
Abstract
Human enterovirus 71 (EV-A71), a major agent of hand, foot, and mouth disease, has become an important public health issue in recent years. No effective antiviral or vaccines against EV-A71 infection are currently available. EV-A71 infection intrudes bodies through the gastric mucosal surface and it is necessary to enhance mucosal immune response to protect children from these pathogens. Recently, the majority of EV-A71 vaccine candidates have been developed for parenteral immunization. However, parenteral vaccine candidates often induce poor mucosal responses. On the other hand, oral vaccines could induce effective mucosal and systemic immunity, and could be easily and safely administered. Thus, proper oral vaccines have attached more interest compared with parenteral vaccine. In this study, the major immunogenic capsid protein of EV-A71 was displayed on the surface of Saccharomyces cerevisiae. Oral immunization of mice with surface-displayed VP1 S. cerevisiae induced systemic humoral and mucosal immune responses, including virus-neutralizing titers, VP1-specific antibody, and the induction of Th1 immune responses in the spleen. Furthermore, oral immunization of mother mice with surface-displayed VP1 S. cerevisiae conferred protection to neonatal mice against the lethal EV-A71 infection. Furthermore, we observed that multiple boost immunization as well as higher immunization dosage could induce higher EV-A71-specific immune response. Our results demonstrated that surface-displayed VP1 S. cerevisiae could be used as potential oral vaccine against EV-A71 infection.
Introduction
E
Given its safe and easy administration, oral immunization is a practical and effective way to prevent diseases caused by enteric pathogens in children from developing countries (2,15,21,22). The mucosal immune system is important in controlling viral infections because EV-A71 infection is initiated at mucosal sites (15). Thus, proper EV-A71 oral vaccines have been attached more interest compared with parenteral vaccine. Many vaccine candidates used for oral immunization, including recombinant VP1 protein, which is produced in tomato (2), Lactococcus lactis (23,27) and baculovirus (14), have demonstrated that the VP1 protein of EV-A71 can act as an antigen to be used for making an oral vaccine against EV-A71 infection.
Saccharomyces cerevisiae is a safe and food-grade yeast that has been used for the oral application of vaccines and drugs. Many heterologous proteins have been previously expressed on the surface of yeast cells to develop possible oral vaccines. These proteins include ApxIIA exotoxin from Actinobacillus pleuropneumoniae (9), porcine epidemic diarrhea virus spike protein (13), and EtMic2 protein from Eimeria tenella (20). In addition, the first commercialized recombinant human vaccine, the hepatitis B vaccine, was produced in S. cerevisiae (7). Therefore, we expressed the VP1 protein of EV-A71 displayed on the surface of yeast cells and evaluated it as a potential oral vaccine against EV-A71 infection in a mouse experimental model in the present study.
Materials and Methods
Ethics statement
All experiments with mice were approved by the Institutional Animal Care and Use Committee of the Guangzhou Institutes of Biomedicine and Health, Chinese Academy of Sciences (Permit No.: 2014005). All animals were housed individually and received humane care. During injection and sample collection, the mice were anesthetized with 1.5% isoflurane to minimize suffering.
Viruses and antibodies
EV-A71 (GZ-08-02 strain, Accession No. FJ360545) was isolated and propagated with Vero cells in Dulbecco's modified Eagle's medium (DMEM; Gibco) supplemented with 2% fetal bovine serum (FBS) (30). EV-A71 virus stock was collected and tittered in Vero cells. The anti-VP1 monoclonal antibody 5C3 and 5A10 were homemade; 22A12 was purchased from AbMax Biotechnology Co., Ltd.
Vector construction
The vector pYD1 was purchased from Invitrogen Life Technology (25). The VP1 gene was amplified from EV-A71 cDNA using polymerase chain reaction (PCR). The vector pYD1-VP1 was constructed by inserting the VP1 gene with 6× His amino-terminal tag into plasmid pYD1 cloning sites at the EcoRI and XhoI sites by standard molecular techniques.
S. cerevisiae transformation and identification
The plasmids pYD1 and pYD1-VP1 were transformed into S. cerevisiae EBY100-competent cells according to the manufacturer's instructions (25). The transformed yeast cells were plated on minimal dextrose plates (0.67% yeast nitrogen base, 2.0% glucose, 0.01% leucine, and 1.5% agar) at 30°C for 3 days. The transformants that contain pYD1 and pYD1-VP1 were identified by colony PCR. Positive colonies of the transformed yeast were named EBY100/pYD1 and EBY100/pYD1-VP1, respectively.
VP1 gene expression and western blot analysis
Yeast cells were cultured in YNB-CAA medium (0.67% yeast nitrogen base, 0.5% Casamino acids, and 2.0% galactose) at 20°C for 48 h. The cells were harvested at different time points (0, 12, 24, 36, and 48 h) by centrifugation at 5,000×g for 10 min and washed three times with phosphate-buffered saline (PBS) buffer at 4°C. The cell pellets of the recombinant S. cerevisiae EBY100 were analyzed by 12% SDS-PAGE and then transferred onto a PVDF membrane. After blocking, the membrane was incubated for 1.5 h at room temperature with anti-VP1 antibody (5C3, 1:300) to detect Aga2p-fused VP1 protein (30). In addition, the membrane of the yeast cells harvested at 36 h was incubated with three kinds of mouse anti-VP1 monoclonal antibodies (5C3, 5A10, 22A12). After washing in PBS, the membrane was incubated with horseradish peroxidase (HRP)-conjugated goat anti-mouse antibodies (1:5,000; Thermo Scientific) for 1 h at room temperature. The protein bands were detected by the Immobilon ECL Plus Western Blotting Detection Reagents (Millipore).
Confocal microscopy to detect expression of VP1 in yeast cells
Yeast cells were harvested and washed three times with PBS buffer at 4°C, then they were incubated with mouse anti-VP1 antibody (5C3, 1:300) at room temperature for 2 h to detect Aga2p-fused VP1 protein, washed five times with PBS buffer, and then incubated with Alexa Fluor 488-labeled goat anti-mouse IgG (IgG, 1:2,000; Life Technologies) as a secondary antibody for 1 h at room temperature. The Alexa Fluor 488-labeled cells were washed five times with PBS and then analyzed using a Zeiss 710 NLO two-photon laser scanning confocal microscope (Carl Zeiss) with an excitation wavelength of 488 nm and an emission wavelength of 514 nm.
Immunization of female BALB/c mice and sample collection
Adult female BALB/c mice aged 6 weeks were immunized orally with 5 × 109 cells/mL (0.5 mL per mouse) of VP1-expressing S. cerevisiae (pYD1-VP1 [500 μL] group). To evaluate whether oral immunization with surface-displayed VP1 S. cerevisiae have a dose-dependent effect in vaccinated mice, one control group was immunized orally with half dose (0.25 mL per mouse) of VP1-expressing S. cerevisiae (pYD1-VP1 [250 μL] group). Before immunization, all mice were fed with saturated sodium bicarbonate for 2 h to protect the antigens from the gastric acid. The negative control group was immunized orally with pYD1-transformed S. cerevisiae (pYD1 group). For immune comparison, the animals in another group were injected intraperitoneally with formalin-inactivated EV-A71 virus (inactivated EV-A71 group). Each mouse in the inactivated EV-A71 group was injected intraperitoneally with inactivated EV-A71 virus (0.2 mL per mouse, 107 TCID50). For further study, one group of mice was orally treated with the inactivated EV-A71 (Oral EV-A71 group, 0.2 mL per mouse, 107 TCID50). All groups were immunized with the same dose as a booster at 2 and 4 weeks postimmunization.
Blood, vaginal fluid, and saliva samples were collected from each mouse at 0, 2, 4, and 6 weeks after the primary immunization. Blood samples (0.5 mL) from each mouse were collected from the orbital artery and concentrated at 3,000 rpm for 10 min. The sera were inactivated at 56°C for 30 min and then stored at −80°C until ELISA and neutralization assays. Saliva samples were induced by carbachol (130 μL of 0.01 mg/mL) (Sigma-Aldrich) (10,28). Vaginal samples were collected by washing the vagina two times with 100 μL of sterile PBS (10,28). Saliva and vaginal fluid were centrifuged at 8,000 g for 20 min and supernatant was collected and stored at −20°C for further analysis. Six mice of each group were euthanized and bled at week 8. Then, the spleens of these mice were removed and homogenized aseptically as described below. In addition, lung lavage fluid and intestinal lavage fluid of these mice were collected and processed as reported (11,18).
Specific EV-A71 VP1 antibody determination by ELISA
The levels of VP1-specific serum IgG, IgM, and mucosal IgA were determined separately by indirect enzyme-linked immunosorbent assay (ELISA). Briefly, 96-well plates were coated overnight with 2 μg/mL purified E. coli-expressed VP1 protein dissolved in carbonate coating buffer (15 mM Na2CO3, 35 mM NaHCO3, pH 9.6) at 4°C. The plates were subsequently blocked with PBS containing 10% FBS at 37°C for 1 h. Diluted serum (1:500) or mucosal secretion samples (1:5) were added into wells (100 μL per well) at 37°C for 1.5 h. After washing five times with PBST (PBS with 0.5% Tween-20), HRP-conjugated goat anti-mouse IgG (1:4,000; Pierce), HRP-conjugated goat anti-mouse IgM (1:2,000; Bioss) or HRP-conjugated goat anti-mouse IgA (1:8,000; SouthernBiotech) was added into the plate to detect IgG, IgM, and IgA antibodies, respectively, in serum and mucosal secretion samples. The plates were washed as previously described and then added with 100 μL of tetramethylbenzidine (TMB) (Millipore) at 37°C for 20 min. The reaction was stopped by adding 50 μL of H2SO4 (2 mol/L) and measured at 450 nm.
A similar ELISA analysis was conducted to measure levels of IgA antibody in newborn mice. Adult female BALB/c mice were divided into five groups: pYD1, pYD1-VP1 (250 μL), pYD1-VP1 (500 μL), Inactivated EV-A71, and Oral EV-A71 group. These animals received immunization as previously described and they were allowed to mate 2 weeks later after primary immunization. Then, six neonatal mice born to each mother were dissected 24 h after birth and their small intestines were collected and processed as previously reported (12). IgA titers were measured by ELISA as described above.
Serum neutralization assay
The neutralization test was carried out as previously reported (3,27). Mouse sera with a two-fold serial dilution from 1:4 to 1:128 were mixed with equal volumes of 100 TCID50 EV-A71 at 37°C for 2 h. The mixture was added to monolayers of Vero cells (1.5 × 104 cells/well) in 96-well plates. After 5 days of incubation at 37°C, the neutralizing antibody titers were read as the highest dilution of serum that resulted in less than 50% cytopathic effect (CPE).
ELISA for quantifying TNF-α production
The spleens of the immunized mice at 56 days after the third immunization were isolated as previously reported (24). Lymphocytes were separated by an EZ-Sep™ Mouse Lymphocyte Separation Kit (Dakewe). Suspensions of splenic cells were seeded in a 48-well plate at a density of 2 × 106 cells per well and subsequently stimulated with purified VP1 protein (5 μg/mL), Con A (5 μg/mL; Sigma), and medium alone for 72 h. The supernatants were collected, and tumor necrosis factor (TNF)-α production was measured by using a mouse TNF-α ELISA Kit (Dakewe) following the manufacturer's instructions.
Enzyme-linked immunospot assay for quantifying IFN-γ production
The levels of interferon (IFN)-γ were determined by using the ELISPOT Kit (Mabtech) following the manufacturer's protocol. A 96-well filtration plate (Nalge Nunc) was coated with 5 μg/mL rat anti-mouse IFN-γ capture Ab (Mabtech). After overnight incubation at 4°C, the plate was washed with PBS, blocked with complete RPMI medium (10% FBS, 2 mM glutamine, 1 mM pyruvate, 10 mM HEPES, 100 U/mL penicillin, 100 U/mL streptomycin, and 55 mM mercaptoethanol) at 37°C for 2 to 4 h, and then washed again. The splenocytes resuspended in complete RPMI medium were seeded in the plates (each well with 1 × 106 cells) and subsequently stimulated with purified VP1 protein (5 μg/mL), Con A (5 μg/mL; Sigma), and medium alone for 24 h at 37°C. The plate was washed with PBST, added with biotin-conjugated rat anti-mouse IFN-γ antibody R4-6A2 (Mabtech) at 2 μg/mL in PBS, and then incubated overnight at 4°C. Subsequently, the plate was incubated with streptavidin-HRP at 37°C for 2 h. Finally, the plate was visualized by adding 100 μL/well of TMB substrate solution (Mabtech). The number of spots was counted by using the enzyme-linked immunospot (ELISPOT) reader.
Virus challenge assay
Virus-challenged experiments were performed as previously described (5). Adult female BALB/c mice were divided into three groups: pYD1, pYD1-VP1, and inactivated EV-A71 group. The mice received primary immunization and then were allowed to mate 2 weeks later. All mice received booster immunization after pregnancy at weeks 2 and 4. Subsequently, the newborn mice were challenged i.p. with EV-A71 strain virus (500 LD50, 100 μL/mouse) 24 h after birth. The mortality rate of the mice was recorded at 21 days after virus challenge.
Results
S. cerevisiae surface expression of VP1 protein
In the constructed pYD1-VP1 plasmid, the VP1 gene was fused with the Aga2 gene to allow secretion and display of VP1 protein (Fig. 1A). The yeast cells were collected at different times points for western blot analysis with three kinds of anti-VP1 monoclonal antibody, which showed that the highest expression level was recorded from 24 to 36 h (Fig. 1B). The results of western blot assay with three kinds of anti-VP1 monoclonal antibodies confirmed the yeast cells transformed with the pYD1-VP1 plasmid expressed Aga2p-VP1 fusion proteins (Fig. 1C). The confocal microscopy results revealed that VP1 fusion proteins were directed to the surface of the yeast cells (Fig. 1D). By contrast, no immunofluorescence was observed in the cells transferred with the pYD1 plasmid. These results demonstrated that VP1 protein successfully expressed on the surface of EBY100 cells.

Construction of the vector pYD1-VP1 and expression VP1 in vitro.
Humoral responses elicited by oral immunization with surface-displayed VP1 S. cerevisiae
To explore whether the surface-displayed VP1 S. cerevisiae could serve as an EV-A71 immunogen in mice, female BALB/c mice were immunized three times at weeks 0, 2, and 4 with EBY100/pYD1, EBY100/pYD1-VP1, and inactivated EV-A71. The sera and mucosal secretion samples were collected at 0, 2, 4, and 6 weeks after pre- and postimmunization (Fig. 2A). The titers of specific anti-VP1 IgG and IgM antibody in the sera of immunized mice were analyzed by ELISA. Immunization with inactivated EV-A71 induced high levels of IgG, but low-to-moderate levels of IgG were induced by EBY100/pYD1-VP1 at 2 weeks after the primary immunization. Titer of IgG was enhanced after booster immunization (Fig. 2B). The IgM titer significantly increased in the pYD1-VP1 and inactivated EV-A71 groups compared with the control mice at 2 weeks after the primary immunization (p < 0.05). At weeks 4 and 6, the IgM titers remarkably increased in the pYD1-VP1 group, but decreased in the inactivated EV-A71 group (p < 0.05, Fig. 2C).
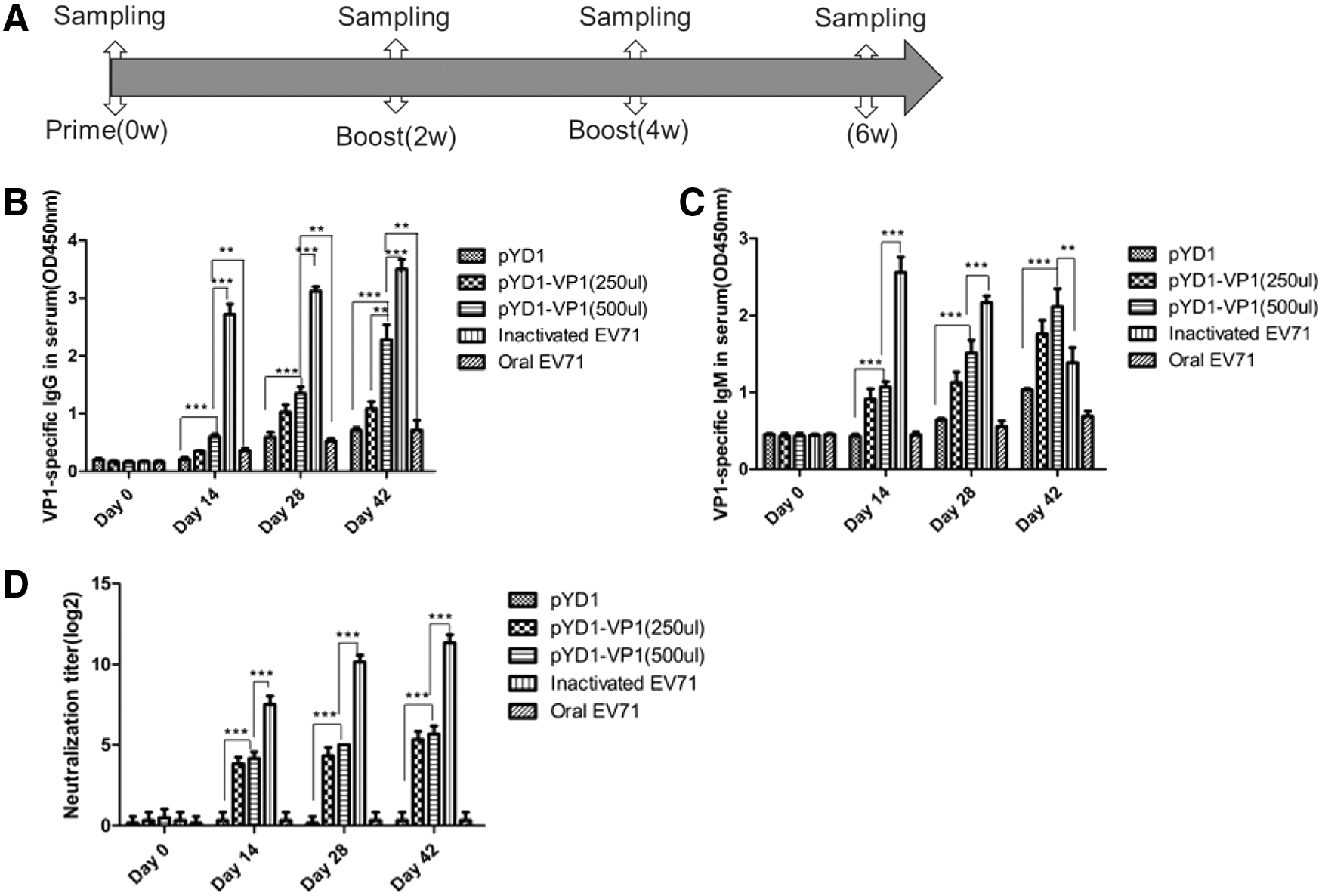
Humoral immune responses induced by surface-displayed VP1 S. cerevisiae.
Serum samples were evaluated further to determine the EV-A71 neutralization in vitro by serum neutralization assay. The results of neutralization assay showed that the mice immunized with inactivated EV-A71 induced higher neutralizing antibody titers than the control mice (p < 0.001, Fig. 2D). After multiple doses, low-to-moderate levels of neutralizing antibody were elicited by EBY100/pYD1-VP1 immunization. The neutralization titer induced by EBY100/pYD1-VP1 was 26, whereas that induced by inactivated EV-A71 was 211.
To assess the effect of the surface-displayed VP1 S. cerevisiae immunization on mucosal immune response, vaginal fluid and salivary IgA antibody response were tested by ELISA. IgA titers in vaginal fluid and saliva of mice immunized with EBY100/pYD1-VP1 were higher than that of mice immunized with EBY100/pYD1 or inactivated EV-A71 (Fig. 3A, B), the difference was also observed with the titer of IgA in the irrigating solution of intestine and lung (Fig. 3C, D). The levels of IgA antibody were enhanced after booster immunization in vaginal fluid and saliva. There was no statistically significant difference in the IgA titer between the pYD1 and inactivated EV-A71 group mice in the irrigating solution of intestine and lung.

Mucosa immune responses induced by surface-displayed VP1 S. cerevisiae.
Overall, IgG, IgM, IgA, and neutralization titers were lower in the pYD1 and oral EV-A71 groups. In addition, a dose-dependent ELISA IgG, IgM, IgA, and neutralization titers antibody response was observed in pYD1-VP1 (250 μL) and pYD1-VP1 (500 μL) groups.
Th1 cytokine production in mice orally immunized with surface-displayed VP1 S. cerevisiae
We analyzed the cytokines from the splenocytes of the immunized mice by ELISA and ELISPOT to evaluate the Th1 cytokine production elicited by oral immunization with surface-displayed VP1 S. cerevisiae. The concentrations of TNF-α in the culture media of the splenocytes after VP1 protein purification from E. coli stimulation were measured by ELISA. Splenic cells from immunized mice were also stimulated with the purified VP1 protein and assayed for IFN-γ by ELISPOT. The levels of TNF-α and IFN-γ were significantly higher in the EBY100/pYD1-VP1 and inactivated EV-A71 groups than in the pYD1 group (Fig. 4). These results indicated that both EBY100/pYD1-VP1 (oral administration) and inactivated EV-A71 (injection) can elicit Th1 cellular immune responses. Furthermore, the pYD1-VP1 group produced higher levels of TNF-α than the inactivated EV-A71 group.
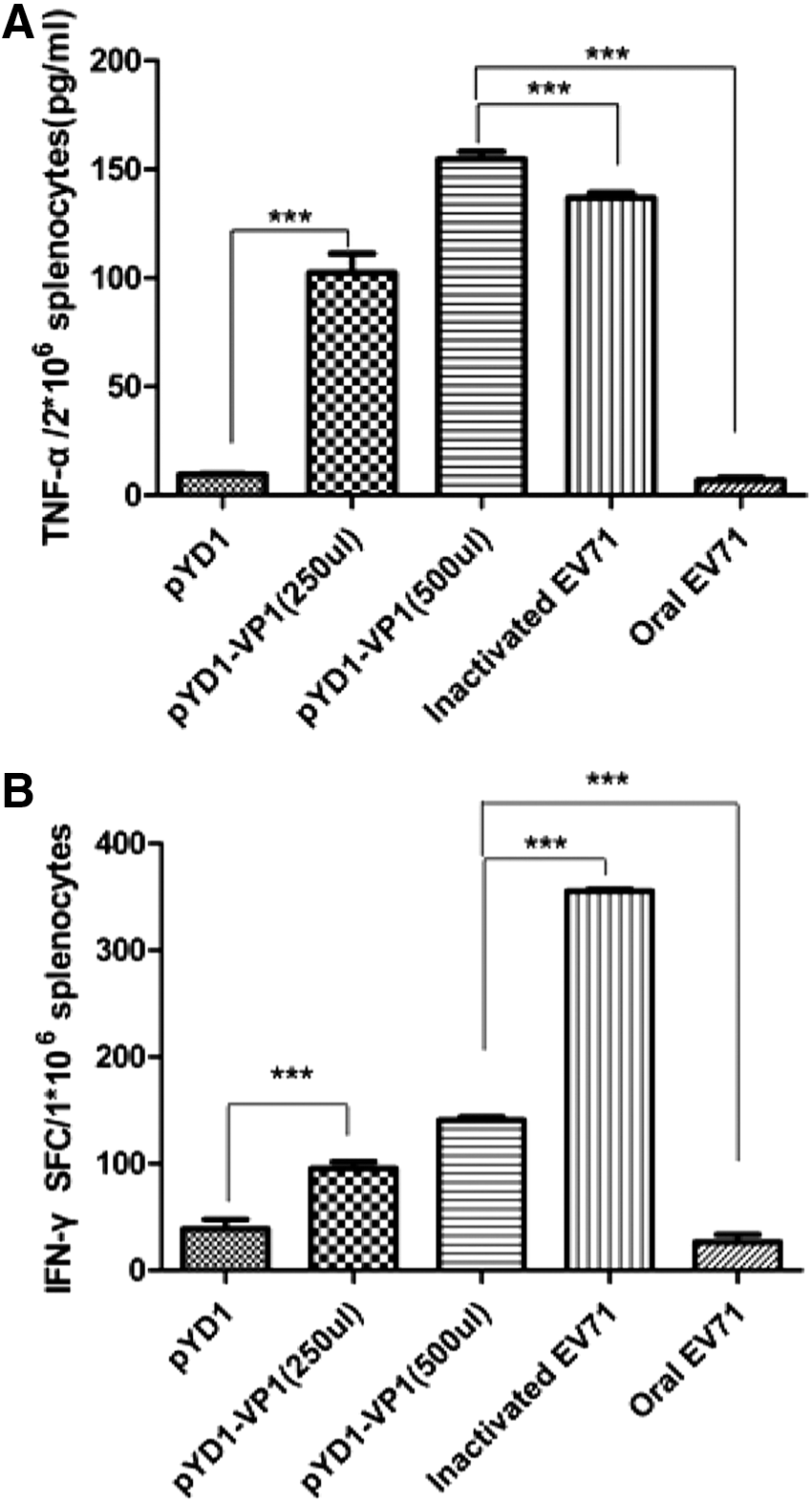
Th1 cytokine production in vaccinated mice.
Protection against lethal EV-A71 challenge in suckling mice
The suckling mice were infected with a lethal dose of EV-A71 to evaluate whether oral immunization with surface-displayed VP1 S. cerevisiae could confer some protection from the mother mice to the newborn mice. At the EV-A71 challenge dose of 500LD50, the survival rates in the pYD1-VP1 and inactivated EV-A71 groups at day 21 were 60% and 100% (n = 10 each group), respectively. By contrast, all mice in the control group died by day 9 after lethal EV-A71 infection (Table 1). On the other hand, intestinal IgA concentration of neonatal mice was higher than that of control mice (Fig. 3C). These results indicated that oral immunization with S. cerevisiae expressing VP1 can confer some protection from the mother mice to the neonatal mice.
Versus pYD1-VP1 group, p < 0.01.
Versus pYD1-VP1 group, p > 0.05.
Discussion
HFMD caused by EV-A71 has become a major emerging infectious disease that presents serious public health problems in Southeast Asia (1). Present EV-A71 vaccine candidates have been developed as inactivated EV-A71 virion or live attenuated virus vaccines for parenteral immunization. Parenteral immunization often involves laborious and time-consuming procedures, produces inflammatory reactions at the injection site, and creates stress in children (22). Moreover, these conventional vaccine candidates require adjuvants to induce sufficient immunity and generally do not induce high levels of secretory immunoglobulin A (IgA) (20).
Recently, oral vaccines have received increasing attention because of their advantages over conventional vaccines, nonpathogenicity to humans, and simple administration. Moreover, oral vaccines could induce effective mucosal or systemic immunity, and could be easily and safely administered to a large number of people in a short period of time (21). Owing to the lack of proper mucosal adjuvants, oral immunization in humans is limited. Many studies have demonstrated that the components of yeast cell wall function as adjuvants that enhance the immune response and cannot be detected as the antibody against itself (29), reducing the possibility of an yeast cell to cause immune rejection (6,19). In the present study, we expressed the VP1 protein of EV-A71 on the cell surface of S. cerevisiae as live oral vaccine to immunize mice against EV-A71 challenge. Compared with traditional vaccines, this novel oral vaccine does not need to add adjuvants. At the same time, one of the advantages of the recombinant yeast vaccines combined with other subunit vaccine is the production of sustainable and high-level antibody (16).
There is an urgent need to strengthen mucosal immunity to protect children from EV-A71 infection starting from invading mucosal surfaces. Oral administration seems to be the main option for mucosal vaccination (14). The major mucosal Ab is IgA present in various mucosal and exocrine fluids. IgA is an antibody that plays a critical role in mucosal immunity, which serves to protect the mucosal surfaces from disease by inhibiting adherence of virus to epithelial surfaces and binding virus in the lamina propria or inside epithelial cells (8). In the present study, VP1-specific IgA antibody in mucosal secretion samples were significantly increased in mice fed with the surface-displayed VP1-expressing yeast compared with the mice immunized with EBY100/pYD1 or inactivated EV-A71. In passive immunization study, VP1-specific IgA antibody of newborn mice is also increased. The results indicated that the mice oral immunization with recombinant yeast can induce specific mucosal immune response effectively. At the same time, we also find that this recombinant S. cerevisiae could induce higher levels of VP1-specific IgG, IgM, and neutralization antibodies in humoral immune responses. Compared with traditional vaccines (killed or attenuated pathogens), oral vaccination with VP1-expressing S. cerevisiae can induce both local mucosal immune responses and systemic immune responses in the present study. More encouraging, our study suggested that multiple boost immunization as well as higher immunization dosage could efficiently enhance EV-A71-specific immune response. Being a live yeast-based oral vaccine with very limited requirement for purification and formulation, high-dose multiple immunizations are easy to achieve. Importantly, the VP1-expressing S. cerevisiae was generally safer than the inactivated virus vaccine.
Furthermore, we confirmed the induction of VP1-specific Th1 response by immunization with surface-displayed VP1-expressing yeast. During viral clearance, cellular immunity plays a central role in eliminating the EV-A71 virus (21). In previous reports, TNF-α and IFN-γ have been shown to independently inhibit the replication of a variety of viruses. Thus, the levels of TNF-α and IFN-γ are two significant indicators to assess the effect of the vaccine candidates. We observed that the levels of TNF-α and IFN-γ, considered as Th1 cytokines, were significantly increased in the splenocytes isolated from the mice vaccinated with the surface-displayed VP1-expressing yeast compared with that isolated from pYD1 control mice. These results proved that surface-displayed VP1 S. cerevisiae can evoke mucosal and systemic immune responses in vivo.
The passive immunization study indicated that recombinant S. cerevisiae as an oral vaccination can confer some protection from the mother mice to the neonatal mice against EV-A71 infection. In other words, immunization of pregnant mothers with this recombinant S. cerevisiae may provide consequent and passive transplacental transmission of antibodies to the fetus to protect neonates. The challenge tests in this study indicated that the recombinant S. cerevisiae was able to induce strong protective immune responses against EV-A71 infection.
In summary, we developed recombinant S. cerevisiae displaying VP1 protein for use as an oral vaccine against EV-A71 infection, and confirmed that this novel vaccine could induce both humoral and cellular immunity in a mouse experimental model. This novel oral vaccine utilized in S. cerevisiae expressing VP1 protein has its own advantages compared with the use of inactivated EV-A71 virus. First, oral immunization is easily accepted in children who need an EV-A71 vaccine from developing countries. Second, S. cerevisiae is a safe and food-grade yeast with its easy administration at any time and any place not only in hospitals. In addition, further studies are required to assess the optimal dose of the recombinant S. cerevisiae fed to pregnant mothers and infants that can protect children against EV-A71 infection.
Footnotes
Acknowledgment
This work was supported by grants from Guangdong Key Science & Technology Project (Grant Nos. 2011A080502006 and 2012A080202007).
Author Disclosure Statement
No competing financial interests exist.