
Abstract
Cervical cancer (CC) is one of the main causes of death among women of reproductive age. Although there are different tests, the disease tends to be diagnosed at late stages. In recent years, the use of complementary tests or sequential diagnostic tests has been implemented. Nevertheless, the results are variable and not conclusive; therefore, more studies for improving the usefulness of these tests in diagnostics are necessary. The human papillomavirus (HPV) infection has been associated with both benign and malignant proliferation of skin and mucosal tissues. Furthermore, some HPV types have been classified as high risk due to their potential to cause cancer, and HPV16 is most frequently associated with this disease. Although between 70% and 80% of precancerous lesions are eliminated by the host's immune system, there is no available test to distinguish between regressive lesions from those that could progress to CC. An HPV infection generates a humoral immune response against L1 and L2 capsid proteins, which can be protective and a response against early proteins. The latter is not a protective response, but these antibodies can be used as markers to determine the stage of the infection and/or the stage of the cervical lesion. Up to now, the humoral immune response resulting from the HPV infection has been used to study the biology of the virus and the efficacy of the HPV vaccines. Although there are no conclusive results regarding the use of these antibodies for diagnosis, we hereby review the actual panorama of the antibody response against the HPV proteins during the development of the disease as well as their possible use as biomarkers for the progression of cervical lesions and of CC.
Introduction
H
During the past years, serological studies have proven to be important in understanding the natural history of HPV infections. More recently, sero-epidemiological studies have focused on confirming the association between the presence of HPV antibodies and the detection of anogenital cancers and their precursors, whereas serological studies evaluate the effectiveness of the HPV vaccine. HPV serologic assays have used either L1 virus-like particles (VLPs) or E6/E7 oncoproteins, and other nonstructural HPV proteins have been used less frequently. An anti-HPV16 E6/E7 response has not only been predominantly found in patients with advanced CC but it has also been found in control patients. This has made the use of anti-HPV antibodies as CC biomarkers controversial. However, there are promising studies that combine different viral antigens as new biomarkers for the early detection and prevention of HPV-associated CC (27,94,138).
Structure and Viral Cycle of the HPV
HPV is a small nonenveloped icosahedral virus of ∼50 nm in diameter belonging to the Papillomaviridae family. This virus has a double-stranded circular DNA genome of around 8 kb. The viral genome is divided into three regions: the long control region (LCR) that acts as regulator of viral DNA transcription and replication; the early region (E1, E2, E4, E5, E6, and E7 genes) involved in viral replication and carcinogenesis, and the late region (L1 and L2 genes) that contributes to viral capsid production (12,78,85,114,145).
The life cycle of HPV requires differentiation of the host epithelial cells and this process is divided into two phases: productive and nonproductive. The nonproductive stage takes place at the proliferating basal epithelial cells where the virus is maintained in an episomal form at low copy number (50–100 copies/cell) (73,78). The productive phase is carried out at the suprabasal differentiating cell layer and is characterized by the amplification of the viral DNA, the expression of the L1 and L2 genes, and the virus production (40).
The initial infection involves access of viral particles to the basal cell layer made possible by a breaching in the stratified epithelium. It has been suggested that viral entry to the host cells is performed through clathrin or caveolin-coated vesicles and it requires the presence of heparan sulfate proteoglycans and laminin-322 as receptors (22,30,42,56). At the same time, structural changes in the viral capsid are necessary, including the furin cleavage of the L2 minor capsid protein that facilitates interaction with a secondary receptor on the surface of the basal cell layer, which is essential for virus internalization and viral genome transport to the nucleus through the L2-viral DNA complex (100,112).
Since virus production is accomplished during cell differentiation, HPV replication starts with the division of the cells in the cervical stratified epithelium, where the virus is internalized. The transcription of the E6 and E7 genes starts through host cell factors that interact with the LCR viral promoter. This promoter regulates the expression of viral proteins and allows the interaction of the E1 protein with the replication origin (9,77). Papillomavirus proteins, E1 and E2, form a heterodimer complex to control viral replication, while E2 protein regulates the expression of the early viral genes, E6 and E7 (86,130).
The sequential expression of the viral proteins starts when major viral E7 oncoprotein, binds tumor suppressor protein, pRb, allowing the cell to continue within the cell cycle in the absence of mitogenic signals (86). Almost at the same time, E6 protein is expressed and it recruits the cellular ubiquitin ligase E6AP to the E6-substrate complex, which in turn results in substrate polyubiquitination and degradation at the proteasome of p53, a transcriptional factor for cell cycle arrest and, in extreme situations, for induction of apoptosis (111). Later on, E5 protein is necessary to maintain cell proliferation since it increases the epidermal growth factor receptor signaling pathway through the retardation of receptor degradation and the inhibition of the H+ endosomal adenylpyrophosphatase; it is suggested that E5 complements the functions of oncoproteins, E6 and E7 (29).
Almost at the end of the viral cycle, the E4 protein is expressed at high levels in cells that are supporting the viral genome amplification and it is able to bind the cytoskeleton and promote its collapse to finally release viral particles (32,33,105).
At the end of the viral cycle, the late (L) region of the HPV genome (3 kb in length) expresses the two viral capsid proteins L1 and L2 that form the icosahedral capsid at the upper layers of the infected tissue once the viral genome amplification has ended (35,68,91,130). The viral particles comprise the viral genome within a capsid, which contains 360 copies of L1 protein and 12 copies of L2 protein, all arranged in an icosahedral structure of 72 pentameric capsomeres (82,130).
Due to the fact that HPV proteins perform their functions within the cell and the viral antigens are poorly exposed to the host's immune system, the antibody response against these antigens is low. However, continuous stimulation of the immune system throughout the viral cycle induces levels of specific antibodies suitable for their study as possible biomarkers. Nevertheless, the usefulness of this immune response against HPV is unclear and requires further investigation.
Immune Response to the HPV Infection
The uterine cervix mucosa, which is the site of infection by HPV like other mucosal tissues, has a high threshold of immune tolerance (17). It is well known that during the acute phase of the infection, a controlled inflammatory environment leads to the elimination of the virus. However, when this does not occur, the immune microenvironment becomes more tolerant, allowing the infection to become persistent and eventually induce cancer.
High-risk HPV types 16 and 18 mainly induce persistent infections without frequent serious complications for the host; they are also highly successful in releasing viral particles transmissible to others (76). This virus takes the host to a point of balance where the infection does not represent a serious drawback and viral replication is not limited by the host's immune response (119) because the virus does not have a blood-borne phase or viremia. The HPV infection does not induce necrosis, cytolysis, or inflammation, and as a result, there is little or no release of proinflammatory cytokines in the local environment (59). The infection is exclusively intraepithelial, and replication and assembly occur in cells already destined for death by anoikis (detachment). Consequently, practically, there are no danger signals to alert the immune system to generate an efficient response to eliminate the infection (102,120).
However, more than 80% of the genital lesions caused by HPV infections are cleared as a result of a successful cell-mediated immune response, during which cells of the innate immune system such as keratinocytes, dendritic cells (DCs), Langerhans cells (LC), macrophages, natural killer (NK), and natural killer T cells may play an important role in clearing the infection by promoting a proinflammatory process (109).
The keratinocytes found in the female genital track are the natural hosts of the HPV infection and they can act as immune sentinels (87). These cells express toll-like receptors (TLRs, belonging to the pathogen recognition receptor [PRR] family) on the cell surface (TLR-1, TLR-2, TLR-4, TLR-5, and TLR-6) and in the endosomes (TLR-3 and TLR-9). In particular, TLR-9 is activated by unmethylated double-stranded CpG-rich DNA (8), leading to the secretion of cytokines such as type 1 interferons (IFNs) that activate the NK cells (43), which in turn kill the HPV-infected cells (61,128).
In contrast, once the expression of HPV antigens is established, the innate immune response is downmodulated, thus facilitating the evasion from the immune system. In this sense, E6 and E7 oncoproteins, downregulate the IFN response by interfering with different molecules involved in the signal transduction pathway.
The E6 oncoprotein can bind the transcriptional factor IRF-3, blocking the synthesis of IFN; it also blocks Tyk2 to downregulate the signaling of IFN-α through Jak-STAT (58,62,92,101). Additionally, the E7 oncoprotein inhibits the activation of the IFN-β promoter by binding to IRF-1 and it deregulates the expression of TLR-9 in the cervical epithelium by repressing its promoter (24,46,47,66,122). Finally, due to the downregulation of IFNs by the HPV oncoproteins, there is a reduction of the NK cell cytotoxic activity against the HPV-infected cells (20,38,62,109). All together, these events allow the establishment of a persistent HPV infection with concomitant progression to CC.
On the other hand, the LCs and DCs (antigen-presenting cells) connect the innate and the adaptive immune response, through antigen-specific presentation to B and T cells in the lymph node, and they initiate the adaptive immune response to eliminate the HPV infection. In this process, the Th cells secrete proinflammatory cytokines that create a Th1-type microenvironment, helping activate Tc cells to kill the infected cells. These T-cell responses are mainly directed to the early HPV proteins, E2, E6, and E7 (142). At the same time, this immunological response is accompanied by the generation of neutralizing antibodies against the L1 capsid protein to further inhibit the propagation of the viral infection (39,119). However, the virus has developed several strategies to further evade this adaptive immune response.
Recent evidences suggests that E2, E6, and E7 interact with the promoters of the immunoregulatory cytokines, IL-10 and TGF-β, to upregulate their expression (122,131). In addition, the E5 oncoprotein also has an impact on the adaptive immune response since it sequesters the major histocompatibility complex (MHC-I) in the Golgi apparatus, thus preventing the transportation of the MHC-I complex to the cell membrane and hence affecting the antigen presentation to the Tc cells (6,95,125). On the other hand, E5 also interferes with the antigen presentation to the Th cells since this protein prevents the acidification of the endosomes, where the MHC-II-restricted antigen is processed (146). As a result, an imbalance between Th1 (proinflammatory) and Th2 (anti-inflammatory) cytokines is produced and the immune system fails to clear the HPV infection, which results in the establishment of a persistent infection with a high risk for the development of CC (65,137).
Finally, an adaptive immune response through the activation of B cells is necessary for the humoral immune response. In this way, the HPV proteins (including the early proteins) generated throughout the infection somehow reach the draining lymph nodes, where the HPV antigens are recognized by the B-cell receptor, present in the membrane of the B-cells, which receive a stimulus by Th cells through CD40 and the release of cytokines (120). In this way, B cells are differentiated into plasma cells that produce antibodies against the different HPV proteins. It is in this sense that differential antibody response against E4, E6, E7, and L1 has been detected in the sera of women infected with HPV (121), and specifically anti-E7 antibodies have been identified in HPV-associated CC (54,55,80,89,94,107,138).
Humoral Immune Response Against HPV Antigens
The specific antibody response against HPV antigens has been investigated for several decades, either as a method to study the viral cycle or as a marker of HPV-associated cancers at different anatomical sites (99,136). In this regard, early studies showed that seroconversion and the presence of anti-HPV antibodies were associated with the existence of cervical lesions and CC. More recently, it has been established that HPV is an important etiological factor in the case of head and neck cancer (4,10,70,106,118).
Several epidemiological studies have investigated the presence of HPV antibodies in groups or populations with different exposures to the virus, finding great diversity of the prevalence and kinetics of these antibodies, depending on the population type, the antigens used, and the detection method (Tables 1 and 2). For this purpose, different procedures have been used to identify the serological antibodies against HPV antigens, including ELISA (enzyme-link immunosorbent assay), Western blot, and more recently, Luminex multiplex. These methodologies use different antigens such as VLPs from different HPV types; synthetic peptides from HPV16 proteins L1, L2, E2, E4, and E7, or the HPV16 bacterial recombinant E4 and E7 proteins. Additionally, these technologies have been optimized and modified to increase the sensitivity and specificity of the assays, finding that Western blot and Luminex multiplex are the ones that display the best performance (45).
Underlined percentages correspond to lesions grouped as CIN1–2 or CIN1–3 according to reference articles.
Abs, antibodies; CC, cervical cancer; CIN, cervical intraepithelial neoplasia; ELISA, enzyme-linked immunosorbent assay; GST, glutathione S-transferase; HPV, human papillomavirus; HPV+, HPV positive; ND, not done; RIPA, radioimmunoprecipitation assay.
Underlined percentages correspond to lesions grouped as CIN1–2 or CIN1–3 according to reference articles.
VLPs, virus-like particles.
Table 1 summarizes a large number of epidemiological and clinical studies that have searched for the presence of antibodies against HPV early proteins and for associations of these serological markers with premalignant uterine cervical lesions and/or CC. In this sense, antibodies against E6 and E7 proteins, are the most explored and most frequently associated with CC (10,99,107) or with precancerous lesions (83,138). However, anti-E7 antibodies are more commonly associated with CC than anti-E6 antibodies (99).
Until now, the results reported have been contradictory as some groups have found a similar prevalence of anti-E7 antibodies in controls and in CC cases (10–25%) (41), while others have shown a higher prevalence of anti-E7 antibodies in CC (30–75%) than in controls (3–10%). These differences can be attributed to the different methodology performances and the type of antigens used (Table 1) (1,18,37,55,64,72,80,93,94,97,99,124,129). Furthermore, other groups have shown that anti-E7 antibodies can also be associated with an earlier stage of the disease such as the cervical intraepithelial neoplasia (CIN)3 lesions (∼75%) (Fig. 1), which point to a more significant prevalence of these antibodies in CC than in precancerous lesions (Table 1) (10,72,138).
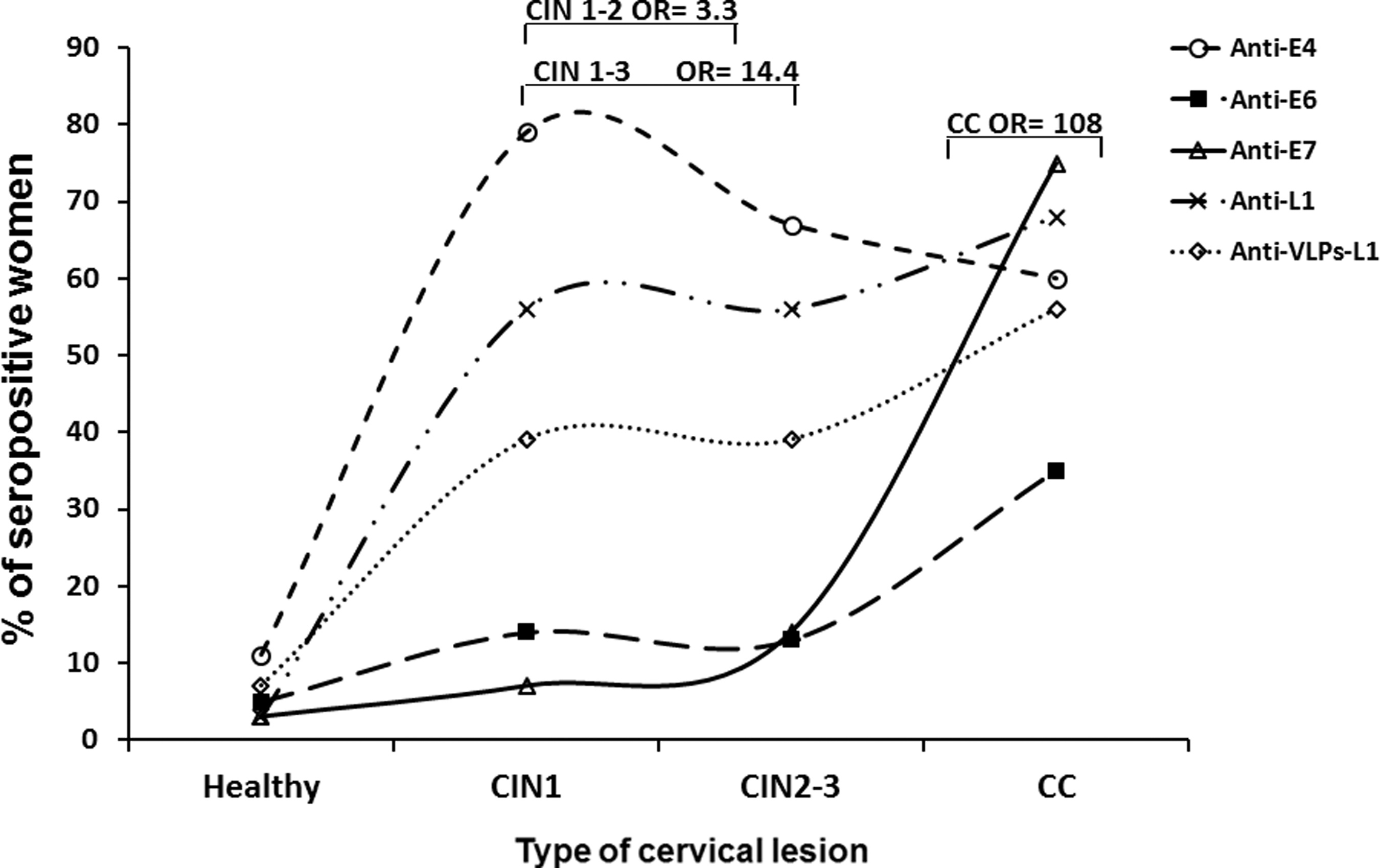
Prevalence distribution of anti-HPV antibodies in different female populations with different degree of cervical lesions and CC. Prevalence from different anti-HPV protein antibodies was taken from different studies and plotted in a graph versus the degree of the lesion at the uterine cervix in different female populations. Anti-E4 antibodies (94,138), anti-E6 antibodies (41), anti-E7 antibodies (94), anti-L1 antibodies (103), and anti-VLPs-L1 antibodies (93) show different patterns of distribution according to the degree of the lesion. Bars show the reported associations (OR) that were statistically significant (p < 0.05) between anti-E4 and anti-E7 antibodies and the different degrees of cervical lesions. These antibodies could be proposed as markers to differentiate the stage of the cervical lesions. CC, cervical cancer; HPV, human papillomavirus; OR, odds ratio; VLPs, virus-like particles.
E4 is the other HPV protein that has been more frequently studied, probably due to its abundance (20–30% of total protein in condylomas) and to its differential production along the viral cycle (31,140). In addition, a high expression of the E4 protein coincides with viral replication in low-grade lesions, while this protein is almost absent in high-grade lesions (31,44,81). In consequence, E4 is proposed as a marker of viral replication (31,57). However, this methodology relies on biopsy samples, which are difficult to obtain. Therefore, the detection of antibodies against HPV antigens has become a better and more ample system to indirectly follow the expression of viral proteins. In this sense, epidemiological studies have shown that a higher prevalence of anti-E4 antibodies is present in women with premalignant lesions than among those with CC or in the general population (18,64,94,123).
Previous studies in our laboratory showed that the prevalence of anti-HPV16 E4 antibodies is low in healthy women (11%), it is elevated in subjects with CIN1–3 lesions (∼66%), and it is reduced in patients with CC (45%) (Fig. 1), all of which suggest early immune recognition of this protein (94). These antibodies were also observed in ∼60% of women with CIN1–2 (138), which suggests that they could serve as early markers of the disease. More recently, it has been reported that anti-E4 antibodies were not associated with any stage of the disease (18,72), but this could be due to the fact that the HPV antigens used in the protein microarrays and in the Luminex multiplex assays were not purified and high backgrounds may hinder the sensitivity of the system.
Another well-characterized antibody response in premalignant and cancer lesions is the one against E2 protein. The prevalence and association with the anti-E2 IgA were higher in CIN1–2 lesions (64%) and decreased with the increasing severity of the cervical lesion (CIN3, 31% and CC, 9%) (74,104). Recently, another group has reported that anti-E2 seroprevalence is low among women with CC; they also did not observe an association with this stage of the disease (18). Overall, these data suggest that anti-E2 antibodies could constitute a good biomarker for CIN lesions, but this needs further studies.
Proteins, E1 and E5, are the less investigated HPV antigens since only a few studies have described the prevalence of antibodies against them. The prevalence of anti-E1 antibodies has been found to be very low in healthy women and women with CC (0.3% and 10%, respectively) (18,64). Anti-E5 antibodies have been analyzed by protein microarrays and no associations were found with any stage of the disease (72). Nevertheless, more studies need to be carried out to better characterize the anti-E5 antibody response, which could be an interesting serological marker, since it was observed that the presence of E5 mRNA was associated with low-grade lesions (71).
The detection of antibodies against late HPV proteins has been carried out specifically through the major L1 capsid protein. The normal anti-L1 immune response generates neutralizing antibodies. However, this response is slow since the average seroconversion is between 8 and 9 months after the first HPV DNA detection; antibody titers are low, and only between 50% and 70% of women seroconvert (Table 2) (14). Furthermore, anti-L1 antibodies persist for at least 10 years and remain protective against the HPV infection in the great majority of women exposed to it (120).
Initially, several studies have evaluated anti-L1 antibodies through ELISA-L1 (denatured protein), showing their prevalence to be higher in patients with low-grade lesions and lower among those with high-grade lesions and CC (41,103,135). However, detection of anti-L1 antibodies has also been reported among healthy women (28), which could be pointing to a previous infection. More recently, the generation of L1 VLPs has allowed the development of a more sensitive and more specific system to measure the L1 neutralizing antibodies as well as the conformational epitopes (143).
In the beginning, VLPs were produced in baculovirus and vaccinia virus and they were used to detect HPV-positive women (60,147). More recently, a VLP-based ELISA has been developed and it has been used in epidemiological studies to characterize the anti-VLPs-L1 immune response in different populations. It was found that these antibodies are associated with CIN3 and CC (139), while another group showed that anti-VLPs-L1 IgA was associated with clearance of the infection (11). Several groups have characterized the anti-VLPs-L1 antibodies in different female populations and they have found a prevalence between 7% and 43% in controls, between 25% and 68% in CIN lesions, and between 43% and 100% in CC (Table 2) (19,49,79,116,124,133). Overall, these results suggest that anti-VLPs-L1 antibodies increase with the severity of the cervical lesion (Fig. 1) and are probably not protecting the system since the cervical lesions continue progressing. The variations regarding the ELISA-VLPs-L1 assays among the different laboratories could be accounted for by the origin of the VLP production (baculovirus vs. yeast) and the different purification systems.
More recently, several groups have been using the new Luminex multiplex technology, which allows higher sensitivity and specificity to analyze several L1 antigens from different HPV types. This assay has been used in epidemiological studies and it has detected higher seroprevalence against L1 HPV antigens in healthy female populations than among women with CC. These results corroborate previous observations that suggested that the higher the evolution of the disease, the lesser the anti-L1 antibodies, although the opposite is observed in the case of anti-E7 antibodies (117).
The presence of anti-HPV antibodies has been analyzed in patients with a different degree of HPV-associated cervical lesions and they have been proposed as markers of disease progression (115) and for CC diagnosis (55,80,84); however, no consistent results have been shown in the different studies that have engaged this task.
Diagnostic and Prognostic Tests for HPV-Related Cervical Lesions
The use of the Papanicolaou (Pap) smear, which is the primary diagnostic test in most cancer prevention programs in developed countries, has helped to reduce the global burden of CC, but this has not been the case in developing countries. The main problem is the low and variable sensitivity of the Pap test (50–84%), which makes the identification of premalignant lesions difficult (2,48). Therefore, more sensitive and specific diagnostic systems to detect earlier stages of the disease and to prevent the development of CC are necessary. In this sense, HPV DNA testing has emerged as a good candidate to replace the cytological technique because although its specificity is low (47–80%), its sensitivity to detect precancerous lesions is high (90–100%), which makes it suitable as a screening test (126,127,132). However, the presence of HPV DNA is not indicative of an active infection, so it has been necessary to develop new diagnostic systems to corroborate possible progression to cancer.
Many studies have focused on the search for new biomarkers associated with the early stages of the HPV disease. These include detection of E6/E7 mRNA, methylation, and detection of p16INK4a, which have shown high specificity and low sensitivity, which make them suitable as prognostic tests (5,23,134). These tests, in conjunction with the Pap smear or the HPV DNA detection, could more accurately identify patients at risk of developing CC. Up to now, the use of the p16INK4a protein marker has been justified when there is a discrepancy between pathologists determining the presence of high-grade lesions. In these cases, an overexpression of the p16INK4a protein has been observed in high-grade cervical lesions, but not in low-grade ones (13).
Other surrogate biomarkers for CC include MCM2, top 2 protein (7,21,26), epidermal growth factor cell surface receptors (88,110), cell cycle inhibitors, p21 and p27 (69), COX-2, vascular endothelial growth factor, and Cav-1 (53), all of which have been shown to detect HPV-associated CC and high-grade lesions or to predict the behavior of the disease before treatment (25). A direct detection of HPV E6/E7 proteins has also been carried out in cell scrapes or cervical tissue samples, but they have low sensitivity as less than 1% of the cells are infected and express the oncogenic proteins (34,98,144). Although all of these tests have high specificity, they still depend on a tissue sample or a cervical lavage where the number of HPV-infected cells is reduced (<1%), making it difficult to identify real patients at risk of developing CC (45).
In this sense, there is an increased need to look for a test or a combination of tests that are inexpensive, highly sensitive and specific, and less invasive. Thus, the detection of anti-HPV serum antibodies has become a good alternative for new biomarkers to detect CC-associated premalignant lesions as this is a naturally amplified system that allows the detection of low viral antigen concentrations in patients at risk of developing CC associated with persistent HPV infection (45). Until now, it has not been determined whether HPV antibody profiles would be useful for screening, prognosis, monitoring therapy, or predicting recurrence of CC. More prospective studies with a larger panel of HPV antigens to detect HPV antibodies need to be carried out to explore and corroborate the usefulness of these antibodies as biomarkers for the detection and prediction of CC-associated premalignant lesions.
The detection of antibodies against HPV antigens has been carried out through ELISA, Western blot, RIPA (radioimmunoprecipitation assay), and, more recently, through variations of these techniques such as Luminex multiplex or protein microarrays. All of them have shown that anti-HPV antibodies can be useful biological markers of different types of lesions in the uterine cervix.
The potential of HPV antibodies as biomarkers for CC is based on the fact that the sequential expression of early HPV proteins in the cervix had correlated with the serological data and this might be useful to identify previous, present, and persistent infections that could be related to the progression of the disease. By looking at the antibody response against other viral infections, it has been cleared that specific antibody isotypes could be indicators of an early or of a current infection (IgM), of an infection in the mucosal tissue (IgA), or of past or persistent infections (IgG) (90). Thus, the presence of serum IgG antibodies to HPV16 VLPs-L1 was found in women around 8 months after the detection of HPV16 DNA (15); they were more frequently detected among those individuals with a high viral load, with persistent infection, and with high-grade squamous intraepithelial lesion or CC (33–41%) (51,139).
In HPV-negative women, anti-L1 HPV16 IgG antibodies were associated with previous virus exposure, while VLPs-L1 IgA was associated with virus clearance in women older than 50 years of age, probably due to infection persistence because of immune senescence (51,75,108,139). Isotype profiles against HPV16 capsids, such as IgA and IgG together, are frequently found in women with persistent HPV16 infection and they might confer protection against future infections with this or other related types (50). Antibodies against HPV16 E6 and E7 have been detected late during the development of CC and point to a bad prognosis (97). In a retrospective Scandinavian study, anti-E6 and anti-E7 IgG antibodies were identified between 0.5 and 5 years before CC diagnosis, suggesting the usefulness of these antibodies as predictors of the disease (67).
Other antibodies that have been studied and are associated with premalignant lesions (CIN1–2) are the anti-E4 antibodies (odds ratio [OR] = 3.3) and it has been suggested that they can be markers of viral replication (138). However, prospective studies need to be carried out to determine the usefulness of these antibodies as early markers and/or predictors of the disease.
Improvement of early detection of the disease depends on the combination of different biomarkers that allow to increase the sensitivity and specificity of diagnostic systems to detect women at risk of developing CC. Few attempts to combine biomarkers for diagnostic procedures have been carried on. For instance, in a prospective sero-epidemiological study in Europe (EPIC, European Prospective Investigation into Cancer and Nutrition), antibodies against HPV, Chlamydia trachomatis (CT), and herpes simplex virus 2 (HSV-2) were studied, and high associations between HPV16 E6 antibodies and invasive CC (ICC) were found (OR = 10); high associations were also found between CIN3 and anti-HPV16 L1 antibodies (OR = 2.4). The results also showed that the coinfection with CT or HSV-2 increased the persistence of the HPV infection as well as the risk of developing CC (16).
Similar results were also observed with oropharyngeal cancer, where a strong association between HPV16 E6 seropositivity and the disease suggests that these antibodies can be predictive markers of the disease as they were present more than 10 years before the cancer diagnosis (3,63).
Recently, in a more detailed serological study that looked for predictive, prevention, and treatment biomarkers in women with cervical precancerous disease, Lazarenko et al. showed that low-avidity anti-HSV IgG antibodies were present in HR-HPV-positive patients and associated with an immunosuppressive state (decreased IFN-γ and increased levels of IL-10 and TGF-β1), suggesting that these HSV antibodies could be useful markers regarding treatment decisions (65). These researchers also suggest that a combined panel of biomarkers (already validated and potential ones) could help to increase the sensitivity and specificity of screening tests, diagnostic procedures, and treatment options to reduce the incidence and mortality of CC.
Combined markers have already been used to detect women at risk for CC. Such is the case of p16ink4a (negative regulator of the cell cycle) and Ki-67 (proliferation marker) surrogate markers for the presence of the oncogenic HPV16 E7 protein. Low sensitivity and specificity of these biomarkers have been observed by immunohistochemistry, but in combination, these markers can identify more than 90% of the HPV-positive women with high-grade lesions (13,96,113).
Therefore, the humoral immune response is an important tool to combine biomarkers and it could allow the determination of the infection stage (transient, latent, persistent) and of the stage of the disease. Thus, it would be necessary to identify immunoglobulin profiles (IgM, IgA, IgG) against different HPV antigens (E2, E4, E5, E6, E7, L1) that enable the identification of the infection state, especially a persistent infection, which is important for the progression to CC. However, more detailed studies of the humoral immune response against HPV need to be carried out to clarify the usefulness of these antibodies for diagnosis.
Conclusions
Several authors have proposed the presence of antibodies in different HPV cervical lesions as markers of disease progression, but not enough studies that validate their use for diagnosis support this. Recently, it has become clear that cytology and colposcopy tests detect cervical lesions at very late stages of the disease. The introduction of HPV DNA detection as a screening test has increased the identification of high-grade lesions, but it is still not enough to distinguish an active infection that could progress to CC, especially because these premalignant lesions can undergo regression in around 70% of the cases. Thus, the introduction of tests that measure the presence of active HPV infections (E6/E7 mRNA screening, detection of early viral proteins, or the presence of anti-HPV antibodies) could be useful to achieve a more accurate diagnostic system.
The advantage of using the antibody response lies in the fact that it is a naturally amplified system with high sensitivity and specificity to identify premalignant lesions and it is able to detect very low concentrations of viral antigen. Furthermore, the high amino acid sequence identity between proteins from different HPV types, which can range from 36% for E4 and up to 74% for L1, enables high antibody cross-reactivity among different HPV types, making the system highly sensitive and specific.
Moreover, the sequential expression of the viral proteins depends on the viral cycle, allowing the identification of different stages of the infection, such as replication (associated with E4 protein), transformation (associated with E6/E7 proteins), and past infections (associated with L1 protein). Meanwhile, the immune system generates an antibody response against the different HPV antigens, which allows the identification of different stages of the infection indirectly. Therefore, this creates the necessity to investigate more closely the anti-HPV antibody response to evaluate the overall HPV infection and simultaneously assess the usefulness of these antibodies as markers of the stage and/or of progression of the disease.
Footnotes
Acknowledgments
This article was carried out within the PhD program in Infectious Diseases of the National Institute of Public Health. D.A.S.-P. was sponsored by the CONACYT postgraduate fellowship program (No. 218122) and through CONACYT grants, 87075 and 168635, and PROMEP-PIFI (UAEMOR-CA-26). The authors thank Adela Iglesias Morineau for editing the English version of this article.
Author Disclosure Statement
No competing financial interests exist.