
Abstract
Today, hepatitis C virus (HCV) infection is considered as one of the most significant international health concerns. Although novel therapeutic regimens against the infection have shown satisfactory results, no approved vaccine exists yet. This study aimed to evaluate the immunogenicity of a DNA vaccine candidate for HCV-3a, based on nonstructural proteins NS3/NS4A, in C57BL/6 mice. Immunogenicity effect of pDisplay-NS3/NS4A was analyzed through immunization with 100 and 200 μg concentrations of the construct with complete Freund's adjuvant, monophosphoryl lipid A (MPL), or without adjuvant. The frequencies of different splenic mononuclear cells were measured using the Mouse Th1/Th2/Th17 Phenotyping Kit. Moreover, the number of T-CD8+ cells was determined using conjugated anti-CD8a and anti-CD3e antibodies by flow cytometry. As observed, the frequencies of Th1, T-CD8+, and Th2 cells increased in all the experimental groups, compared with the controls. The highest levels of the respective cells were seen in the group immunized with 200 μg of the construct with MPL. Also, there were positive correlations between the frequency of Th1 cells and those of Th2 and T-CD8+ cells in all the immunized groups, but were significant in those receiving adjuvants. The frequency of Th17 cells did not statistically change among the groups. Taken together, our findings revealed that the constructed DNA vaccine encoding HCV-3a NS3/NS4A gene induces the cell-mediated immune responses significantly. However, its coadministration with adjuvants exhibits more efficient results than the recombinant plasmid alone. Further study is currently underway to evaluate the specific immune responses and recognize the responsible antigenic epitopes.
Introduction
T
HCV belongs to Flaviviridae family with the estimated RNA genome size of 9.4 kb (33), which encodes a polyprotein of 3010–3011 amino acids, cleaved into different structural and nonstructural proteins (18). Given the extreme level of genome virus mutations (31), universal HCV vaccine development is faced with major challenges (12) due to the lack of genome conserved regions with adequate immunogenic epitopes. Thus, the attempts to construct a specific vaccine of the genotype in each region could be helpful.
Among the HCV viral proteins, nonstructural ones, especially NS3, have been recognized as one the central targets of interest for antiviral discovery, as well as new vaccine development, owing to multiple important epitopes with the ability to induce potent immune responses (17). Utilizing the NS4A protein fused to NS3 protein showed the enhancement in the stability, expression, and immunogenicity of the NS3 protein (10,36). Because of the significant role of cell-mediated immune responses and in particular cytotoxic T-cell immunity in the elimination of the infection (6), investigations into the vaccines that induce this type of immunity are sped up, with more emphasis on nonstructural proteins. Besides, development of the structural protein-based vaccines to stimulate humoral immunity with the production of neutralizing antibodies is also increased for the prevention of the disease (19). However, currently, just some of these attempts have reached the clinical trial phases, and no approved vaccine exists yet (37). In addition, most of such studies are mainly focused on genotype 1 of the virus, while based on the results of the recent successful therapeutic protocol, genotype 3 is known as the most difficult to treat (14,38). Moreover, according to the previous epidemiological studies, this genotype is the second most prevalent one worldwide (20). Thus, further investigations on this genotype seem to be critical.
Recently, applying HCV DNA vaccines because of their safety and potency to elicit both humoral and cellular immune responses in hosts has been extended and serves as a promising means for the production of an effective generation of vaccines (34). Activating T-helper and cytotoxic T-cell and antibody responses in animal models using such vaccines for genotype 1 of the virus (32) could raise hopes to develop the vaccine for other genotypes as well. To our knowledge, by far, limited research has been carried out on designing DNA vaccines for genotype 3. Hence, the present study was conducted to evaluate the immunogenicity of a novel DNA vaccine candidate for HCV genotype 3a, based on nonstructural proteins NS3/NS4A, in mice.
Materials and Methods
Generation of HCV genotype 3a NS3/NS4A expression plasmid
Construction and in vitro expression of the plasmid containing all sequences of NS3 and NS4A genes (pDisplay-NS3/NS4A), isolated from the plasma of a HCV-3a-infected patient, were previously described (2). It was shown that these target proteins were expressed as a membrane-anchored peptide in Huh7 cells due to some engineered parameters in the plasmid related to the secretory pathway and are responsible for anchoring the fusion protein to the plasma membrane (the murine Ig k chain leader sequence and platelet-derived growth factor receptor transmembrane domain (PDGFR-TM).
Mice immunization
Forty female C57BL/6 mice (6–8-week old) were purchased from the Center of Comparative and Experimental Medicine of Shiraz University of Medical Sciences, Shiraz, Iran. The mice were divided into 10 groups (6 experimental, 2 control, and 2 treated with adjuvant alone) and housed in accordance with the Animal Care Committee of the Shiraz University of Medical Sciences guidelines. The six experimental groups were immunized with different concentrations of pDisplay-NS3/NS4A (100 or 200 μg) in sterile PBS with 50 μL per mouse of complete Freund's adjuvant (CFA; Sigma, Saint Louis, MO), 50 μg per mouse of monophosphoryl lipid A (MPL; Sigma), or without adjuvant. The two control groups were sensitized either with the empty expression vector pDisplay or PBS. Moreover, the other two groups were treated with MPL or CFA with the above-mentioned doses. Each animal received three injections, 2 weeks apart, at a total volume of 200 μL divided equally into two portions and injected into quadriceps femoris muscles of both sides of each mouse (Fig. 1a). All the injections were carried out using insulin syringes with a 29-gauge needle attached.
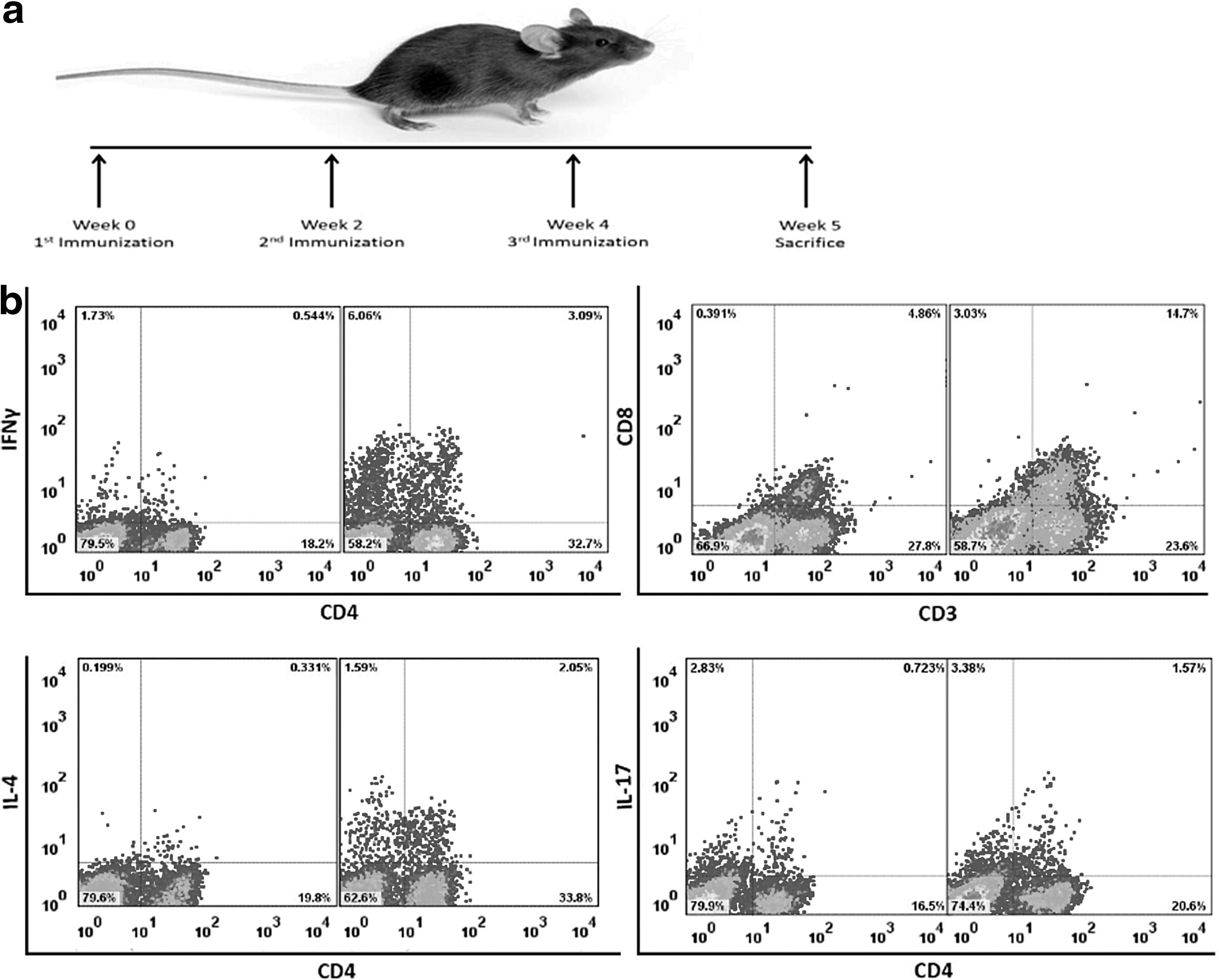
Immunization scheme and typical flow cytometry dot plots.
Mononuclear cell isolation
One week after the last injection, the animals were sacrificed, the spleens were separated, crushed, filtered, and the mononuclear cells were isolated by the Lymphodex (Inno-Train, Kronberg im Taunus, Germany) gradient. Briefly, after 20 min of centrifugation at 400 g, the middle layers containing the cells were collected, washed twice, and suspended in the RPMI-1640 medium supplemented with 10% fetal bovine serum, 1% glutamine, 100 IU/mL penicillin, 100 μg/mL streptomycin, and 2% HEPES. Then, the cell numbers and viability were determined using a hemocytometer by trypan blue staining.
Cell stimulation
To determine the different types of T cells in each sample, 2 × 106 cells per 1 mL of supplemented RPMI-1640 medium were initially mixed with GolgiStop™ (4 μL for every 6 mL of cell culture; BD Pharmingen, San Diego, CA), containing monensin (an intracellular protein transport inhibitor), to prevent the cytokine release from the cells and then stimulated with 50 ng/mL phorbol myristate acetate (Sigma) and 1 μg/mL calcium ionomycin (Sigma) for 5 h. Thereafter, the cells were harvested and fixed with the Cytofix™ buffer (BD Pharmingen) and permeabilized with the Perm/Wash™ buffer (BD Pharmingen). Cell staining was performed using the Mouse Th1/Th2/Th17 Phenotyping Kit (BD Pharmingen) containing anti-CD4, anti-IL-17A, anti-IFN-γ, and anti-IL-4 antibodies conjugated with PerCp, PE, FITC, and APC (BD Pharmingen), respectively. In addition, FITC anti-CD8a (BD Pharmingen) and PE anti-CD3e (BD Pharmingen) antibodies were used to detect T-CD8+ cells. The stained cells were processed using a four-color flow cytometer instrument (FACSCalibur; BDBiosciences, San Jose, CA). The frequency of Th1, Th2, and Th17 was determined among the gated CD4+ T cells expressing IFN-γ, IL4, and IL-17, respectively. In addition, the percentages of CD8+ T cells were measured throughout the gated CD3+ cells. The data were analyzed by FlowJo software, version 7.6.1 (Tree Star, Ashland, OR) (Fig. 1b).
Statistical analyses
The frequency of Th1, T-CD8+, Th2, and Th17 cells and the ratio of CD4/CD8, as well as Th1/Th2 cells in different groups, are reported as mean ± standard deviation. The data were compared between the groups using the independent-samples t-test. Moreover, the correlations of the Th1 cell frequencies with those of Th2 and T-CD8+ cells, and Th2 cell frequencies with those of T-CD8+ in the groups injected with recombinant plasmid with and without adjuvants were determined by linear correlation analysis. The statistical analyses were performed by SPSS for Windows (version 16; SPSS, Inc., Chicago, IL) and the data were considered statistically significant at a two-sided p < 0.05. All the comparison graphs were generated using GraphPad Prism, version 5.0 (GraphPad Software, Inc., La Jolla, CA).
Results
Induction of T-helper subpopulation in experimental and control groups
Following the last intramuscular immunization with the construct, mononuclear cells were isolated from the spleen of each animal and the frequency of T lymphocytes was determined. The results showed that the frequencies of Th1, T-CD8+, and Th2 cells significantly increased in the groups receiving the construct either with or without adjuvants (MPL or CFA), in comparison with the control groups (Tables 1 –3). The highest rate of increase was observed in the group immunized with 200 μg of the construct with MPL (Fig. 2a–c). Although the frequency of Th17 cells increased in the experimental groups, and more in those that received adjuvants besides construct, no statistically remarkable changes were observed among the groups (Fig. 2f and Table 4).
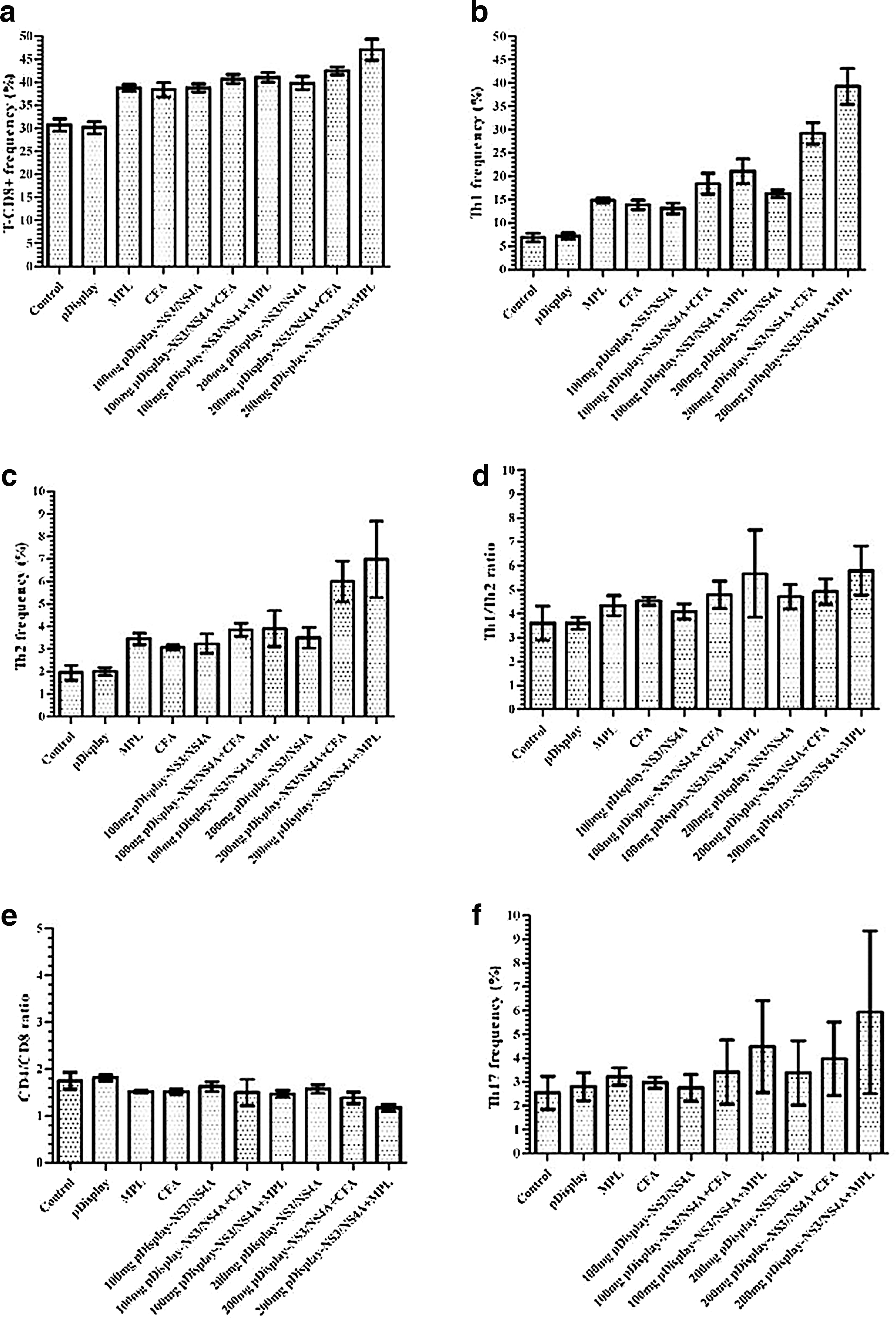
The frequency of different immunity cells and their ratios in different groups. An increase was observed in the frequency of T-CD8+
CFA, complete Freund's adjuvant; MPL, monophosphoryl lipid A; SD, standard deviation.
Th1/Th2 and CD4/CD8 ratios among different immunized groups and controls
The comparison of the Th1/Th2 ratio among different experimental and control groups revealed a significant increase in all experimental ones, except in the group that received 100 μg of the construct with MPL (Fig. 2d and Table 5). Furthermore, the ratio of CD4/CD8 showed a significant decrease among the experimental groups immunized with the construct plus adjuvants except the group that received 100 μg of the construct with CFA, compared to those in the controls (Fig. 2e and Table 6).
Correlations among different T-helper subpopulations in the groups immunized with the construct with or without adjuvant
When the correlations of the Th1 cell frequencies with those of Th2 and T-CD8+ cells, and Th2 cell frequencies with those of T-CD8+ in the groups that received adjutants besides the recombinant construct were calculated, positive correlations were observed with significant p-values (Fig. 3a–c). Although the p-values were not statistically significant for the groups immunized with the construct without adjuvant, positive correlations with r 2 = 0.373 and r 2 = 0.42 were seen for Th1 with T-CD8+ and Th2, respectively, and r 2 = 0.18 for Th2 with T-CD8+ (Fig. 3d–f).
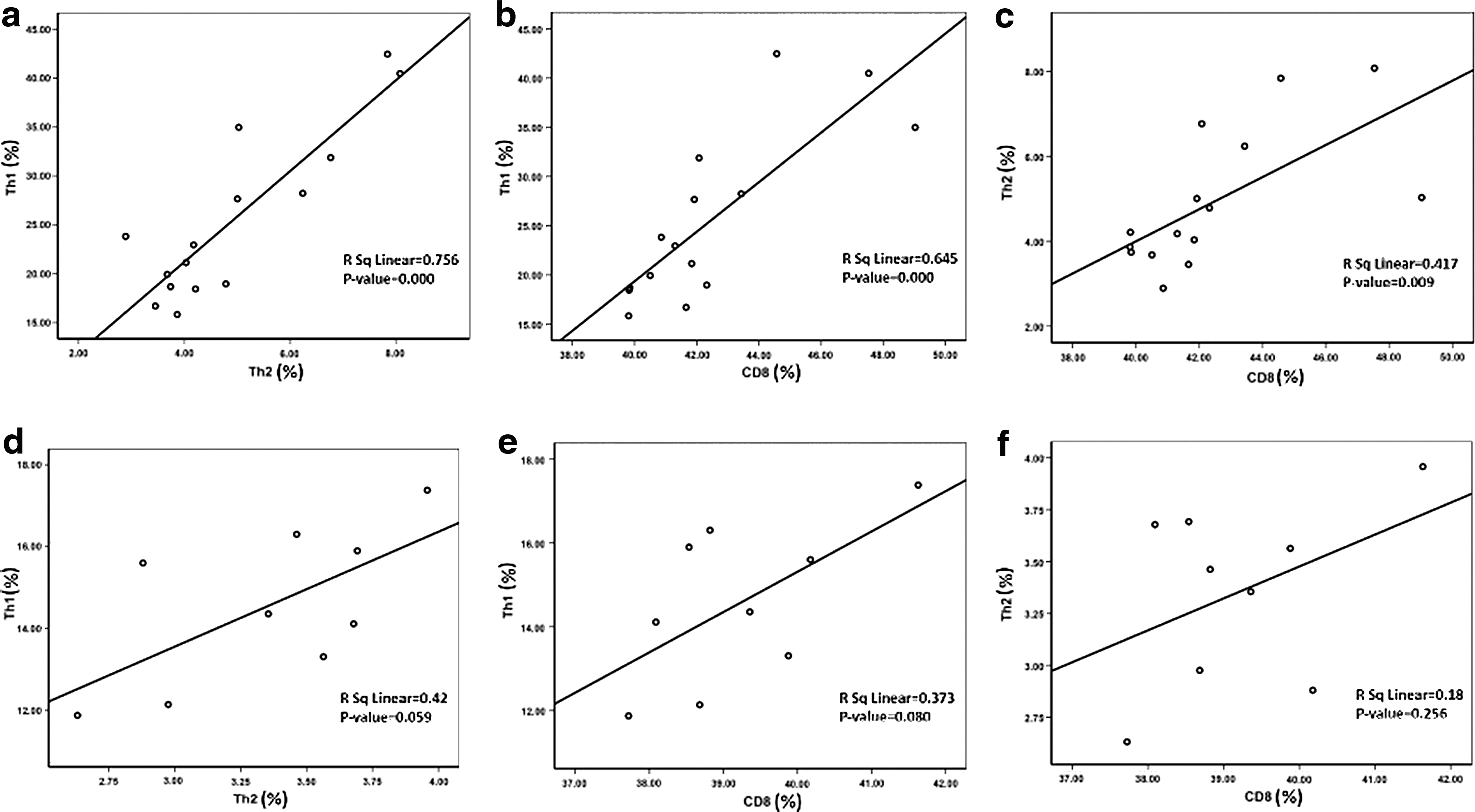
The correlation analysis between the frequencies of different cells in the different immunized groups. The graphs exhibit significant positive correlations between the frequency of Th1 and Th2
Discussion
In this study, we showed that the cell-mediated immune response was elicited in mice, following immunization with mammalian expression plasmid encoding the NS3/NS4A proteins of HCV genotype 3a. Nowadays, DNA vaccine strategy has been recognized as one of the most reliable, safe, and also highly potential methods for the development of vaccines against different pathogens, including HCV (34). Moreover, the high resemblance between the antigenic epitopes of the target proteins expressed by DNA vaccine plasmids and the native viral proteins (32) makes it more attractive for researchers. HCV vaccine strategies particularly focused on eliciting T-cell response are somehow related to these attempts. Ratnoglik et al. reported the induction of cell-mediated immune responses in mice using a DNA vaccine expressing an altered NS3 protein of HCV genotype 1b (26). Strong cellular immune responses were also observed in both mice and Rhesus macaques after immunization by HCV genotype 1a/1b consensus DNA vaccine encoding the NS3/NS4A proteins (17).
In the current study, the frequencies of T-CD8+ cells were statistically elevated in all the vaccinated mice, but the group immunized with higher concentration of the recombinant plasmid with MPL showed a notable increase than the others. Meanwhile, the ratio of CD4/CD8 showed a significant decrease among the experimental groups than those in the control ones implying the elevation of CD8+ T-cell frequency. The association between HCV clearance and CD8+ T cells, in the context of MHC class-I presentation, was previously determined (34). The studies on the NS3-based DNA HCV therapeutic vaccines also revealed the important role of this protein in producing CD8+ T-cell responses and resolution of the infection (24,26,34). Regarding the pivotal function of CD8+ T cells in response to viral infections (35), in our study, a higher frequency of CD8+ T cells in the presence of adjuvants may result in more viral clearance activity in the infected cases.
Similar to CD8+ T-cell frequencies, our results showed a significant increase in the frequencies of Th1 cells in the immunized groups, compared with those in the controls. Earlier studies reported that activating cellular immune responses, including Th1 and cytotoxic T cells at the first frontier, could be a promising means for the prevention or even elimination of the HCV infection in animal models (16,17,27). As revealed previously, an increase in the production of IFN-γ by Th1 cells promotes more expression of both MHC class-I and class-II on antigen-presenting cells, as well as cytotoxic immunity, which gives rise to stronger antiviral effects (11,29). Therefore, the elevation in the frequency of Th1 in response to pDisplay-NS3/NS4A may result in higher production of IFN-γ, leading to enhanced antiviral activity of the immune cells.
Moreover, at the next level of protection against HCV infection, antibody production induced by Th2 cells against different HCV proteins is assumed to be effective, especially in the prevention of the infection by neutralization of the virus. In this study, we also found that Th2 cells exhibited a significant increase in all the vaccinated groups. Previous studies revealed that antibodies in response to recombinant E1 and/or E2 proteins in vaccinated chimpanzees have been shown to be partially protective against the infection (21). Another study, using a mixture of HCV Core, E1, E2, and NS3, demonstrated the control of viremia by neutralizing antibodies in challenged mice with the virus (19). Furthermore, the important functions of both Th1 and Th2 cells in increasing primary CD8+ T-cell responses and the establishment of long-lived CD8 memory were documented (7). Thus, it seems that the construct used in our study may be effective against the infection by producing Th1, Th2, and activating as well as inducing the long-lived CD8+ memory T cells.
Although the levels of Th1 and Th2 cells increased significantly in all the vaccinated groups, when the ratio of Th1/Th2 was analyzed, the higher increase was observed for Th1. This may cause more production of IFN-γ, which can promote the higher rate of cellular immune responses, including Th1 and cytotoxic T cells.
The results of the correlations analysis of the Th1 frequencies with those of Th2 and T-CD8+ in this study, revealed a notable positive role of adjuvant in increasing the r 2 significantly. Although the positive correlation of the Th2 frequencies with those of CD8+ T cells is not as important as those for Th1 and CD8+ T cells for promoting cytotoxic immunity, this could help increase the lifetime of memory CD8+ T cells (7) and induction of specific antibody by B cells.
Besides, we demonstrated the insignificant changes in the Th17 cell frequency, as one of the factors involved in the inflammatory phenomenon, across all the groups. It was shown that the cytokine IL-17 produced by Th17 cells plays as a proinflammatory and profibrotic mediator (22). Moreover, the role of this cytokine in several inflammatory autoimmune diseases, as well as in models of tissue injury, was reported (15). Thus, our results may imply that the adjuvant induces its positive role with no significant adverse effects of inflammation.
Applying adjuvants in combination with DNA vaccines to induce stronger and long-lasting immune responses, as well as shifting it from a Thl cell-mediated to a Th2 humoral immune response or vice versa, was investigated in previous studies (28). Earlier studies revealed the function of CFA in slower releasing of antigens and altering their trafficking to the immune system stimulating both cell-mediated and humoral immunity, but more Th1 responses (4). In addition, MPL has been shown to deviate the Th0 cells into Th1 more than Th2 (8,25). Having compared the two adjuvants, we observed that MPL is more efficient than CFA. Because of the ability of MPL as an adjuvant for incorporation into human vaccines (5), the use of such an adjuvant is recommended in combination with the regular HCV DNA vaccines, for better stimulation of the acquired immunity.
In conclusion, given the increased levels of Th1, T-CD8+, Th2 cells in the experimental groups, compared to those in controls, the elevated specific immune response can be suggested. We also found that coadministration of the constructed DNA vaccine encoding the HCV-3a NS3/NS4A gene in combination with the adjuvant is a more potent inducer of the cell-mediated immune responses, compared with the recombinant plasmid alone. Meanwhile, a further study is currently underway to evaluate the specific immune responses and recognize the responsible antigenic epitopes.
Footnotes
Acknowledgments
Our special thanks to Hassan Khajehei, PhD, for copyediting the article. This research was financially supported by PhD thesis No. 93-7033 of Shiraz University of Medical Sciences and Professor Alborzi Clinical Microbiology Research Center.
Author Disclosure Statement
No competing financial interests exist.