
Abstract
The H7N9 influenza virus emerged in February 2013 in China, and underlies over 20% of human mortality in the country. Many efforts are being made to develop an effective vaccine against this highly pathogenic virus. We made H7N9 vaccine virus with six internal genes of A/PR/8/34 (H1N1) and two surface genes of hemagglutinin and neuraminidase from A/Anhui/1/2013 (H7N9) by reverse genetics, and the H7N9 vaccine antigens were produced in eggs. Protective antibodies were induced in mice immunized with a single dose (7.5 μg) of the H7N9 antigen. These mice survived lethal infection by the H7N9 virus, although few viruses were found in their lung tissues. However, mice administered with two doses of the H7N9 antigen survived without any viral antigen being detected in their lung tissues. Furthermore, the IgG antibody subtypes were also pronounced in lung tissues of the immunized mice. Therefore, our results suggest that the inactivated whole antigen of the H7N9 influenza virus might protect animals and humans from its lethal infection.
Introduction
I
During the 20th century, there were three human pandemics caused by the H1N1, H2N2, and H3N2 subtypes of the influenza A virus (12,17,21). The pandemic in 1918, which is caused by the H1N1, resulted in over 50 million human deaths (21). The 21st century faced a much milder pandemic caused by the swine-origin H1N1 virus (9,11,13). In the event of another pandemic, the H5 and H7 subtypes might be possible candidates since these subtypes of the influenza virus sporadically infect humans (14,15,20).
A novel influenza virus against which humans are not immune, H7N9, emerged in China in February 2013 (5,24). As of March 31, 2015, 253 of the 631 humans infected with the H7N9 virus died (8). These individuals were infected in their lower respiratory tracts with severe clinical signs of pneumonia, respiratory failure, acute respiratory distress syndrome, and multiorgan failure (6). The H7N9 virus binds to its avian and human receptors (α2,3- and α2,6-linked sialic acid, respectively); and replicates in the epithelial cells of the human lower respiratory tract (22).
Vaccination is one of effective ways to prevent influenza virus infections in humans. There are two kinds of licensed vaccine against seasonal influenza viruses: live attenuated and killed or subunit (10,16). Among them, killed or subunit vaccines produced in embryonated eggs have been widely used.
Studying the immunogenicity of inactivated H7N9 antigen using an animal model is important for the development of an effective vaccine against the pandemic-potential H7N9 influenza virus.
In this study, we constructed an H7N9 vaccine strain with six internal genes (PB2, PB1, PA, NP, M, and NS) of A/PR/8/34 (H1N1), and two genes (HA and NA) of A/Anhui/1/2013 (H7N9), and produced the inactivated whole antigen in embryonated eggs. We then assayed the efficacy of the vaccine in outbred ICR mice. Additionally, we determined the vaccine dose that can protect mice from the lethal H7N9 challenge, as well as the antibody subtypes related to this protection.
Materials and Methods
Virus and animals
The H7N9 influenza virus (A/Anhui/1/2013) was obtained from the World Health organization (WHO) Collaborating Center for Influenza, Centers for Disease Control and Prevention. It was amplified in 10-day-old embryonated eggs in an incubator at 37°C. Experiments with the H7N9 virus were performed in a biosafety level 3 (BSL-3) facility approved by the Korean government.
Outbred ICR mice were bred in the animal facility of the Chungnam National University. Seven-week-old female ICR mice were used for the vaccine study.
Generation of the H7N9 vaccine strain
The reassembled H7N9 vaccine virus was constructed with the A/PR/8/34 (H1N1) backbone as described previously (18). Reverse genetics plasmids of PB2, PB1, PA, NP, M, and NS of A/PR/8/34 (H1N1) were kindly provided by Dr. Robert G. Webster of the St. Jude Children's Research Hospital (Memphis, TN). Reverse genetics plasmids of the HA and NA genes of A/Anhui/1/2013 (H7N9) were constructed as follows: The cDNAs were synthesized from RNAs extracted from allantoic fluids containing A/Anhui/1/2013 (H7N9) using the RNeasy Kit (Qiagen), uni12 primer (5′-AGCAAAAGCAGG-3′), and ImProm-II™ Reverse Transcription System (Promega Corporation). The HA gene of A/Anhui/1/2013 (H7N9) was amplified by polymerase chain reaction (PCR) using the Expand High-Fidelity PCR System (Roche Diagnostics) and the following primers:
Bm-HA-1 (5′TATTCGTCTCAGGGAGCAAAAGCAGGGG-3′) and
Bm-NS-890R (3′-ATATCGTCTCGTATTAGTAGAAACAAGGGTGTTTT-5′)
Meanwhile, the NA gene of A/Anhui/1/2013 (H7N9) was amplified with the primers: Ba-NA-1 (5′-TATTGGTCTCAGGGAGCAAAAGCAGGAGT-3′) and
Ba-NA-1413R (5′-ATATGGTCTCGTATTAGTAGAAACAAGGAGTTTTTT-3′)
Both the amplified PCR products were cloned into the reverse genetics vector pHW2000 as described previously (7). The reason why the reassembled H7N9 vaccine strain was made is that the wild-type H7N9 virus cannot be used for the vaccine production in general vaccine production facility since this virus is highly pathogenic to humans.
To rescue the reassembled H7N9 vaccine virus, Vero cells grown in a six-well plate were transfected with 1 μg of each plasmid using 18 μL of the TransIT LT-1 tranfection reagent (Panvera). The transfected cells were incubated for 72 h, and 200 μL of the supernatants were treated with L-(tosylamido-2-phenyl) ethyl chloromethyl ketone (TPCK)-treated trypsin (1 μg/mL) before they were inoculated into the allantoic cavities of 10-day-old embryonated eggs. The rescued viruses in the allantoic fluids of the inoculated eggs were identified by genetic analyses of HA and NA genes.
Purification of the inactivated H7N9 vaccine antigen
The reassembled H7N9 viruses were inoculated into the allantoic cavities of 10-day-old embryonated eggs at 35°C for 72 h before the allantoic fluids were collected. The viral titer of the reassembled virus was 107 of egg infectious dose 50/mL (107EID50/mL). The collected fluid was then concentrated using the Amicon concentrator apparatus. Next, the concentrated viruses were purified by a continuous gradient of 20–75% sucrose (26,000 rpm, 4°C) for 2 h. To inactivate it, the purified vaccine virus was treated with formalin (0.02% per volume) at 4°C overnight. The inactivated virus was then spun down by centrifugation at 13,000 g (4°C) for 2 h. Finally, the concentration of whole protein in the purified H7N9 vaccine was determined by the Bradford method (Bio-Rad) against a bovine serum albumin (BSA) standard curve.
Vaccination of outbred mice
The H7N9 vaccine antigen from 0.12 μg to 7.5 μg was intramuscularly (i.m.) inoculated into the hind legs of 8-week-old female ICR mice (n = 10 per) and 4 weeks later, the immunized mice were then intranasally (i.n.) infected with 50 μL of 106 EID50 (egg infectious dose 50) of A/Anhui/1/2013 (H7N9). The mice were observed for changes in body weights and mortality for 14 days post infection (p.i.). The one-dose immunized mice with 7.5 μg antigen were boosted on 4 weeks after the first inoculation with 7.5 μg antigen. All animal experiments were approved by the Animal Experimental Ethics Committee of the Chungnam National University.
Measurement of viral titers in lung tissues
Lung tissues of the mice immunized with one or two doses of the H7N9 vaccine antigen (7.5 μg) and infected with A/Anhui/1/2013 (H7N9) were homogenized in 0.5 mL of phosphate-buffered saline (PBS) (pH 7.4) mixed with 2× antibiotic–antimycotic solution (Sigma-Aldrich). Next, supernatants of the homogenized lung tissues were serially diluted 10 folds in PBS (pH 7.4), and each dilution was inoculated into 10-day-old embryonated eggs (n = 4 per dilution). The viral presence in the inoculated eggs was identified by hemagglutination inhibition (HI) assay using 0.5% turkey red blood cells. Viral titers were then determined by calculating log10EID50/mL. The detection limit of the virus was 1 log10EID50/mL.
Determination of antibody titers in the immunized mice by HI assay
Sera collected from the immunized mice were treated with receptor-destroying enzyme (RDE) (DENKA SEIKEN) to inhibit nonspecific reactions. The RDE-treated sera were serially diluted two folds in PBS (pH 7.4) in V-bottom 96-well plates. They were then reacted with an equal amount of eight HA units (25 μL) of A/Anhui/1/2013 (H7N9). The plate was incubated for 15 min at room temperature, and then 50 μL of 0.5% turkey red blood cells were added to each well. The reaction was performed for 40 min before the results were evaluated. The HI titers were expressed as reciprocal dilutions that completely inhibited hemagglutination.
Histopathology and antigen staining of the lung tissues
Lung tissues from mice used for the viral titer measurements were fixed in 10% neutral buffered formalin and embedded in paraffin. Sections (5 μm) were stained with Hematoxylin and Eosin (H&E), and observed under an Olympus DP70 microscope (Olympus Corporation) as described previously (1).
Tissue sections (5 μm) were deparaffinized, hydrated in distilled water, and fixed in 100% chilled acetone for 2 h before they were stained with mouse anti-influenza A virus nucleoprotein (NP) antibody (Serotech). Endogenous peroxidase activity in the sections was blocked by incubation in 3% H2O2 (15 min, 37°C) followed by blocking with 5% BSA in PBS (pH 7.4) at room temperature for 1 h. The blocked tissue sections were then reacted with the mouse anti-influenza NP antibody (1:1,000 dilution) by incubating at room temperature for 1 h. The reacted tissue sections were stained with biotin-labeled goat anti-mouse immunoglobulin, VECTASTAIN ABC-AP, and Vector red alkaline phosphatase substrate (Vector). Finally, the stained tissue sections were counterstained with Hematoxylin QS (Vector) before they were examined under the Olympus DP70 microscope.
Detection of antibody subtypes in the immunized mice
Mice immunized with two doses of the H7N9 vaccine antigen were euthanized with high doses of Zoletil (Virbac Laboratories). A through perfusion was carried out to deplete blood before tissues (lung, liver, intestine, brain; 0.05 g each) were collected in 0.5 mL PBS (pH 7.4). The collected tissues were homogenized to determine the antibody subtypes using Enzyme-Linked Immunosorbent Assay (ELISA).
Each well of a 96-well ELISA plate (Greiner bio-one) was coated with 100 μL (0.05 μg) of the purified and inactivated H7N9 antigen of A/Anhui/1/2013 (H7N9) [diluted with 1 mL of carbonate–bicarbonate buffer (pH 9.6)]. Hundred μL of each supernatant of the homogenized tissues [in PBS (pH 7.4) containing 5% horse serum and 0.05% Tween 20 (PBS-Tween 20)] were added to the coated wells and incubated at room temperature for 1 h followed by washing with PBS-Tween 20 thrice. Hundred microliters (1:1,000 dilution in PBS-Tween 20) of horseradish peroxidase (HRP)-conjugated goat anti-mouse IgA (KPL), IgE, IgG1, IgG2a, IgG2b, IgG2c, IgG3, and IgM (SouthernBiotech) were then added and reacted at room temperature for 1 h. This was followed by washing with PBS-Tween 20 thrice. Next, to each well, 100 μL of ABTS peroxidase substrate (KPL) was added and incubated at room temperature for 30 min before ABTS peroxidase stop solution (KPL) was added to stop the reaction. Finally, the optical densities (OD) were read at 405 nm in an ELISA microplate reader (TECAN systems, Inc.).
Statistical analysis
Statistical significance was determined by a two-tailed, paired Student's t-test. p values <0.05 were considered significant.
Results
Antibody induction in outbred mice immunized with inactivated whole H7N9 antigen
Outbred ICR mice (n = 10) were inoculated i.m. with the inactivated H7N9 vaccine doses from 0.12 μg to 7.5 μg. The H7N9 (A/Anhui/1/2013) antibody titers determined by HI assay 4 weeks after the immunization are shown (Fig. 1A). The mean HI titer after the single-dose vaccination was from 53 to 144. The results suggest that the inactivated whole H7N9 vaccine is immunogenic in the outbred ICR mice.
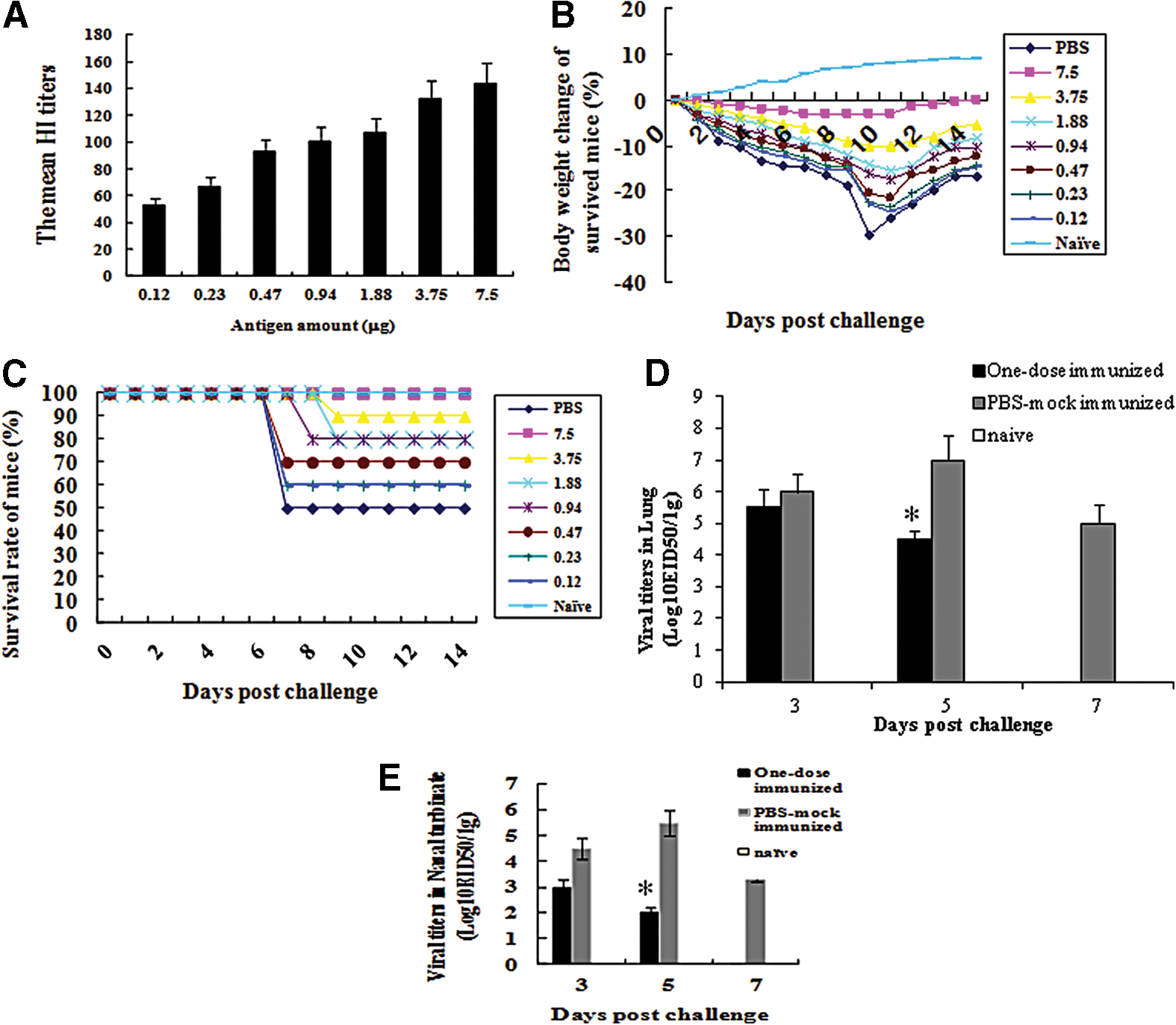
Antibody titers in the immunized outbred mice, the change of body weights and mortality of the one-dose immunized outbred mice infected with H7N9 influenza virus, and the viral titers in the lung tissues of the one-dose immunized outbred mice infected with H7N9 influenza virus.
Clinical signs and viral titers in outbred mice immunized with a single dose of the inactivated whole H7N9 antigen
Since a pandemic situation might lead to vaccine shortage, we first investigated whether a single dose of the inactivated whole H7N9 antigen can protect the outbred ICR mice against lethal challenge with the H7N9 (A/Anhui/1/2013) influenza virus. The outbred mice (n = 10) were immunized i.m. with a single dose of the inactivated whole H7N9 antigen from 0.12 μg to 7.5 μg, and 4 weeks later, they were challenged with the H7N9 virus. Our results demonstrated that overall, the protection rates of mice immunized with single dose of vaccine antigens depended on the inoculated antigen amount (Fig. 1B, C). The immunized mice, with 7.5 μg antigen and infected with the virus, lost body weights up to 2.5% and 3.2% on 5 and 9 days p.i, respectively, while those immunized with 0.12 μg antigen and infected with the virus lost body weights up to 12.5% and 23.0% on 5 and 9 days p.i., respectively. The mice mock immunized with PBS and infected with the virus lost body weights up to 29.5% on 9 days p.i, compared with those before infection (Fig. 1B).
No mortality was observed in the infected mice immunized with 7.5 μg antigen, however, the mock-immunized and immunized mice with 0.12 μg showed 50% and 40% mortality, respectively (Fig. 1C).
We further used 7.5 μg antigen to find out the viral titers and tissue pathology in mice since this dose protected the immunized mice from H7N9 challenge with the mild loss of body weights compared to those in mice immunized with other lower doses. In addition, children who are less than 36 months old receive 7.5 μg antigen of seasonal influenza vaccine (23).
Viral titers were detectable in lung and nasal turbinate tissues of both the single-dose immunized and infected, and mock-immunized and infected mice. In lungs, viral titers were 5.5 and 6.0 log10EID50/g on 3 days p.i., while those were 4.5 and 7.0 log10EID50/g on 5 days p.i. in the immunized and mock-immunized mice, respectively (Fig. 1D). In nasal turbinates, viral titers were 3.0. and 4.5 log10EID50/g on 3 days p.i., while those were 2.0 and 5.5 log10EID50/g on 5 days p.i. in the immunized and mock-immunized mice, respectively (Fig. 1E). However, no virus was detected in the immunized mice, although the viral titers in the mock-immunized mice were 5.0 and 3.5 log10EID50/g on 7 days p.i. in lungs and nasal turbinates, respectively (Fig. 1D, E).
Viral antigens in lung tissues of the single-dose immunized mice
The lung tissues were stained with H&E to evaluate the pathological damage, and the viral antigen was stained with influenza A anti-NP antibody. Lung tissues of the mock-immunized mice revealed numerous inflammatory cells on 3, 5, and 7 days p.i (Fig. 2A–C). Meanwhile, although inflammatory cells infiltrated the lung tissues of the single-dose immunized and infected mice on 3 and 5 days p.i., they were found to be reduced on 7 days p.i. (Fig. 2D–F). The naive lung tissues did not show any infiltration, as expected (Fig. 2G).
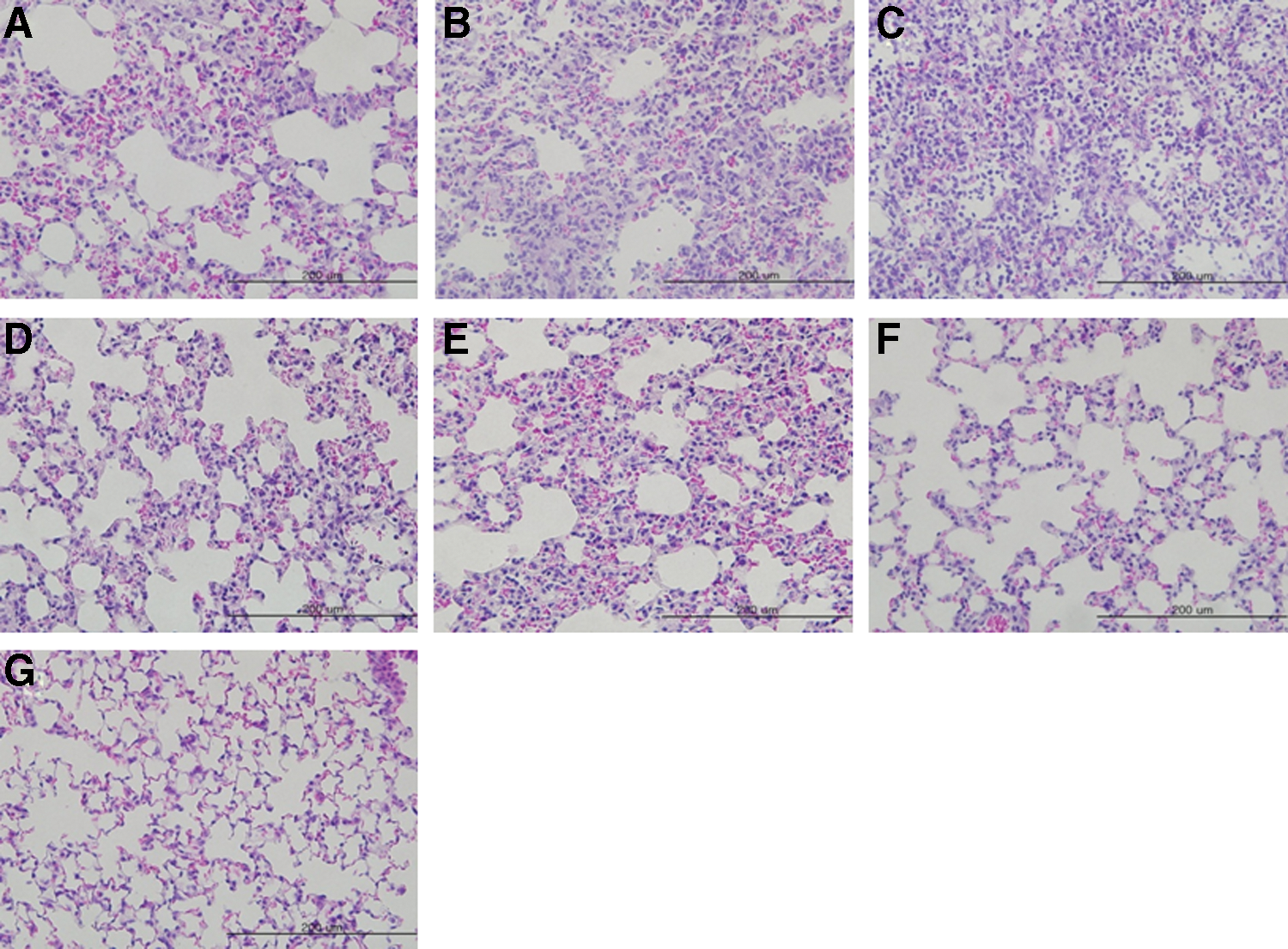
Pathological damage to the lung tissues of immunized outbred mice with one dose of whole inactivated H7N9 antigen. The lung tissues of mice (Fig. 1D) were stained with H&E and observed under the microscope.
Furthermore, antigen staining revealed numerous H7N9 NP antigens in the lung tissues of PBS (50 μL) mock-immunized mice on 3, 5, and 7 days p.i. (Fig. 3A–C). In contrast, lungs tissues of the single-dose immunized and infected mice showed only a few antigens 3 and 5 days p.i. whereas none was detected on 7 days p.i. (Fig. 3D–F). Lung tissues of the naive mice did not show any positive staining (Fig. 3G).
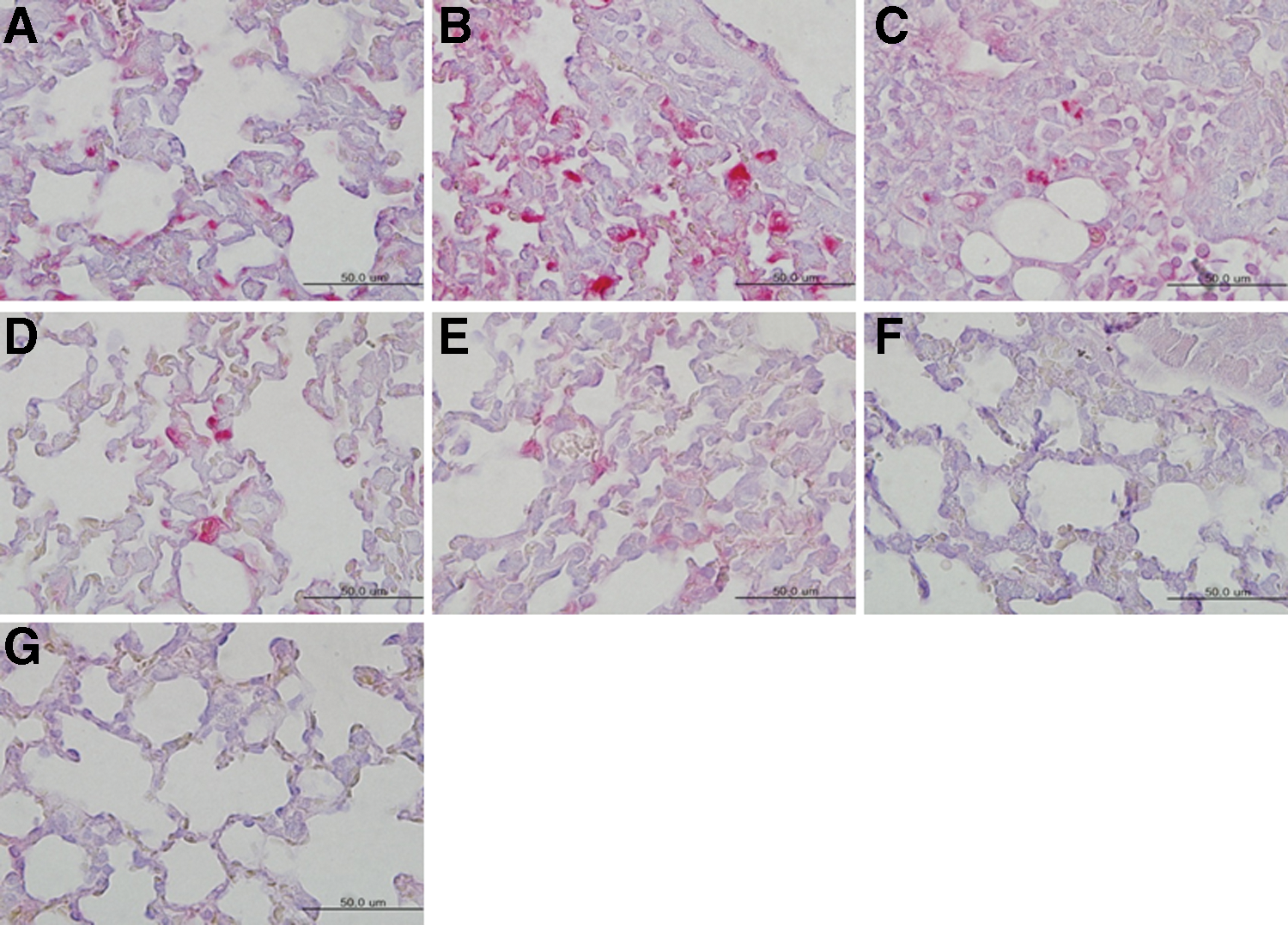
Antigen staining in the lung tissues of immunized outbred mouse with one dose of whole inactivated H7N9 antigen. The lung tissues of mice (Fig. 1D) were stained with antibody of mouse influenza A virus nucleoprotein and red alkaline phosphatase substrate, and observed under the microscope.
Clinical signs and viral titers in mice immunized with two doses of the inactivated H7N9 antigen
A single dose vaccine of the inactivated whole H7N9 antigen was unable to provide sterile immunity to the H7N9 infection. Therefore, we next examined whether two doses of the vaccine can provide the sterile immunity against the virulent H7N9 virus. Outbred ICR mice (n = 10 per group) were immunized with the inactivated H7N9 antigen (7.5 μg) and 4 weeks later, were boosted with the same antigen. They were then challenged with the virus (106 EID50/mL) i.n., and monitored for change in body weights and mortality for 14 days p.i. (Fig. 4A, B). The mean HI titers in mice immunized with two doses of antigens were 150 (data not shown). The two-dose immunized and infected mice showed slight reduction in body weights up to 6 days p.i., but from the seventh day p.i., they caught up with the body weights of the naive controls (Fig. 4A). The mock-immunized and infected mice lost body weights up to 30% compared with those before infection (Fig. 4A). Furthermore, no viral titer was detected in the lung tissues of the two-dose immunized and infected mice, whereas the mean viral titer in the lung tissues of the mock-immunized and infected mice was 6.5 log10EID50/g 5 days p.i. (Fig. 4C).
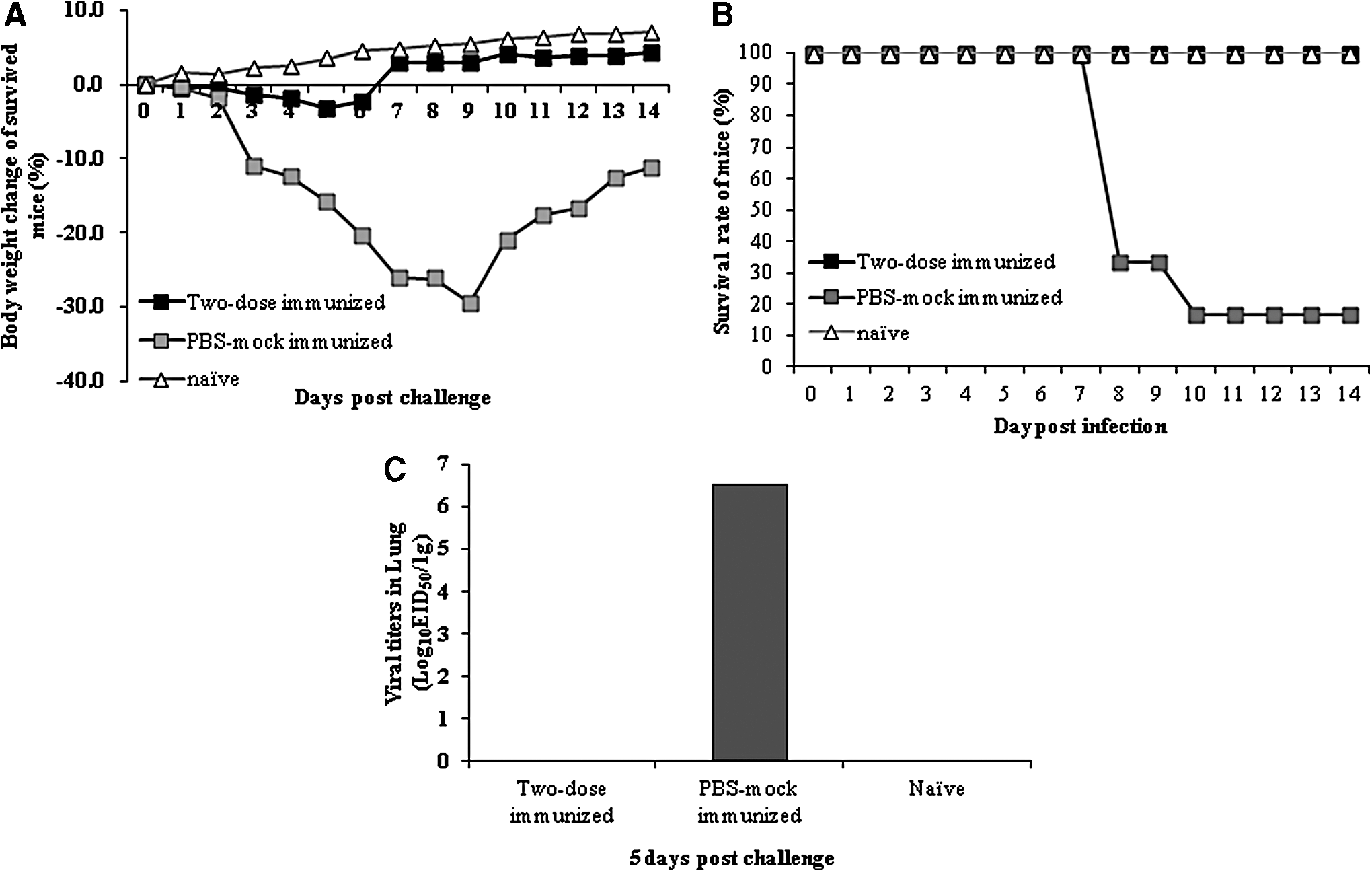
The change of body weights and mortality of the two-dose immunized outbred mice infected with H7N9 influenza virus, and the viral titers in the lung tissues of the two-dose immunized outbred mice infected with H7N9 influenza virus.
Viral antigen in lung tissues of the two-dose immunized mice
Staining of lung tissues with H&E revealed the presence of numerous inflammatory cells in the mock-immunized and infected mice 5 days p.i. (Fig. 5A), but only few in the two-dose immunized and infected mice 5 days p.i. (Fig. 5B). Furthermore, staining with influenza A anti-NP antibody revealed that the lung tissues of the mock-immunized and infected mice were antigen-positive 5 days p.i. (Fig. 5C). No antigen was detected in lung tissues of the two-dose immunized and infected mice (Fig. 5D).
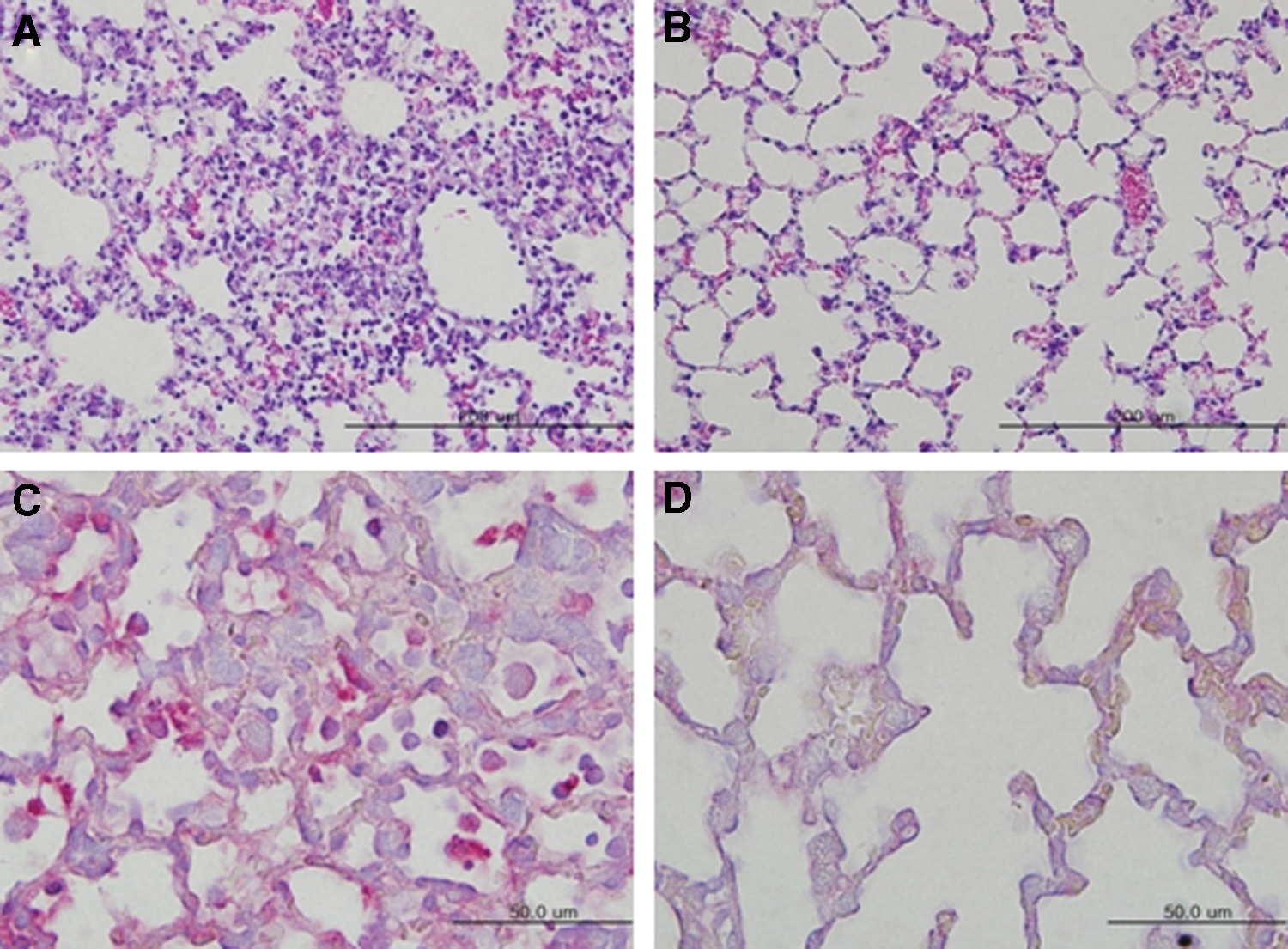
Pathological damage and antigen staining in the lung tissues of immunized outbred mice with two doses of whole inactivated H7N9 antigen. The lung tissues of mice (Fig. 4C) were stained with H&E, with mouse influenza A virus nucleoprotein and red alkaline phosphatase substrate, and observed under the microscope.
Antibody subtypes in the sera and tissues of two-dose immunized mice
To evaluate which antibody subtypes are involved in protecting the immunized mice from the H7N9 influenza virus infection, we collected sera and tissues (lung, liver, intestine, and brain) from the two-dose immunized mice (n = 10) with the H7N9 antigen. Our results showed that the IgG subtype was dominant in both the sera and the tissues (Fig. 6). In the sera, the mean OD values for IgG1, IgG2a, and IgG2b were >1.1 in the two-dose immunized mice. In the lung tissues, the mean OD values for the same subtypes were >0.5 in the two-dose immunized mice.
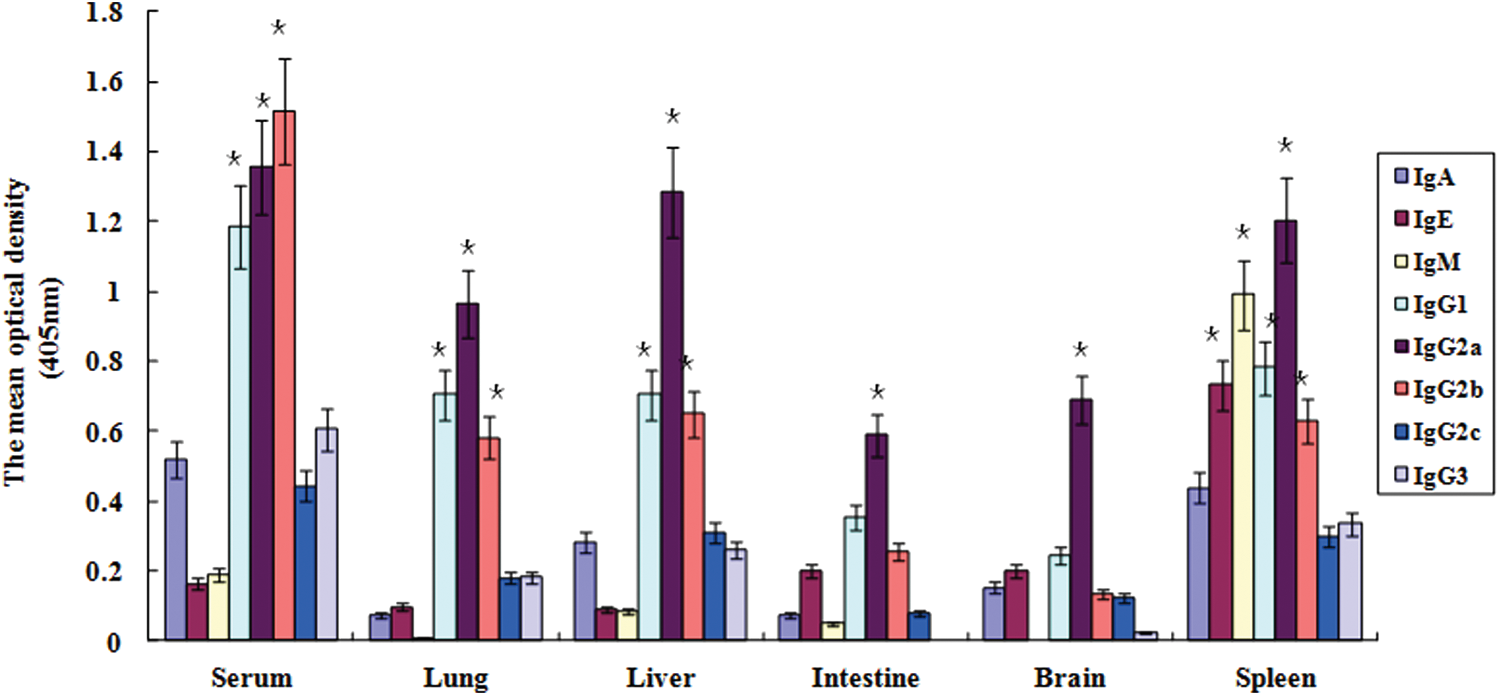
Determination of antibody subtypes in immunized mice with whole inactivated H7N9 antigen. Mice (n = 10) immunized with two doses of H7N9 vaccine antigen were euthanized, and tissues (lung, liver, intestine, and brain) and sera were collected on 6 weeks after vaccination. The subtypes of antibody that bind to H7N9 antigen were detected with HRP-conjugated antibodies of IgA, IgE, IgG1, IgG2a, IgG2b, IgG2c, IgG3, and IgM and with peroxidase substrate. Optical density (OD) was read at 405 nm with ELISA microplate reader. Data are the mean ± standard deviation. The OD value was subtracted from the background of the mock-immunized mice. Statistical analysis was performed between the immunized and PBS mock-immunized mice. *p ≤ 0.05. Color images available online at
Discussion
A novel influenza virus, H7N9, emerged in China in early 2013, which urgently demands the development of an effective vaccine against it. In this study, we developed an H7N9 vaccine strain using reverse genetics and amplified it in the embryonated eggs. We also demonstrated that although the inactivated H7N9 vaccine provides some immunity to mice, two doses of the vaccine are required for sterile immunity. Moreover, the protective immunity was mediated chiefly by the IgG antibody subtype.
Upon testing the efficacy of the inactivated H7N9 antigen in outbred ICR mice, the H7N9 influenza virus was found to be immunogenic when administered with antigens from 0.12 μg to 7.5 μg per dose. The mean HI titer of the mice immunized with a single dose of the H7N9 antigen was 144 four weeks p.i., which indicates that sufficient antibody titer was induced in the mice. Previous studies have also shown that the inactivated whole H7N9 vaccine is immunogenic to mice. For instance, the H7N9 inactivated vaccine grown in Vero cells was found to be immunogenic at two doses (3.75 μg each) in both mice and guinea pigs. The mean HI titers in chicken erythrocytes were 1280 and 155 in the guinea pigs and mice, respectively (26). However, the inactivated H7N9 vaccine grown in MDCK cells was not immunogenic in ferrets immunized with two doses of the whole H7N9 antigen without adjuvant, although the antibody response increased when injected with H7N9 antigen coupled with aluminum hydroxide (3). The mean HI titer was 17 in the ferrets immunized with two doses of the inactivated whole H7N9 antigen (1.5 μg/dose), and 190 in those immunized with two doses of the antigen (1.5 μg/dose) along with aluminum hydroxide (3). The discrepancy observed in our study might be due to the different substrate used for producing the vaccine antigen and different animals used. Specifically, we used eggs and mice, whereas the other study used MDCK cells and ferrets. More study may be needed to find out the immunogenicity of H7N9 vaccine in mice or other animal models by using other vaccine antigens such as 2009 pandemic H1N1 and highly pathogenic H5N1 influenza viruses.
The positive number of viral antigen in the immunostaining of lung tissues was fewer in the one-dose (7.5 μg) immunized mice than in PBS mock-immunized mice on 3 days p.i., however, the viral titers were similar between two groups. The discrepancy may be due to the sensitivity difference between two methods for viral detections, viral titration in eggs, and tissue staining with antibody.
We showed that two doses of the vaccine were needed to provide sterile immunity. Although the single-dose vaccine protected the immunized mice against the lethal H7N9 viral infection, we detected few viruses in their lung tissues along with pathological damage. The two-dose immunized mice, meanwhile, completely protected the immunized mice with no viral antigen or pathological damage observed in their lung tissues. The study with ferrets showed that a single dose of the inactivated whole H7N9 vaccine (15.0, 30.0, or 50.0 μg) successfully reduced the clinical signs, viral shedding, and titers in the lungs of immunized ferrets compared with those of the unimmunized infected ferrets. However, it is important to bear in mind that they did not use two doses of the vaccine (27). Interestingly, a single dose of inactivated whole H7N9 vaccine (10 or 50 μg) prepared from low pathogenic avian influenza virus (A/duck/Monglia/119/2008) isolated from migratory birds provided protective immunity to mice immunized against the H7N9 virus. The viral titers were also significantly reduced in the lung tissues of these mice (4).
Our study further showed that protective immunity by the inactivated whole H7N9 antigen is probably conferred by the IgG antibody subtype. This result is similar to a previous study with ferrets immunized with inactivated whole vaccine of the H5N1 influenza virus. They reported that IgG antibodies were dominant in the tracheae, nostrils, lungs, hearts, livers, kidneys, intestines, spleens, and sera of these ferrets (19).
In markets, the less reactive subunit influenza vaccines containing HA and NA proteins than whole inactivated vaccine are more acceptable in USA and Europe. Therefore, further study on the immunogenicity of subunit vaccine of H7N9 influenza virus may be needed in mice or other animal models.
In summary, our study suggests that the inactivated whole H7N9 vaccine is immunogenic in mice even at a single dose, although two doses of the vaccine are required to induce sterile immunity in the mice.
Footnotes
Acknowledgments
This work was funded, in part, by a Basic Science Research Program through the National Research Foundation of Korea (NRF) from the Ministry of Education, Science and Technology (2015R1A2A2A01003943). This article has been checked for language usage by Editage, an English language editing service.
Author Disclosure Statement
No competing financial interests exist.