
Abstract
While most HIV patients beginning antiretroviral therapy (ART) with advanced immunodeficiency recover CD4+ T cell numbers, the profiles and functions of the newly acquired CD4+ T cells have not been monitored in a resource-limiting setting. In this study, HIV patients (n = 31) from Jakarta, Indonesia, were studied 9 months after commencing ART with nadir CD4+ T cell counts <200 cells/μL. All patients were hepatitis C virus (HCV) seropositive, but asymptomatic. Twelve healthy age-matched controls from the same community were included. CD4+ T cell subsets, immune activation (HLA-DR), and expression of the interleukin (IL)-7 receptor α chain (CD127) were quantitated by flow cytometry. Proliferation (expression of Ki67) was measured following in vitro stimulation (5 days) with anti-CD3 antibody or IL-7. Fifty-two percent of patients recovered CD4+ T cell counts >200 cells/μL over 12 months. At 9 months, patients had fewer naive and CD31+-naive CD4+ T cells, more effector memory (EM) CD4+ T cells, and higher HLA-DR expression on CD4+ T cells than controls. CD127 expression was low on all CD4+ T cell subsets except for naive cells, where it was similar to controls. Similarly, after anti-CD3 antibody or IL-7 stimulation, patients had lower Ki67 expression than controls in all subsets, except naive CD4+ T cells where it was normal or elevated. Overall in the first year of ART, patients had fewer naive and more EM CD4+ T cells. Ongoing immune activation and, antigen-driven stimulation and differentiation of naive T cells may reduce the naive T cell pool, while driving the maturation and accumulation of memory cells with proliferative defects.
Introduction
T
Untreated HIV disease is characterized by perturbations in T cell homeostasis and depletion of the naive T cell compartment, with functional impairments evident in the remaining cells. These changes reflect reduced thymic function and increased immune activation. A failure to regenerate naive T cells through thymopoiesis (15,33) and/or homeostatic proliferation has been associated with incomplete CD4+ T cell recovery on ART (25). The latter may reflect a reduced capacity of naive T cells from HIV-infected patients to proliferate following stimulation with interleukin (IL)-7 and/or T cell receptor (TCR) ligation (6).
IL-7 has a critical role in T cell homeostasis and is required for thymopoiesis and the survival and proliferation of naive and memory T cells (8,32). Circulating levels of IL-7 are elevated during conditions of lymphopenia, such as untreated HIV infection. However, surface expression of CD127 (the IL-7 receptor α chain) on peripheral T cells is diminished; so responses to IL-7 may be impaired (5,28). Levels of IL-7 decrease as CD4+ T cell numbers increase on ART, but expression of CD127 may remain low (7).
Despite prolonged suppression of viral replication with ART, a subset of patients experience ongoing immune activation (17), This may reflect residual HIV viral replication (26), the reactivation of HIV, CMV or other infections from cellular reservoirs (4,12), or the translocation of microbial products from the gut (10). While few studies have compared immune activation in HIV patients treated in Asia with those in resource-rich countries, we demonstrated greater immune activation in patients treated in Malaysia than in Australian patients. Similar results were observed in the healthy control cohorts, with Malaysian controls displaying greater immune activation than controls from Australia (24).
In this study, we examined CD4+ T cell profiles in Indonesian HIV patients coinfected with hepatitis C virus (HCV) (34). Patients were studied 9 months after they started ART with severe immunodeficiency. This scenario is common in populations where the HIV epidemic is fuelled by illicit intravenous drug use. We describe enhanced CD4+ T cell activation and high proliferative responses to IL-7 and anti-CD3 antibody stimulation by naive CD4+ T cells; so increased immune activation and turnover may drive ongoing functional CD4+ T cell deficits.
Materials and Methods
Study population
Samples from HIV/HCV coinfected patients (n = 48) commencing ART at the Pokdisus AIDS Clinic at Cipto Mangunkusumo Hospital (Jakarta, Indonesia) have been used in a series of investigations (18,23,34). Inclusion criteria were age 17–50 years, ART and HCV therapy naive, with CD4+ T cell counts <200 cells/μL. Exclusion criteria included pregnancy, hepatitis B seropositivity, liver cirrhosis, kidney failure, or heart failure. Blood samples were collected before ART, and at 1, 2, 3, 6, 9, and 12 months of treatment. This study used samples collected from 31 patients after 9 months on ART and from 12 healthy controls matched with the patients by age and ethnicity (Table 1; 34).
Median (range).
Log10 copies/mL.
N/A indicates data not available.
HCV diagnoses were based on anti-HCV antibody (Elecsys Anti-HCV Assay; Roche Diagnostics, Branchburg, NJ). Plasma HIV-1 RNA levels were determined by a quantitative reverse-transcriptase PCR assay (Cobas Amplicor; Roche Diagnostics) at baseline and 12 months after ART. CD4+ T cell counts were enumerated after 0, 1, 2, 3, 6, and 12 months of ART, in a routine flow cytometry laboratory at Cipto Mangunkusumo Hospital. The study was approved by the Human Research and Ethics Committee (University of Indonesia). Informed consent was obtained from all participants.
T cell immunophenotyping
Peripheral blood mononuclear cells (PBMC) were obtained by Ficoll-Paque density centrifugation and cryopreserved in liquid nitrogen. Thawed PBMC were washed, resuspended at 106 cells/mL in 1% BSA/PBS, and stained for surface markers (15 min). Fluorescently conjugated monoclonal antibodies used to assess T cell subsets were CD4-FITC, CD45RA-PE, CD27-PerCP-Cy5.5, and CD31-AF647 (BD Biosciences; San-Jose, CA). To assess immune activation, PBMC were incubated with CD4-FITC, CD45RA-PE, HLA-DR-PerCP-Cy5.5, and CD38-APC (BD Biosciences). Cells able to respond to IL-7 were characterized using CD4-FITC, CD45RA-PE, CD27-PerCP-Cy5.5, and CD127-APC (BD Biosciences). To assess α4β7 gut-homing memory CD4+ T cells, PBMC were incubated with CD4-FITC, CD45RO-PeCy7, and β7-APC (BD Biosciences). From the lymphocyte population (identified by forward and side light scatter properties), CD4+ T cell subsets were defined as follows: naive (CD45RA+CD27+), central memory (CM) (CD45RA−CD27+), effector memory (EM) (CD45RA−CD27−), and terminally differentiated effector memory (TD) (CD45RA+CD27−). The CD31+ subpopulation of naive CD4+ T cells was assessed as recent thymic emigrants (20). α4β7 memory CD4+ T cells were identified as CD4+CD45RO+β7hi. Gating strategies are presented in Figure 1. A minimum of 100,000 events per sample were analyzed using a FACS Canto II cytometer (BD Biosciences) and gates were set using appropriate controls. Files were exported in FCS 3.0 format and visualized using FlowJo software, version 7.6.3 (Tree Star, Ashland, OR).
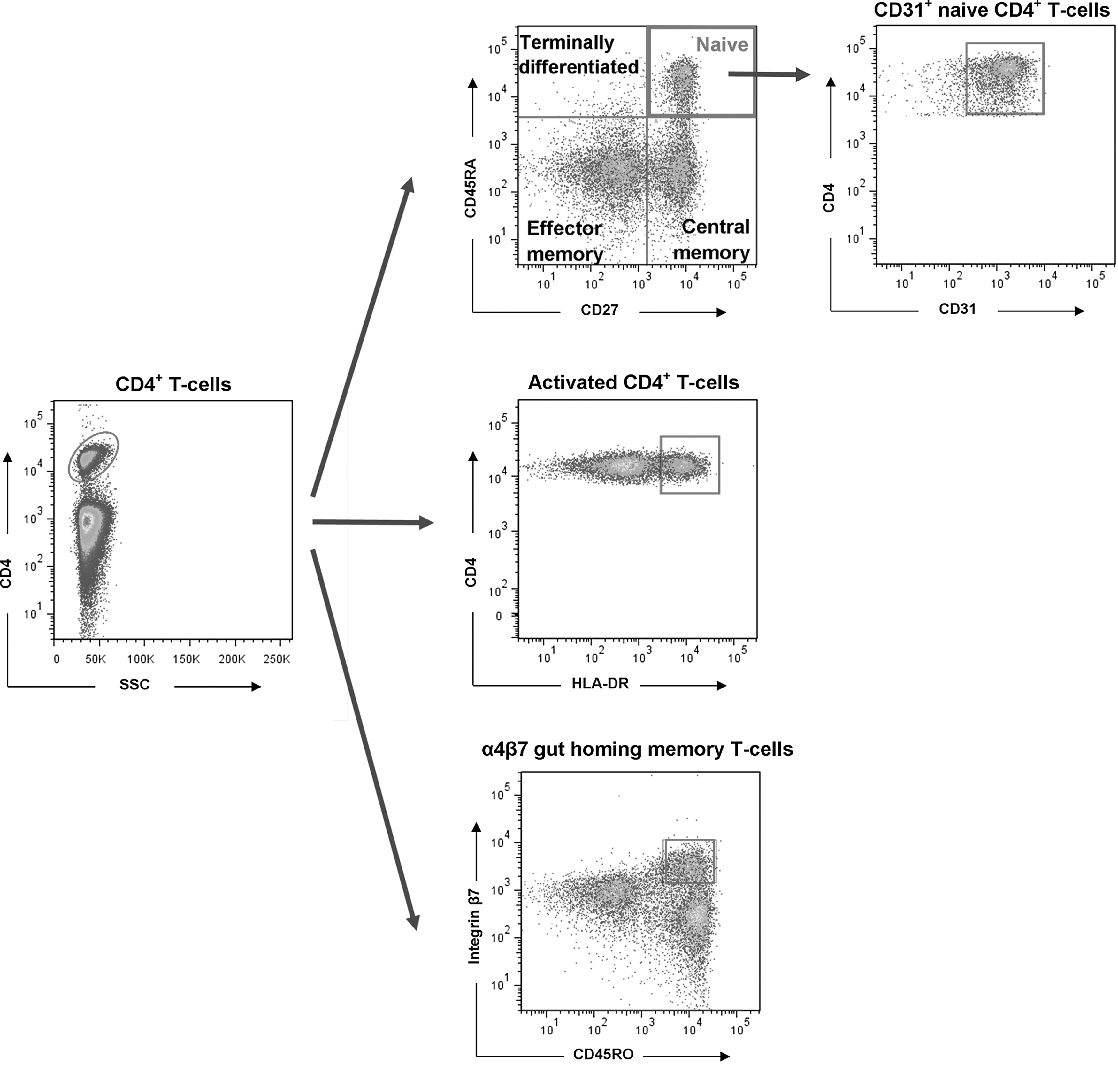
Gating strategy used to define CD4+ T cell subsets. Color images available online at
PBMC cultures and detection of proliferating cells
Thawed PBMC were washed, resuspended at 106 cells/mL in RPMI 1640 with 10% fetal calf serum, and cultured with or without 10 ng/mL anti-CD3 antibody (Mabtech, Stockholm, Sweden) or 5 ng/mL IL-7 (R&D Systems, Minneapolis, MI) at 37°C in 5% CO2 for 5 days. PBMC were stained for surface (15 min) and intracellular (30 min) markers, using the Human FoxP3 Buffer Set (BD Biosciences). Proliferating cells were characterized using CD4-FITC, Ki67-PE, CD27-PerCP-Cy5.5, and CD45RA-APC (BD Biosciences).
Statistical analyses
Statistical analyses were performed using Prism 5 (GraphPad Software, San Diego, CA). Mann-Whitney tests were performed for continuous variables and correlation coefficients were calculated with Spearman's tests. Fisher's exact tests were performed for categorical analyses. p-Values less than 0.05 were considered to be statistically significant.
Results
Patient characteristics
T cell phenotypes were assessed in 31 HIV patients using PBMC cryopreserved after 9 months on ART and in 12 healthy controls. All patients were HCV-seropositive with minimal HCV-associated liver fibrosis or necrosis, and stable HCV RNA loads (18,34). Patients identified unsafe intravenous drug use as a probable route of HCV and HIV transmission. Demographic and clinical data are presented in Table 1. Patients were predominantly male and groups were matched for age (p = 0.25) and gender (p = 1.0). ART regimens, coinfections, and liver histology have been described previously and the patients investigated in this study are representative of the parent study (18,34).
The median (range) CD4+ T cell count rose from 24 (45–415) cells/μL after 12 months of ART, indicating steady immune recovery in most patients (Fig. 2A). Baseline CD4+ T cell counts before ART correlated with counts at 12 months (r = 0.54, p = 0.002). HIV-1 RNA levels declined below 400 copies/mL in 94% of patients after 12 months of ART.
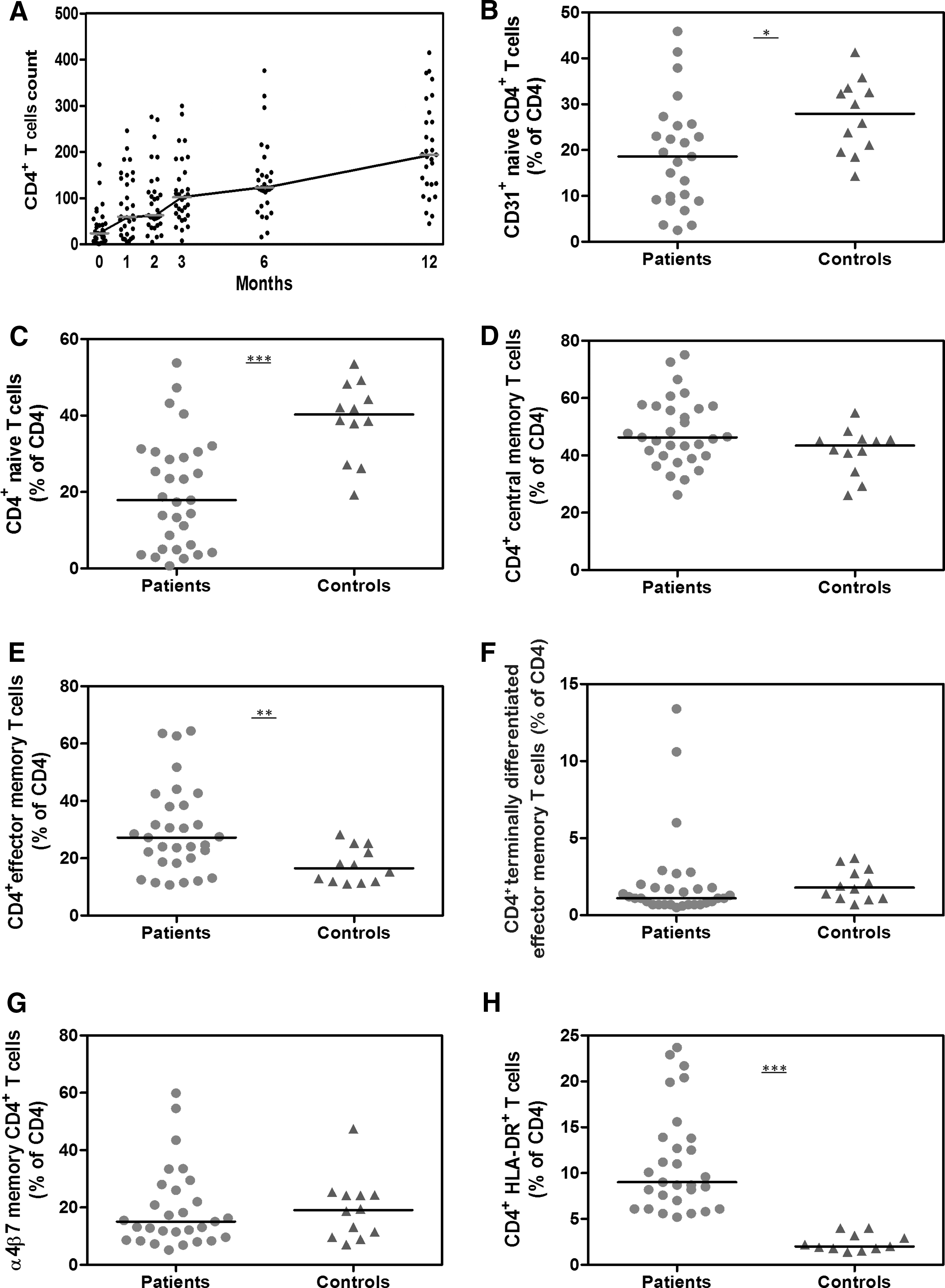
CD4+ T cell subsets and activation after 9 months on antiretroviral therapy (ART). The median (range) CD4+ T cell counts rose on ART
HIV patients have fewer naive CD4+ T cells and greater immune activation than healthy controls, with similar expression of a gut-homing receptor
Following 9 months of ART, HIV patients had lower proportions of total CD4+ T cells than healthy controls [median (range): 8% (2–20) vs. 29% (18–41), respectively; p < 0.0001]. The proportion of CD4+ T cells at 9 months correlated with CD4+ T cell counts at 12 months (r = 0.61, p = 0.0006).
HIV patients had lower proportions of CD31+-naive and naive CD4+ T cells than controls (p = 0.02 and p = 0.0005, respectively; Fig. 2B, C), but higher proportions of EM CD4+ T cells (p = 0.008; Fig. 2E). Proportions of CM and TD CD4+ T cells were similar in patients and controls (p = 0.11 and p = 0.12, respectively; Fig. 2D, F). In HIV patients, proportions of naive CD4+ T cells correlated inversely with proportions of EM CD4+ T cells (r = −0.72, p < 0.0001). CD4+ T cell counts at 12 months correlated directly with proportions of naive (r = 0.74, p < 0.0001) and CD31+-naive CD4+ T cells (r = 0.62, p = 0.001), and inversely with proportions of CM (r = −0.36, p = 0.05) and EM CD4+ T cells (r = −0.36, p = 0.05).
We assessed CD4+ T cells likely to traffic between the peripheral blood and intestinal mucosa through expression of the gut-homing receptor, α4β7. Proportions of α4β7 memory CD4+ T cells were similar in patients and controls (p = 0.71; Fig. 2G), and correlated inversely with proportions of naive CD4+ T cells in HIV patients (r = −0.46, p = 0.01).
The proportion of total CD4+ T cells expressing HLA-DR was greater in patients than controls (p < 0.0001; Fig. 2H). The proportion of total CD4+ T cells expressing HLA-DR correlated inversely with proportions of naive (r = −0.70, p < 0.0001) and CD31+-naive CD4+ T cells (r = −0.51, p = 0.01), and directly with EM (r = 0.68, p < 0.0001) and α4β7 memory CD4+ T cells (r = 0.64, p < 0.0001). Furthermore, inverse correlations were observed between HLA-DR expression on total CD4+ T cells and CD4+ T cell counts at baseline (r = −0.35, p = 0.05) and after 12 months on ART (r = −0.60, p = 0.0007).
Expression of CD127 on CD4+ T cell subsets is directly related to the size of the naive CD4+ T cell pool, and inversely related to immune activation
To assess the capacity of IL-7 to maintain CD4+ T cells in HIV patients, we measured mean fluorescence intensity (MFI) of CD127 staining in 23 HIV patients. Gating strategies for determination of CD127 expression on CD4+ T cell populations are shown in Figure 3A. CD127 expression on total, CM, EM, and TD CD4+ T cells was lower in HIV patients (p < 0.0001, p < 0.0001, p < 0.0001, and p = 0.05, respectively), but CD127 expression on naive CD4+ T cells was similar in patients and controls (Fig. 3B). In HIV patients, CD127 expression on naive CD4+ T cells was higher than on CM (p = 0.0006), EM (p = 0.002), and TD (p < 0.0001) CD4+ T cells. In controls, CD127 expression was higher on CM and EM CD4+ T cells (Fig. 3B).
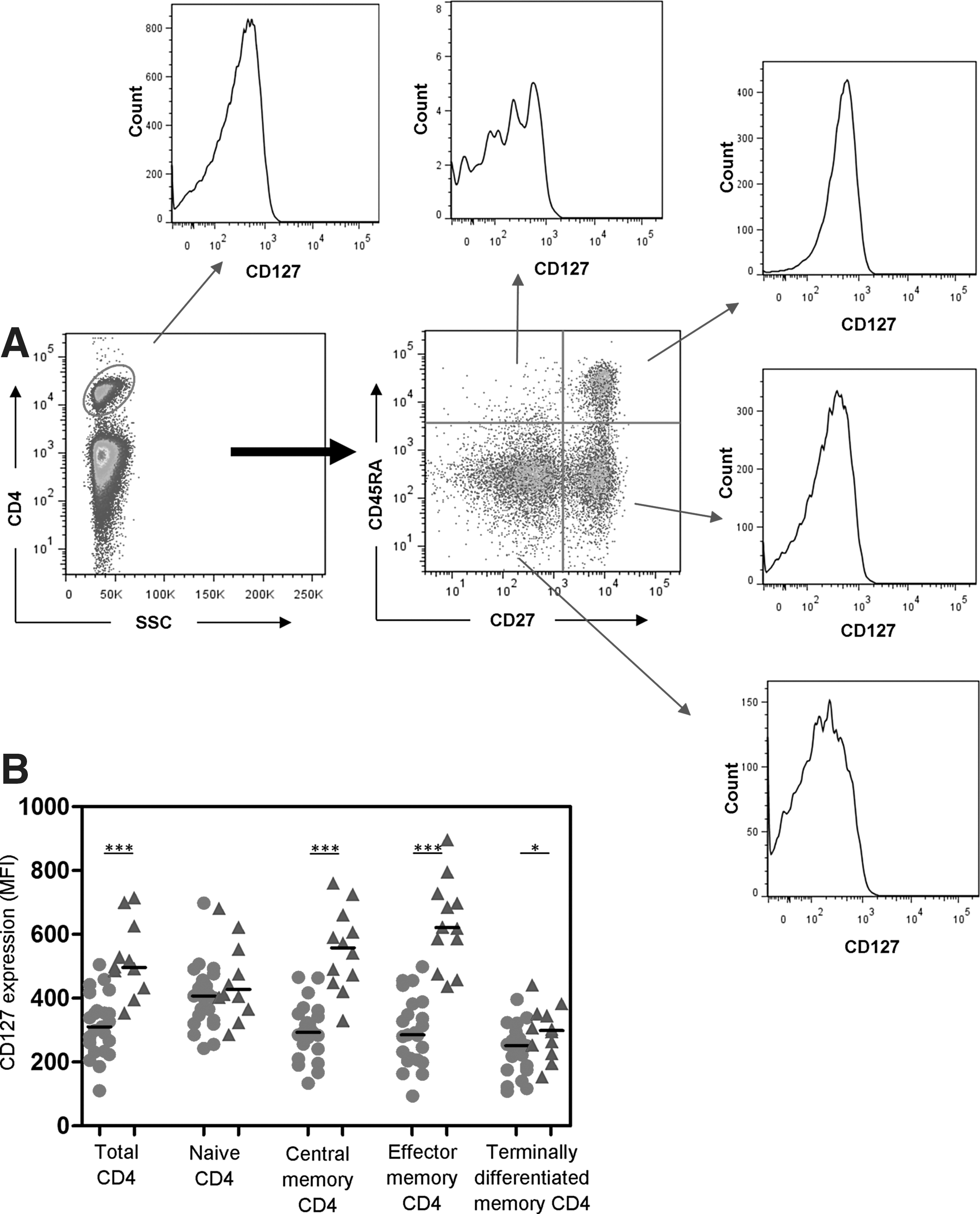
CD4+ T cell expression of CD127.
CD127 expression on total CD4+ T cells correlated directly with CD4+ T cell counts at 12 months (r = 0.55, p = 0.009) and with proportions of naive (r = 0.58, p = 0.004) and CD31+-naive (r = 0.56, p = 0.02) CD4+ T cells. Inverse correlations were observed between CD127 expression on total CD4+ T cells and proportions of EM (r = −0.43, p = 0.04) and TD (r = −0.47, p = 0.02) CD4+ T cells, implicating CD127 in the maintenance of naive T cells.
There were moderate-weak inverse correlations between HLA-DR expression on CD4+ T cells and CD127 on total CD4+ T cells (r = −0.40, p = 0.06) and EM CD4+ T cells (r = −0.43, p = 0.04), highlighting the negative impact of immune activation.
TCR and IL-7 stimulation induce similar proliferative responses in the naive CD4+ T cell subset from HIV patients and healthy controls
PBMC were cultured for 5 days with anti-CD3 antibodies or IL-7. CD4+ T cell proliferation was assessed by subtracting the percentages of CD4+ T cells expressing Ki67 in unstimulated cultures from the percentages expressing Ki67 after stimulation (ΔKi67). The gating strategies for Ki67 expression on CD4+ population is shown in Figure 4A. Low cell numbers precluded analyses of ΔKi67 in terminally differentiated EM CD4+ T cells. IL-7-mediated ΔKi67 values correlated with anti-CD3-mediated ΔKi67 in total, naive, and CM CD4+ T cells ([r = 0.95, p < 0.0001], [r = 0.92, p < 0.0001], and [r = 0.84, p = 0.001], respectively).
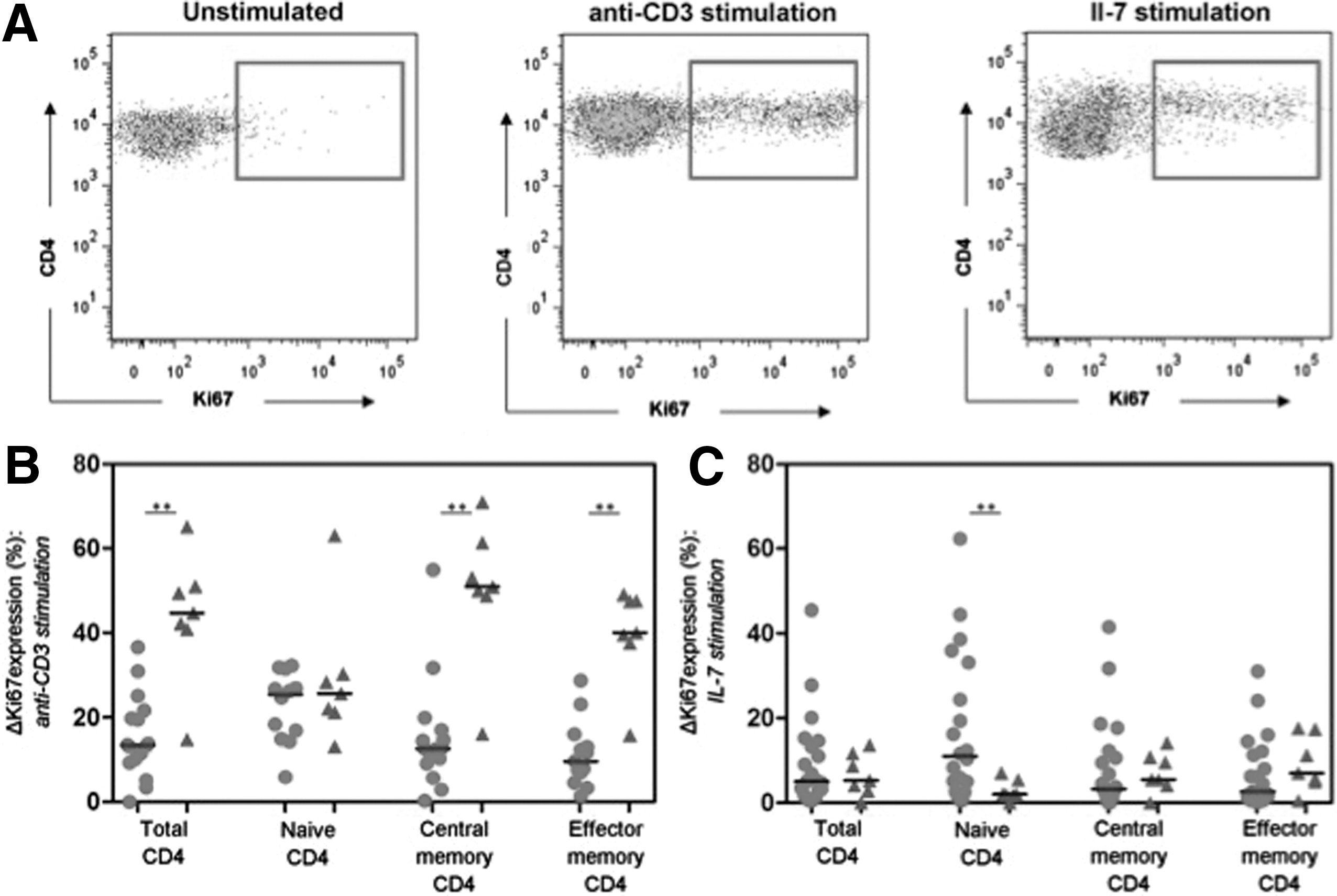
Interleukin (IL)-7 and anti-CD3 mediated CD4+ T cell proliferation. ΔKi67 represents the proportion of T cells expressing Ki67 following stimulation minus that seen without stimulation.
Induction of Ki67 by anti-CD3 antibodies in samples from HIV patients was higher in naive CD4+ T cells than in CM (p = 0.02) or EM (p = 0.004) CD4+ T cells. Conversely in healthy controls, the responses of naive CD4+ T cells were weaker than responses of CM or EM (p = 0.02) CD4+ T cells. Accordingly, Ki67 induction in total, CM, and EM CD4+ T cells was lower in HIV patients than healthy controls (p = 0.009, p = 0.002, and p = 0.0009, respectively), but the ΔKi67 values for naive CD4+ T cells were similar (Fig. 4B).
Similarly, following IL-7 stimulation of cells from HIV patients, ΔKi67 values for naive CD4+ T cells were higher than for CM (p = 0.05) or EM (p = 0.02) CD4+ T cells. In healthy controls, ΔKi67 induction by IL-7 was low in all subsets, but marginally lower in naive CD4+ T cells than in CM (p = 0.09) or EM (p = 0.07) CD4+ T cells. Strikingly, IL-7 induced higher ΔKi67 values in naive CD4+ T cells from HIV patients compared to healthy controls (p = 0.02) (Fig. 4C).
Discussion
In this study, we examine the phenotypes and proliferative capacity of CD4+ T cells in a cohort of severely immunodeficient HIV/HCV-coinfected Indonesian patients commencing ART. These patients are representative of large numbers of patients in Asia, where cotransmission of HCV and HIV through intravenous drug use is common. Most patients were relatively young (median age; 29 years). All had baseline CD4+ T cell counts <200 cells/μL and nearly 80% had CD4+ T cell counts <50 CD4+ T cells/μL. Baseline counts correlated with CD4+ T cell recovery following 12 months of ART. Although most patients (94%) were able to suppress viral replication to undetectable levels, incomplete immune reconstitution (<200 CD4+ T cells/μL) was observed in 52% of patients after 12 months of ART, and just 12% achieved >350 CD4+ T cells/μL.
Studies of patients in Europe and North America (11,27) demonstrate two phases of CD4+ T cell restoration after ART initiation; an initial rapid increase in peripheral CD4+ T cell counts reflecting redistribution of memory T cells from lymphoid tissue, followed by a slower second phase increase reflecting T cells generated through homeostatic expansion and thymic activity. In this study, after 9 months on ART, HIV patients still had lower proportions of total, naive, and CD31+-naive CD4+ T cells, and higher proportions of CD4+ EM T cells, than healthy controls. Proportions of CM, terminally differentiated EM, and α4β7 memory CD4+ T cells were similar to controls.
Thymic function was evaluated by the expression of CD31 on naive T cells (20). The proportions of CD31+-naive CD4+ T cells were low in patients when calculated as a percentage of CD4+ T cells (Fig. 2B). Hence, thymic dysfunction is reason for reduced proportions of naive CD4+ T cells in this cohort. Studies from Europe, North America, and Japan (1,29,30) show that patients who initiate ART with low nadir CD4+ T cell counts retain fewer naive and more memory CD4+ T cells than patients starting with >350 CD4+ T cells/μL. After 3–11 years of effective ART and despite increases in total CD4+ counts, proportions of each functional CD4+ subset may remain perturbed in patients with nadir CD4+ T cell counts <100 cells/μL (30). In such patients, incomplete CD4+ T cell recovery was associated with ongoing CD4+ T cell activation and turnover. The highly differentiated mature CD4+ T cell phenotype predominant in our cohort may affect CD4+ T cell reconstitution as these cells express less CD127 and display poor proliferation following stimulation with anti-CD3.
Despite suppression of viral replication, persistent immune activation may deplete the CD4+ T cell pool (13,16,22) through activation-induced cell death or persistent stimulation of naive T cells to a differentiated phenotype (2). Immune activation can induce fibrosis of lymphoid tissue, increasing apoptosis and reducing numbers of circulating naive CD4+ T cells (31,35). Indonesian HIV patients had higher levels of CD4+ T cell activation (assessed by HLA-DR) than controls, which correlated inversely with CD4+ T cell counts and proportions of total, naive, and CD31+-naive CD4+ T cells. Direct correlations were observed with EM and α4β7 memory CD4+ T cell subsets.
HIV disease progression is associated with low expression of CD127, particularly on memory CD4+ T cells (21,28). In this study, HIV patients had reduced expression of CD127 on all CD4+ T cell subsets, except for naive T cells. Expression of CD127 was highest on naive CD4+ T cells in HIV patients, but highest on EM cells in controls. This has been observed in less immunodeficient patients treated in the USA (5) and attributed to modulation of CD127 expression in response to increased IL-7 or the accumulation of CD127− EM T cells (21). This is pertinent in this study as naive CD4+ T cells were selectively decreased in HIV patients. Increased CD127 expression may be a compensatory mechanism, priming naive T cells to be highly responsive to IL-7.
In HIV patients, low expression of CD127 on CD4+ T cells has been correlated with T cell activation (HLA-DR or CD38 expression), decreased expression of antiapoptotic factors (such as Bcl-2 and Bcl-xl), and increased T cell apoptosis (7,14,21). We confirmed the link between CD127 expression and T cell activation (19); so activation-induced loss of CD127 expression may contribute to ongoing CD4+ T cell deficiency, despite virologically suppressive ART.
We compared the proliferative capacity of various CD4+ T cell subsets in response to anti-CD3 (mimicking T cell activation) or IL-7 (reflecting homeostatic proliferation). Anti-CD3 was more generally effective at inducing proliferation than IL-7, but naive CD4+ cells in HIV patients responded equally to both stimuli. Responses of naive CD4+ T cells to anti-CD3 were similar in patients and controls as shown previously (5). Responses to IL-7 stimulation were higher in naive CD4+ T cells from patients than those from controls; so the paucity of naive cells in the HIV patients cannot be attributed to their inability to maintain themselves in the periphery by homeostatic proliferation.
In conclusion, despite suppression of viral replication and increased CD4+ T cell counts, proportions of each T-cell subset did not normalize following 9 months on ART and T cell activation remained higher than in control donors living in the same environment. It appears that persistent immune activation, antigen-driven stimulation, and differentiation of naive T cells (rather than thymic insufficiency) depletes the naive T cell pool and drives the accumulation of highly activated and differentiated short-lived memory T cells with reduced proliferative capacity. Preservation of a functional naive T cell compartment is critical for effective immune responses to newly encountered pathogens, malignancies, and vaccinations. Further studies (in this or a similar Asian cohort) at a later time point (1–5 years post-ART) are needed to assess the long-term effects of immune reconstitution on CD4+ T cell subset proportions and function in this environment.
Footnotes
Acknowledgments
The authors thank all patients who participated in this study, Dr. Agus Kosasih (Clinical Pathology Laboratory at Dharmais National Cancer Center) for access to the flow cytometer, Dr. Budiman Bela for laboratory facilities, and Azmier Adib Arin for assistance in the laboratory. The work was supported by the Strategic Research Scheme of the University of Indonesia and the Goodeve Foundation (Western Australia). S.F. is supported by the Raine Medical Research Foundation (Australia).
Authors' Contributions
E.Y. and R.A.G. recruited patients and arranged sample collection. H.S. archived samples. S.T. performed the flow cytometry and analyzed the data with guidance from S.F. P.P. and S.D. coordinated the project.
Author Disclosure Statement
No competing financial interests exist.