
Abstract
Regulatory T cells (Tregs) play a pivotal role in suppressing specific antiviral immune responses during the progression of chronic hepatitis B virus infection (CHB) as well as tumorigenesis. Programmed death-1 ligand-1 (PD-L1) expressed on Tregs can transduce an inhibitory signal into effector T cells through interacting with programmed death-1 (PD-1). However, in CHB patients, the clinical significance of PD-L1 expression on Tregs has not been clearly described. This study investigated the frequency of circulating Tregs and PD-L1 expression on Tregs and analyzed their correlations with clinical parameters. The data show that both the frequency of CD4+CD25+FoxP3+ Tregs and PD-L1 expression on Tregs in the peripheral blood increased significantly in CHB patients when compared with healthy controls. At the same time, it is shown that PD-L1 expression on Tregs was positively correlated with the percentage of Tregs in CHB patients. Moreover, the results demonstrated that both Treg frequency and PD-L1 expression on Tregs positively correlated with the levels of alanine aminotransaminase (ALT) and aspartate aminotransferase (AST), both of which are indicators of the extent of liver injury. Taken together, these findings suggest that PD-L1 on Tregs might contribute to progression of hepatitis B virus infection through mediating the inhibitory function of Tregs. Thereby, blockade of interaction between Treg-expressing PD-L1 and PD-1 on effector T cells may be adopted as a potential therapeutic approach in CHB.
Introduction
C
Of all the coinhibitory molecules (also called checkpoint molecules), cytotoxic T lymphocyte-associated antigen-4 (CTLA-4), programmed death-1 (PD-1), T cell immunoglobulin and mucin domain-3 (TIM-3), and B and T lymphocyte attenuator (BTLA) were found to correlate closely with the exhaustion of CD8+ T cells during progression of infectious diseases and tumorigenesis (2,6,10,11,16 –18,27). PD-1, based on interacting with its ligands PD-L (PD-L1 or PD-L2), can transmit a negative signal intracellularly, leading to inhibition of T cell activation and proliferation (19). Studies by the authors and others have previously described that PD-1 expression obviously increased on CD4+ and CD8+ T cells of patients with HBV infection and was positively correlated with viral replication and liver injury (23,28,36,38). Because blockade of PD-1/PD-L inhibitory signal remarkably restores T cell function and enhances antitumor immune responses, current antitumor immunotherapies with anti-PD-1 or anti-PD-L1 antibodies have achieved great clinical success (29,34). With regard to infectious diseases, experimental data revealed that blocking PD-1 signal could also improve the antiviral function of CD8+ T cells (4,22).
Tregs, characteristically expressing foxhead box P3 (Foxp3) in the nucleus, play a vital important role in suppressing T cell activation, expansion, and effector function. Mounting data have demonstrated that an increase in Treg frequency can attenuate HBV-specific CTLA immune responses, resulting in failure to eradicate viruses and persistent replication of HBV in the body (1,13). By releasing inhibitory cytokines such as IL-10, TGF-β, and IL-35 and competitively binding with IL-2 through high expression of CD25 on the surface, Tregs play a role in suppressing activation of effector T cells (5,8). Moreover, Tregs can also exert their inhibitory function by employing the negative coinhibitory ligands through cell–cell-dependent contact between Tregs and effector T cells. For example, PD-L1 and HVEM, upon binding to PD-1 or BTLA expressed by effector T cells, can drive the suppressive functions of Tregs (15,20,33,35). Further, blockade of the interaction between Treg-expressing PD-L1 and PD-1 on effector CD8+ T cells could rescue CD8+ T cell function, and adoptive transferred wild-type but not HVEM−/− mouse-deriving Tregs were able to control alloresponses in vivo by normal effector T cells (33,35).
However, the expression pattern of PD-L1 on Tregs of CHB patients and its clinical significance have not been well investigated. The current study examined PD-L1 expression on circulating CD4+CD25+FoxP3+Tregs, as well as the frequency of Tregs from CHB patients, and analyzed their correlations with serum levels of HBV DNA load, aspartate aminotransferase (AST), and alanine aminotransaminase (ALT). This work is valuable for exploring the function and mechanism of Tregs on CHB progression further, and provides a potential indicator for evaluating CHB development.
Materials and Methods
Study subjects
Forty-nine CHB patients and 40 healthy donors (HC) were enrolled for this study. Fresh heparinized whole blood was collected from these subjects for investigation. The diagnoses of CHB patients were complied with The Diagnostic Criteria of Diagnostic Criteria for Viral Hepatitis B issued by The Ministry of Health of the People's Republic of China. Briefly, CHB patients presented a clinical course of hepatitis B virus infection for 6 months, with HBeAg being positive or detection of HBV DNA, and they may have had abnormal hepatic function. Patients were excluded for co-infection with hepatitis C virus (HCV), hepatitis D virus (HDV), human immunodeficiency virus (HIV), acute hepatitis, and other causes of liver disease. Healthy donors were identified without HBV, HCV, HDV, and HIV-1 infection or other known diseases. The study was conducted at the Affiliated Infectious Hospital of Soochow University and approved by the ethical committee of the hospital. Written informed consent was obtained from each subject.
Serological assays for HBV markers, viral load, and other basic clinical features
HBV serological markers, including hepatitis B surface antigen (HBsAg), hepatitis B surface antibody (HBsAb), hepatitis B e antigen (HBeAg), and anti-hepatitis B e antibody (HBeAb), were measured by using assay kits (Abbott Ireland, Sligo, Ireland). The HBV viral load was determined using a HBV LC PCR Kit (Shanghai Shenyou Technology Co. Ltd., Shangahi, China). At the same time, the routine biochemical assays, including liver function, albumin (ALB), total bilirubin (TILB), and so on were also assayed.
Flow cytometric analysis
For determining CD4+CD25+FoxP3+ population and PD-L1 expression, mouse antihuman CD4-FITC, CD25-PE/Cy7, Foxp3-Alexa Flour 647, and PD-L1-PE were purchased from BD Pharmingen (San Diego, CA). For staining surface markers and Foxp3, a Human Regulatory T Cell Whole Blood Staining Kit purchased from eBioscience (San Diego, CA) was adopted according to the standard procedure. Briefly, for each test, 100 μL of fresh heparinized whole blood from CHB patients or healthy donors was incubated with mouse antihuman CD4-FITC, CD25-PE/Cy7, as well as PD-1-PE or mouse IgG2a (10 μL) for 15 min, then lysed with RBC lysis buffer, subsequently dealt with fixation/permeabilization solution after washing and centrifuging. Further mouse antihuman Foxp3-Alexa Flour 647 or mouse IgG1-Alexa Flour 647 was added and incubated at 4°C for 30 min in the dark. Eventually, cells were suspended in fetal bovine serum, 4′,6-diamidino-2-phenylindole (DAPI) was added and determined by BD FACSAria (San Jose, CA) supported by DB FACSDiva software (San Jose, CA). Data were analyzed using FlowJo (San Carlos, CA).
Statistical analysis
The data were analyzed with GraphPad Prism v6.0 software (San Diego, CA) and are shown as means ± standard error of the mean (SEM). Nonparametric tests (Mann–Whitney U-test) were performed, and two-tailed p-values of < 0.05 were considered statistically significant between the compared groups. A nonparametric correlation test (Spearman) was applied to analyze the correlation between Tregs frequency or PD-L1 expression and virus load, ALT, or AST levels. Each bar presented in the figures represents the mean ± SEM.
Results
Investigation of frequency of CD4 + CD25 + FoxP3 + Tregs of CHB patients
The recruited participants include CHB patients (n = 49) and HC (n = 40). Table 1 demonstrates the demographic and clinical characteristics of study populations, including age, sex, HBeAg sero-status, serum HBV DNA concentrations, serum levels of AST and ALT, serum total protein, albumin, and total bilirubin.
Data are presented as number or mean ± standard error of the mean.
CHB, chronic hepatitis B virus infection; HC, healthy controls; HBeAg, hepatitis B e antigen; HBV-DNA, hepatitis B virus-DNA; ALT, alanine aminotransaminase; AST, aspartate aminotransferase; ALB, albumin; TBIL, total bilirubin; NA, not applicable.
Tregs have been described as contributing to the suppression of anti-HBV immune responses and the development of CHB (13,24,37). This study investigated further the percentage of the circulating CD4+CD25+FoxP3+population from subgroups of CHB patients with different levels of ALT, AST, or HBV DNA load. Gating strategies for the CD4+CD25+FoxP3+population are shown in Figure 1A. Similar to previous reports (24,37), the percentage of Tregs in the peripheral blood of CHB patients was clearly elevated compared with the HC (Fig. 1B). Interestingly, when patients were divided into subgroups according to the levels of clinical indicators such as serum HBV DNA levels, ALT, and AST, the percentage of Tregs presented a dynamic change compared with HC (Fig. 1C–E). The percentages of Tregs from CHB patients with low and high serum HBV load (<107 and ≥107 DNA copies, respectively) were both significantly higher than that of HC, but there was no significant difference between the two subgroups (Fig. 1C). The results further showed that the percentage of Tregs in CHB patients with low levels of serum ALT (<75 IU/L) or AST (<60 IU/L) was correspondingly lower than those with high levels of ALT (≥75 IU/L) or AST (≥60 IU/L), respectively (Fig. 1D and E). However, there was no significant difference between HC and CHB patients with low levels of serum ALT (<75 IU/L) or AST (<60 IU/L; Fig. 1D and E).

The frequency of CD4+CD25+Foxp3+regulatory T cells (Tregs) in circulating CD4+ T cells of chronic hepatitis B (CHB) patients and healthy controls (HC).
Examination of PD-L1 expression on CD4+CD25+FoxP3+ Tregs of CHB patients
Since PD-L1 expressed on Tregs can mediate a coinhibitory signal toward effector T cells and thereby takes part in the mechanisms of Treg inhibitory function (15,20,35), this study examined PD-L1 expression pattern on Tregs of CHB patients from subgroups of CHB patients with different levels of clinical parameters such as ALT, AST, and HBV load.
Gating strategies for determination of PD-L1 on CD4+CD25+FoxP3+ population are shown in Figure 2A. Surprisingly, PD-L1 expression on circulating CD4+CD25+FoxP3+ Tregs of CHB patients profoundly increased compared with that of HC (Fig. 2B). Further analysis demonstrated that similar to the Treg percentage, PD-L1 expression on Tregs of CHB patients with low and high serum HBV load (<107 and ≥107 DNA copies, respectively) were also higher than that of HC, but no difference between the two subgroups was found (Fig. 2C). With regard to the serum levels of AST and ALT, PD-L1 expression on Tregs presented variations with similarities to the Treg percentage. That is, PD-L1 expression on Tregs of CHB patients with low levels of serum ALT (<75 IU/L) or AST (<60 IU/L) was accordingly lower than those with high levels of ALT (≥75 IU/L) or AST (≥60 IU/L), respectively (Fig. 2D and E). Moreover, no differences were found between HC and CHB patients with low levels of serum ALT (<75 IU/L) or AST (<60 IU/L; Fig. 2D and E).
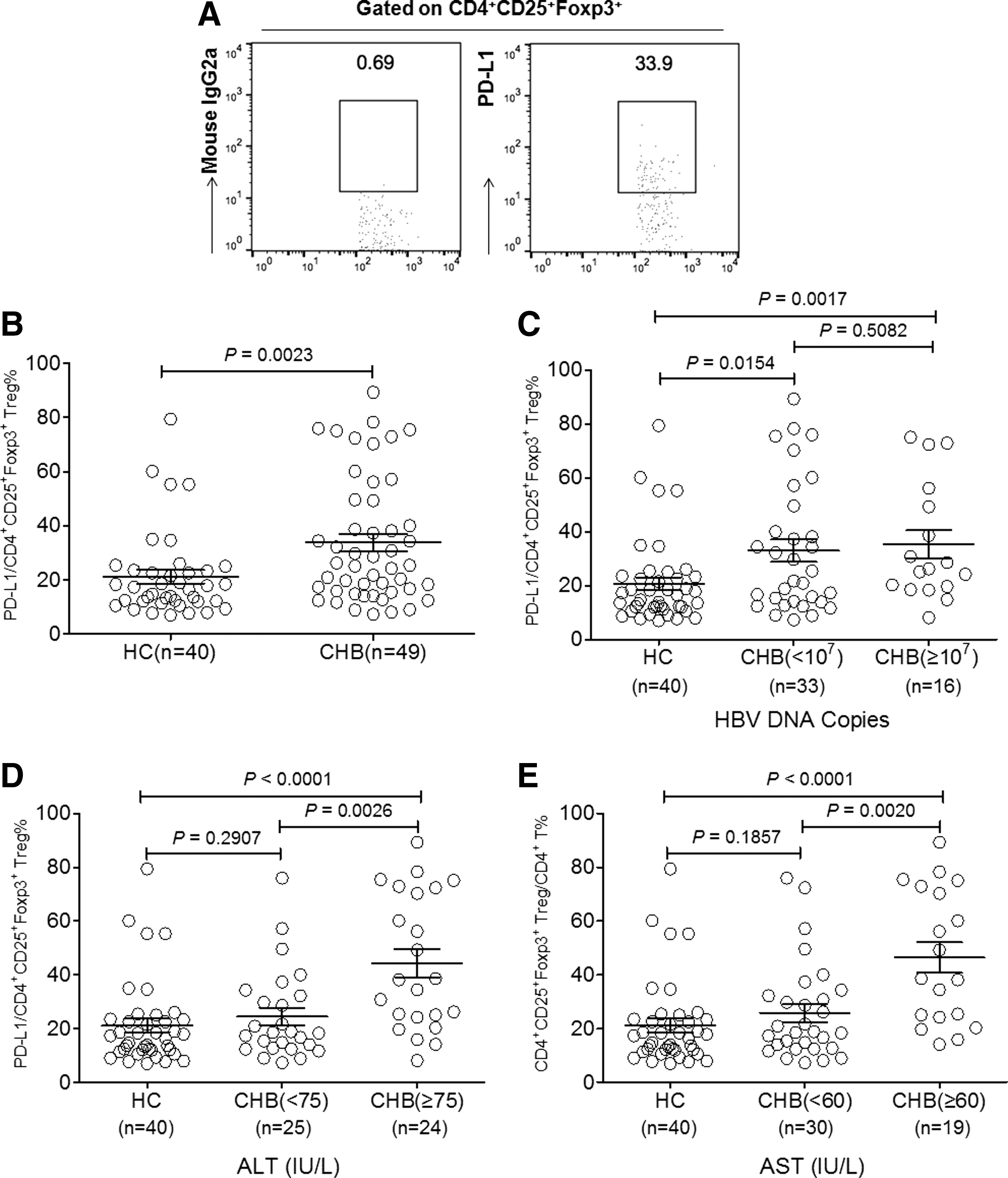
The expression profiles of programmed death-1 ligand-1 (PD-L1) on circulating CD4+CD25+Foxp3+regulatory T cells (Tregs) of CHB patients and HC.
Analysis of correlations of the Treg percentage and PD-L1 expression on Tregs with clinical parameters
With both percentages of Treg and expression levels of PD-L1 on Tregs differing in subgroups of CHB patients with different levels of ALT or AST, their correlations with such clinical parameters as well as HBV DNA load were analyzed. First, it was concluded that both the percentage of Treg and the level of PD-L1 expression on Tregs were not correlated with the load of HBV DNA (Fig. 3A and B). Second, it was found that percentages of Treg and the level of Treg-expressing PD-L1 were positively correlated to ALT and AST levels (Fig. 3C–F). In addition, the levels of PD-L1 expression on Tregs positively correlated with the percentage of Tregs (Fig. 3G).
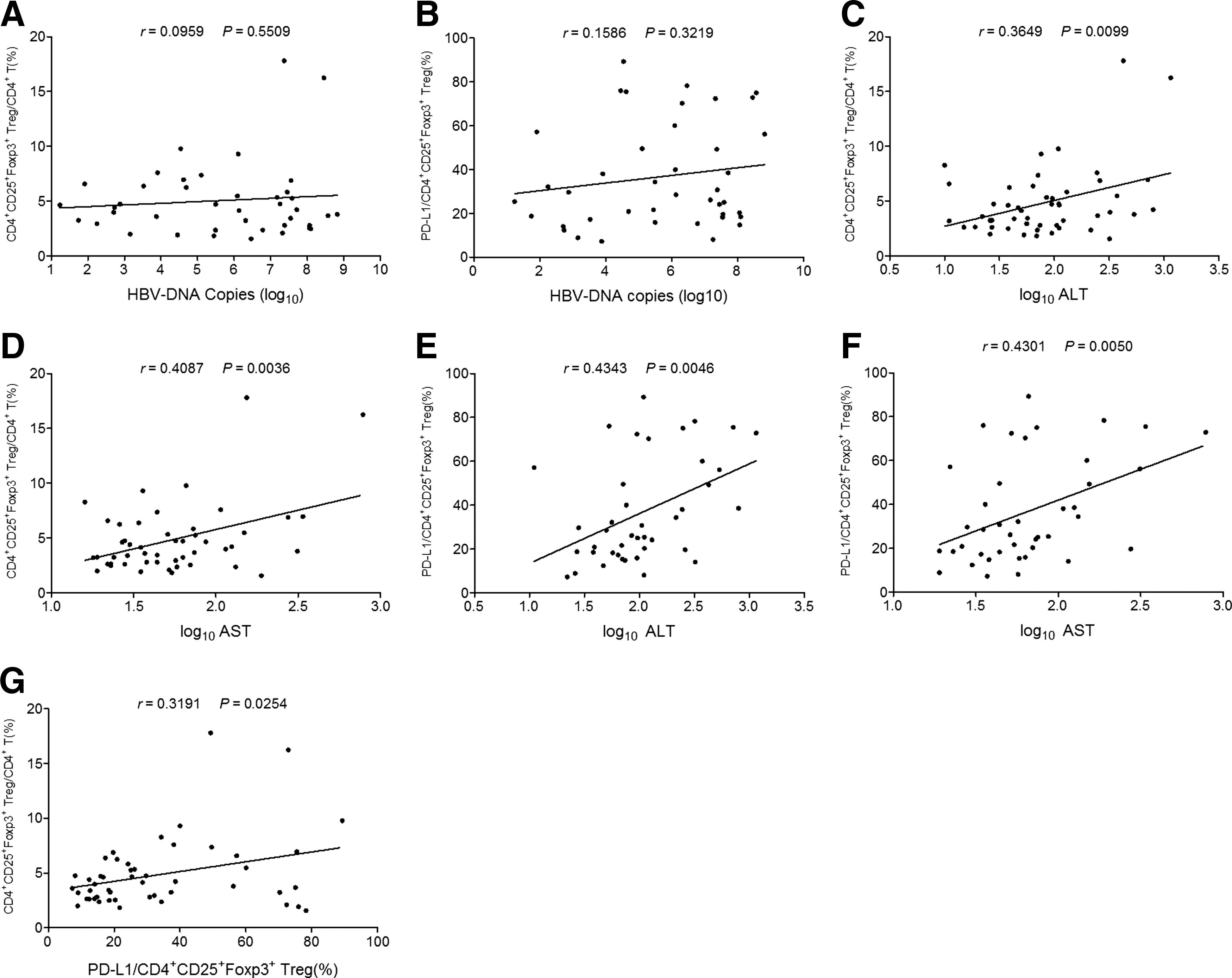
The correlations of Treg frequency and the levels of PD-L1 expression on Tregs with clinical parameters of CHB patients.
Discussion
In the progression of CHB, due to persistent replication, HBV continually infects hepatocytes, induces nonspecific immune responses, and causes intermittent liver damage, which is closely associated with hepatocirrhosis and hepatic carcinoma. In recent years, mounting data have explored the mechanisms involved in immune evasion of HBV and the causes contributing to chronic infection. Of these, elevation of Treg frequency and upregulation of immune checkpoints on CD8+ T cells have been identified to contribute greatly to restraining and exhaustion of CD8+ T cells, the backbone of the immune system exerting pressure on anti-HBV infection (9,11,24,25,37).
As a type of regulatory feedback mechanism, the frequency of Tregs increases, along with priming and sustaining of immune responses against pathogens for preventing the body from immunopathological damage (3). However, aberrant elevation of Treg frequency participated in restraining effective anti-HBV immune responses and resulted in persistent HBV replication. The present study showed that the frequency of CD4+CD25+FoxP3+ Tregs of CHB patients is clearly higher than that of HC, in agreement with previous reports (24,37). Further analysis revealed that the frequency of Tregs correlated with the levels of AST and ALT, indicating that during CHB progression, the immunopathological damage could lead to elevated inflammation, which might have resulted in increases in Tregs. As a result, increased Tregs can further inhibit anti-HBV immune responses and favor HBV evasion. In contrast to liver damage, the present study showed no obvious correlation between Treg frequency and HBV DNA load, which was consistent with some earlier studies (12,32), but differed from other reports (24,37). The discrepancies might be due to samples used in different studies having variable levels of HBV DNA load.
Treg-mediated suppression relies on cell–cell-dependent contact. Currently, it has been revealed that Treg-expressing PD-L1 and HVEM, based on binding to PD-1 or BTLA, can trigger inhibitory signals into effector T cells (15,33). Although PD-1 has been identified as being upregulated on the surface of CD4+ and CD8+ T cells of CHB patients (28,38), the expression patterns of PD-L1 on Tregs and its clinical significance in CHB have not been depicted. The present study revealed that PD-L1 expression on CD4+CD25+FoxP3+ Tregs was obviously upregulated when compared with the HC. Then it was identified that the level of PD-L1 expression on Tregs was significantly correlated to the percentage of CD4+CD25+FoxP3+ in CD4+ subsets with a positive correlation. These results suggest that in accordance with elevation of Treg frequency and upregulation of PD-1 on T cells, PD-L1 could increase its expression on Tregs so as to help achieve a full inhibitory effect on T cell activation. Further, it was also found that PD-L1 expression was positively correlated with the levels of ALT and AST, which concurred with variation of Treg frequency, indicating that inflammation mediated by liver injury might not only induce Treg increase, but also drive PD-L1 upregulation on Tregs. Hence, Treg-expressing PD-L1 as well as Treg frequency could be considered as a potential clinical indicator to reflect CHB progression. Further, blocking PD-L1/PD-1 axis between Tregs and effector T cells may be adopted as a potential therapeutic approach in CHB.
Conclusion
In summary, this study first described PD-L1 expression profiles on CD4+CD25+FoxP3+ Tregs of CHB patients and the correlations with clinical parameters. PD-L1 expression on circulating Tregs of CHB patients was obviously higher than that of HC and changed dynamically, associated with liver damage and Treg expansion, indicating that the expression of PD-L1 on Tregs may contribute to HBV infection progression through mediating the inhibitory function of Tregs.
Footnotes
Acknowledgments
We appreciate the participation of the volunteers in this study. The research was supported by “973” grant of China (2013CB530501) and the Foundation of Science and Technology Project of Suzhou City (SYS201101, SS201246).
Author Disclosure Statement
No conflicting financial interests exist.