with heteroclitic peptides relative to CTL stimulated with native peptides (19).
The different variables recently reported to affect levels of PD-1 expression on HIV-specific CD8+ T cells suggest that relatively subtle features associated with TCR signaling strength and integration of TCR signaling with that through modulatory pathways can impact significantly on clonotypic expansion and progression toward an exhausted phenotype (14). Therefore, the overall impact on TCR avidity of aa variations affecting HLA binding, TCR interactions, or both may fine tune T cell signaling strength across thresholds determining differential activation of those transcription factors around which cytokine production patterns pivot (9).
Surprisingly, in a number of cases, we observed enhanced proliferation of HIV-specific CD8+ T cells in response to heteroclitic peptides without detectable IL-2 production. Although IL-2 is a T cell growth factor, it can also increase expression of PD-1. Culture of purified T cells with common γ-chain cytokines such as IL-2, IL-7, IL-15, and IL-21 markedly enhances PD-1 expression in vitro and this effect was confirmed in vivo when higher levels of PD-1 expression were demonstrated on PBMC isolated from HIV-infected individuals receiving IL-2 immunotherapy (13). In cases where peptides increased proliferation without detectable IL-2 production or drove proliferation of cells expressing less PD-1, this could reflect very low levels of autocrine IL-2 driving proliferation resulting in less PD-1 expression.
This study provides further evidence of potential benefits to incorporating heteroclitic HIV peptides into therapeutic vaccines aimed at improving HIV-specific CD8+ T cell responses. Clarification of whether distinct, partially, or wholly overlapping sets of T cells are stimulated by heteroclitic and reference peptides will require comparative TCR analysis of responding populations. A better understanding of how the aa substitutions make heteroclitic peptides more antigenic in the context of different TCRs could help in designing more active heteroclitic variants with optimal substitutions at multiple positions. It is important to note that heteroclitic peptides employed in this study were variants of immunodominant epitopes and that there may also be many lesser epitopes with heteroclitic variants that can stimulate superior responses.
We have also only investigated the effect of heteroclitic peptides on PD-1 expression, while many other inhibitory molecules, including TIM-3, LAG-3, CD160, CTLA4, 2B4, and SLAM, are implicated in T cell dysfunction and exhaustion. Further investigation should also determine whether heteroclitic peptides modulate the expression of other inhibitory molecules through their effects on T cell proliferation and differentiation and if this is a factor in the enhanced T cell responses generated by heteroclitic peptides. Given the need for novel methods of enhancing HIV-specific CD8+ T cell characteristics for control of HIV replication, viral activation, and eradication strategies, it will be important to further investigate the potential benefits of heteroclitic HIV peptides (6,21).
Footnotes
Acknowledgments
We thank all the blood donors who participated in this study. This study was supported by the Memorial University Faculty of Medicine and a research operating grant (HOP-111725) from the Canadian Institutes for Health Research (CIHR) awarded to Michael Grant.
Author Disclosure Statement
No competing financial interests exist.