
Abstract
Avian influenza virus (AIV), a mucosal pathogen, gains entry into host chickens through respiratory and gastrointestinal routes. Most commercial AIV vaccines for poultry consist of inactivated, whole virus with adjuvant, delivered by parenteral administration. Recent advances in vaccine development have led to the application of nanoparticle emulsion delivery systems, such as poly (
Introduction
A
The ability to identify an efficacious mucosal adjuvant that elicits an antibody response when administered with an inactivated virus by the aerosol route could be advantageous for improved safety and for mass administration. This approach, however, has not been successfully employed in chickens, although a previous attempt was made to investigate the immunogenicity of whole, inactivated H9N2 AIV delivered by single aerosol administration with aluminum hydroxide, chitosan, cholera toxin B subunit, and Stimune (9). Recently, the ability of aerosolized H9N2 AIV to cause infection in the respiratory tract of chickens has been demonstrated (11), further emphasizing the need to explore the potential of aerosol delivery.
Adjuvants play a critical role in enhancing vaccine efficacy by improving antigen-specific immune responses through modulation of the magnitude and type of adaptive immune responses generated (6). Pathogen-associated molecular patterns, ligands for pattern recognition receptors (PRRs), are considered to be adjuvants since activation of cells of the innate immune system is a prerequisite for induction of an adaptive immune response (29).
Toll-like receptor (TLR) ligands have been established as immunostimulatory in several species, including chickens (24) and, therefore, can be exploited to enhance immune responses to AIV vaccines in the capacity of prophylactic administration (27) or as vaccine adjuvants (12,23,25). TLR 21 is regarded as the PRRs for CpG ODN in chickens (16). Recent evidence suggests that TLR15 is significant in the recognition of CpG ODN by chicken macrophages (5); however, the implication of this finding is currently unknown.
The evolution of vaccine development has resulted in increased emphasis on application of novel adjuvants like CpG ODNs and particulates such as polymeric nanoparticles [for example, poly (
The immunogenicity of a formalin-inactivated H9N2 virus administered intramuscularly with CpG ODN 2007 has also been shown in chickens (22). A previous study by Mallick et al. (19) has also demonstrated the induction of enhanced immunogenicity (antibody-mediated and cell-mediated) of an H4N6 virosome-based vaccine containing CpG ODN as an adjuvant, delivered subcutaneously in chickens.
Nanoparticles, particulate materials ranging from 1–1000 nm in size, possess features of biodegradability and biocompatibility and can function as delivery systems and immunostimulants (32). Nanoparticle emulsion-containing vaccines confer advantages such as enhanced immunogenicity, prevention of degradation of encapsulated contents, persistence and prolonged release (depot effect) at the site of administration, facilitating targeted delivery to antigen-presenting cells (APCs) with uptake by endocytosis, and an overall dose-sparing effect (15,32).
Noncondensing synthetic polymers such as PLGA are particularly useful for targeting APCs due to their neutral or slightly negative charge (2). The size of nebulized nanoparticles can be optimized for deposition at specific sites along the respiratory tract, maximizing the possibility of interaction with APCs. Small particles in the range of 1–3 μm have been shown to have a more homogeneous distribution in the respiratory tract of chickens (7).
The immunogenicity of these novel adjuvants and delivery systems in combination with inactivated AIV in chickens has been investigated to a limited degree, particularly in the context of different routes of administration. We have previously demonstrated the adjuvant potential of CpG ODN 2007 (CpG 2007) at a dose of 2 μg, in combination with a formalin-inactivated H9N2 virus delivered intramuscularly (22). Therefore, the present study was aimed at comparing the adjuvant potential of nonencapsulated CpG 2007 (2 μg) with PLGA-encapsulated CpG 2007 (2 μg) in combination with a formalin-inactivated H9N2 virus using intramuscular and aerosol routes of administration.
Materials and Methods
Chickens
Specific pathogen-free (SPF) chickens were acquired from the Canadian Food Inspection Agency. All animal procedures were approved by the University of Guelph Animal Care Committee and conducted under strict compliance with the guidelines of the Canadian Council on Animal Care.
CpG 2007 and non-CpG 2007 sequences
CpG 2007 containing a modified phosphorothioate backbone (5′-TCGTCGTTGTCGTTTTGTCGTT-3′) and non-CpG 2007 (5′-TGCTGCTTGTGCTTTTGTGCTT-3′) were purchased from Sigma-Aldrich Canada and reconstituted in sterile endotoxin-free water.
AIV strain and immunization studies
Formalin (final concentration 0.02%)-inactivated H9N2 virus (A/Turkey/Ontario/1/66) was prepared as described previously (22). Ninety-six, 1-day-old SPF White Leghorn chickens were randomly allocated to 16 treatment groups with each treatment group containing a total of 6 chickens. Chickens in groups 1–8 were vaccinated intramuscularly, while those in groups 9–16 were vaccinated through the aerosol route using the same vaccine formulations as shown in Table 1. Primary vaccination was performed on day 7 posthatch, while secondary vaccination occurred on day 21 posthatch. In the case of aerosol administration, a third vaccination was carried out on day 35 posthatch. Each chicken received 15 μg of formalin-inactivated virus with or without 2 μg (selected dose based on a previous study) (22) of various encapsulated or nonencapsulated ODN formulations (except for negative control chickens). The aerosol chamber set up for administering vaccine formulations in this experiment was previously established and tested for delivery of cell-free aerosols of Marek's disease virus in our laboratory (1). The dimensions of the chambers were 40 × 20 × 25 cm and aerosols (1.91 μm in size) were nebulized into the chamber as previously described (1). Sera and lacrimal secretions were collected weekly until day 42 posthatch for evaluation of the dynamics of serum and mucosal immune responses after administration of vaccine formulations using two routes of delivery.
AIV, avian influenza virus; PLGA, poly (
Encapsulation of CpG ODN 2007 and non-CpG ODN 2007 in nanoparticles
The polyethylenimine (PEI)-ODN (CpG 2007 or non-CpG) complex was formed by mixing 434 μg of ODN/125 μL of DNAse/RNAse-free water with a solution of PEI [0.29 mg PEI (low molecular weight, Sigma-Aldrich)] prepared in 150 mM NaCl as described by Boussif et al. (3). This complex was sonicated in 56 mg of PLGA Resomer® RG 503H (acid terminated, 24–38 kD Sigma-Aldrich), then dissolved in 1.25 mL of dichloromethane (Sigma-Aldrich) using a tip sonicator (Ultrasonic processor, 3 mm probe diameter; Fisher scientific) for 1 min (10 s sonication and 5 s pause at 40% amplitude). Subsequently, the resulting emulsion was sonicated with 3.25 mL of 2% polyvinyl alcohol (PVA) and 1% Poloxamer 407 (30–70 kD and 87–90% hydrolyzed PVA and Poloxamer; Sigma-Aldrich) as described above. This emulsion was poured into 50 mL of 2% PVA/1% Poloxamer solution and stirred for evaporation of dichloromethane. The particles were collected by centrifugation at 20,000 g for 30 min at 4°C, washed three times in HyClone, Molecular Biology-Grade water (GE, Healthcare Life Sciences), lyophilized, and stored at 4°C until use. Blank PLGA NPs (mock-NPs) were produced using a similar method.
The loading and encapsulation efficiency of ODNs was determined by dissolving a known quantity of lyophilized NPs (1 mg/mL in Tris-EDTA buffer) in 1 ml dichloromethane (20). The amount of ODNs released into the aqueous solution was measured by the Quant-iT™ OliGreen® ssDNA Reagent and Kit system (Invitrogen) and with a GloMax®-Multi Detection System Fluorometer (Promega). The size of the NPs was determined by the dynamic light scattering method (Zetasizer Nano, Malvern Instruments).
IgM, IgY, and IgA enzyme-linked immunosorbent assays
Serum IgM and IgY antibody titers were determined as reported previously (22). Assay conditions remained unchanged for determining virus-specific IgY antibody titers in lacrimal secretions; however, modifications were made for IgA. Briefly, a whole, inactivated H9N2 virus concentration of 0.5 μg/100 μL of carbonate–bicarbonate buffer (pH 9.6) was used for coating 96-well polystyrene plates (Nunc; MaxiSorp) at 4°C overnight. The plates were washed and blocked as previously described (22) before addition of 100 μL of a 1/10 dilution of lacrimal secretions to duplicate wells, followed by a 1 h incubation period. After washing, 100 μL of mouse anti-chicken IgA (AbD Serotec) was added to each well at a 1/500 dilution. The plates were washed and 100 μL of goat anti-mouse IgG (H/L), HRP (STAR 117P; AbD Serotec), was added at a dilution of 1/10,000. After incubation for an hour, the plates were washed and 100 μL of an HRP substrate solution (ABTS peroxidase substrate system, Kirkegaard and Perry Laboratories) was added and the plates were incubated for 2 h at room temperature in the dark. One hundred microliters of a stop solution (1% sodium dodecyl sulfate) was added before determining the optical density at a wavelength of 405 nm and calculating the sample to positive ratios as described previously (22).
Hemagglutination inhibition
The hemagglutination inhibition (HI) assay was performed as described previously (19) and antibody titers were determined based on the reciprocal of the highest serum dilution resulting in complete HI of red blood cells.
Statistical analyses
General linear models (GLMs) were used in SAS (version 9.2 Cary) to compare immune responses among experimental groups. The interaction of ODN type with encapsulation was examined. HI antibody titers were expressed as the log2 of the reciprocal of the endpoint dilution; S/P ratios were used for analysis of enzyme-linked immunosorbent assays (ELISA) data. Duncan's multiple range test was used to compare multiple groups post-GLM. A p < 0.05 was considered as significant.
Results
Generation of HI antibody responses by different vaccines delivered intramuscularly
To assess antibody responses generated with intramuscular administration of vaccine formulations, HI antibody titers were determined in serum samples. Mean HI antibody titers (Fig. 1) of all vaccine formulations peaked at 1 week postsecondary vaccination, based on the duration of the study. Vaccine formulations containing nonencapsulated CpG 2007 elicited significantly higher HI antibody titers compared to vaccine adjuvanted with PLGA nanoparticles or the inactivated AIV vaccine formulation (without adjuvant) at 1 week postsecondary vaccination. Antibody titers measured by the HI assay were compared between the vaccine adjuvanted with nonencapsulated CpG 2007 and that containing encapsulated CpG 2007. The interaction term “ODN type*encapsulation” was not significant (p > 0.05), indicating that encapsulation of CpG 2007 and encapsulation of non CpG 2007 had similar effects on HI responses. Significantly higher HI antibody titers were observed with vaccines adjuvanted with nonencapsulated CpG 2007 compared to encapsulated CpG 2007 from 1 to 3 weeks postsecondary vaccination.
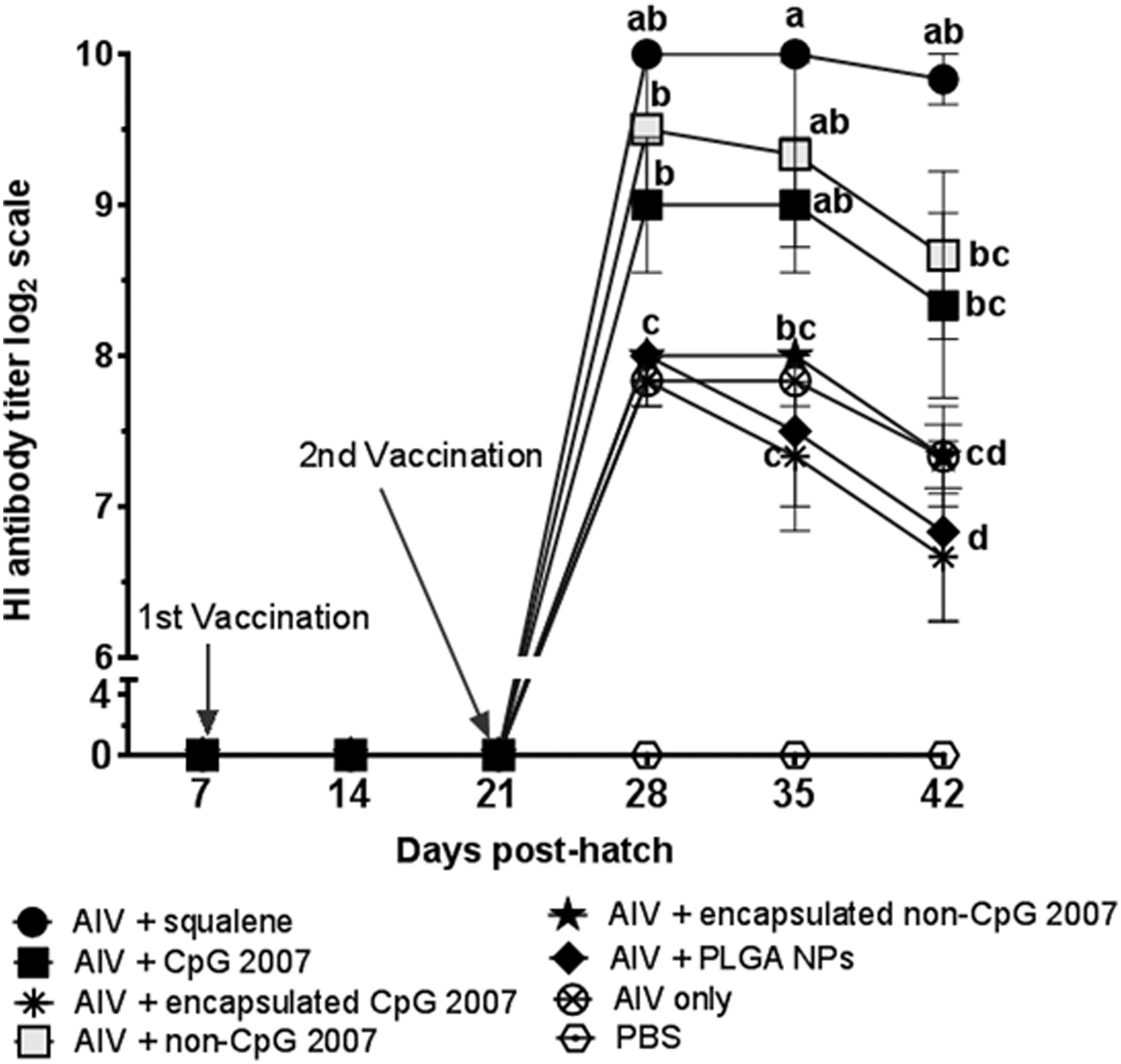
Mean serum hemagglutination inhibition (HI) titers in chickens vaccinated intramuscularly at 7 and 21 days posthatch using encapsulated or nonencapsulated CpG ODN 2007 with formalin-inactivated H9N2 virus. A squalene-containing commercial oil-in-water emulsion (AddaVax™) was used as a positive control adjuvant. Standard errors of the mean are represented by error bars. Within a single time point, means with the same alphabetical letter (a, b, c, or d) do not differ significantly. A PROC general linear model with Duncan's Multiple Range test was used for data analysis with Duncan's Multiple Range test, ANOVA (p < 0.05).
Virus-specific IgY and IgM antibody S/P ratios in serum generated by intramuscular administration
Virus-specific IgY and IgM antibody responses in serum generated by intramuscular administration of vaccine formulations were determined. At 3 weeks postsecondary vaccination, no statistically significant differences were observed in mean serum IgY antibody S/P ratios between vaccine formulations containing nonencapsulated CpG 2007 (group 2) and nonencapsulated non-CpG 2007 [group 4 (Fig. 2A)]. However, mean serum IgY S/P ratios induced by the vaccine adjuvanted with nonencapsulated non-CpG 2007 were significantly higher than those for vaccines containing squalene (group 1), encapsulated CpG 2007 (group 3), or inactivated AIV only (group 7). No significant differences were observed in mean serum IgY S/P ratios between vaccines containing nonencapsulated or encapsulated CpG 2007 at 3 weeks postsecondary vaccination. At 1 week postprimary vaccination (day 14), mean IgM S/P ratios in chickens induced by the vaccine containing squalene were significantly higher than those elicited by the vaccine adjuvanted with nonencapsulated CpG 2007 (Fig. 2B). A decrease in mean IgM S/P ratios was observed in chickens receiving the vaccine containing non-CpG 2007 at 2 weeks postprimary vaccination (day 21), followed by an increase until 3 weeks postsecondary vaccination (day 42). At 1 week postsecondary vaccination (day 28), mean IgM S/P ratios in chickens receiving the vaccine adjuvanted with nonencapsulated CpG 2007 were statistically similar to those attained in chickens treated with vaccines adjuvanted with squalene or encapsulated CpG 2007.
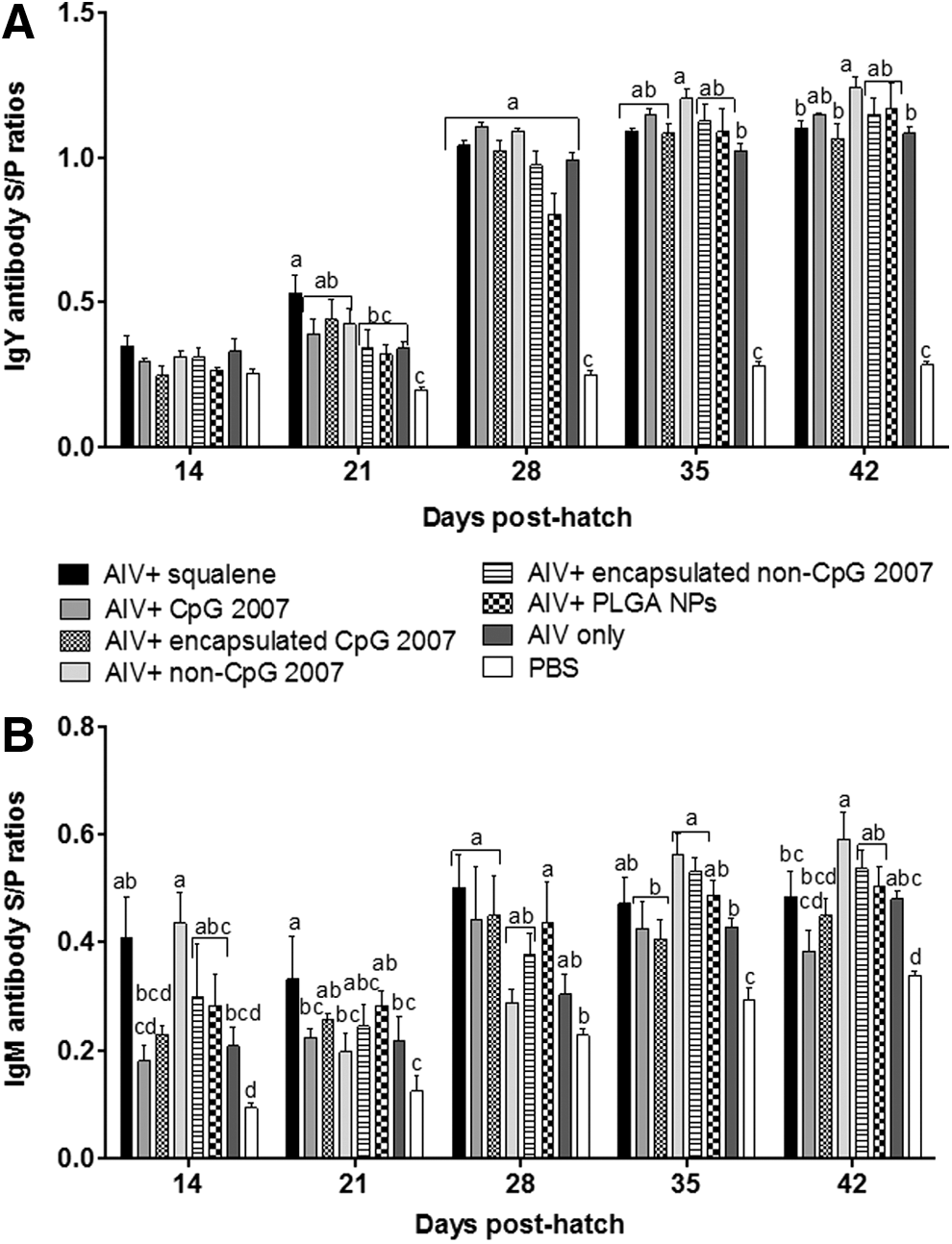
Serum IgY
Virus-specific IgY antibody S/P ratios in lacrimal secretions generated by intramuscular administration
At 2 weeks postsecondary vaccination (day 35), mean IgY S/P ratios in lacrimal secretions (Fig. 3) induced by the vaccine containing nonencapsulated CpG 2007 were significantly higher than the ratios generated by the vaccine adjuvanted with encapsulated CpG 2007. No significant differences were observed between the vaccines adjuvanted with squalene or nonencapsulated CpG 2007 and the AIV only formulation at 2 weeks postsecondary vaccination. At 3 weeks postsecondary vaccination (day 42), mean IgY S/P ratios in lacrimal secretions elicited by the vaccine containing nonencapsulated CpG 2007 were higher than ratios generated by the vaccine adjuvanted with encapsulated CpG 2007; however, this difference was not significant. Mean IgY S/P ratios generated by the vaccine adjuvanted with squalene were not significantly higher than those of AIV only; however, these IgY responses were significantly higher than those induced by the vaccine containing encapsulated CpG 2007.
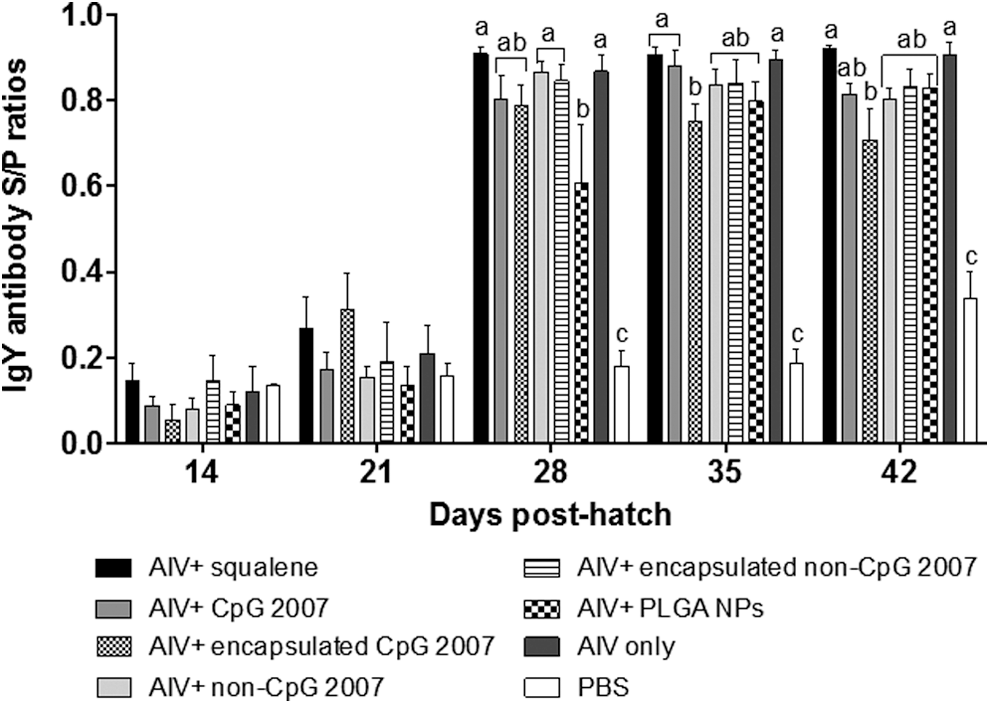
IgY S/P ratios in lacrimal secretions for chickens vaccinated intramuscularly with encapsulated or nonencapsulated CpG ODN 2007 with formalin-inactivated H9N2 virus. Within a single time point, means with the same alphabetical letter (a, b, or c) do not differ significantly (p < 0.05).
Serum HI and IgY antibody responses generated by vaccines administered through the aerosol route
To determine if the administration route had an effect on immunogenicity of the encapsulated formulation versus the nonencapsulated vaccine formulation, vaccines were delivered by the aerosol route. Significantly higher HI antibody titers (Fig. 4) were generated at 1 week post-tertiary vaccination (day 42) in chickens receiving the vaccine adjuvanted with encapsulated CpG 2007 compared to the formulation with nonencapsulated CpG 2007. No significant differences were observed between inactivated AIV vaccines adjuvanted with nonencapsulated CpG 2007 and nonencapsulated non-CpG 2007 at 1 week post-tertiary vaccination. Although the encapsulated CpG 2007 vaccine formulation elicited higher mean serum IgY S/P ratios (Fig. 5A) compared to the nonencapsulated CpG 2007 formulation at 1 week post-tertiary vaccination, results were not significant. Mean IgY S/P ratios induced by the vaccine containing encapsulated non-CpG 2007 were significantly higher compared to the nonencapsulated formulation at 1 week post-tertiary vaccination.
Mean serum HI titers in chickens vaccinated at 7, 21, and 35 days posthatch using the aerosol route with formalin-inactivated H9N2 with encapsulated or nonencapsulated CpG ODN 2007. Within a single time point, means with the same alphabetical letter (a, b, or c) do not differ significantly (p < 0.05).
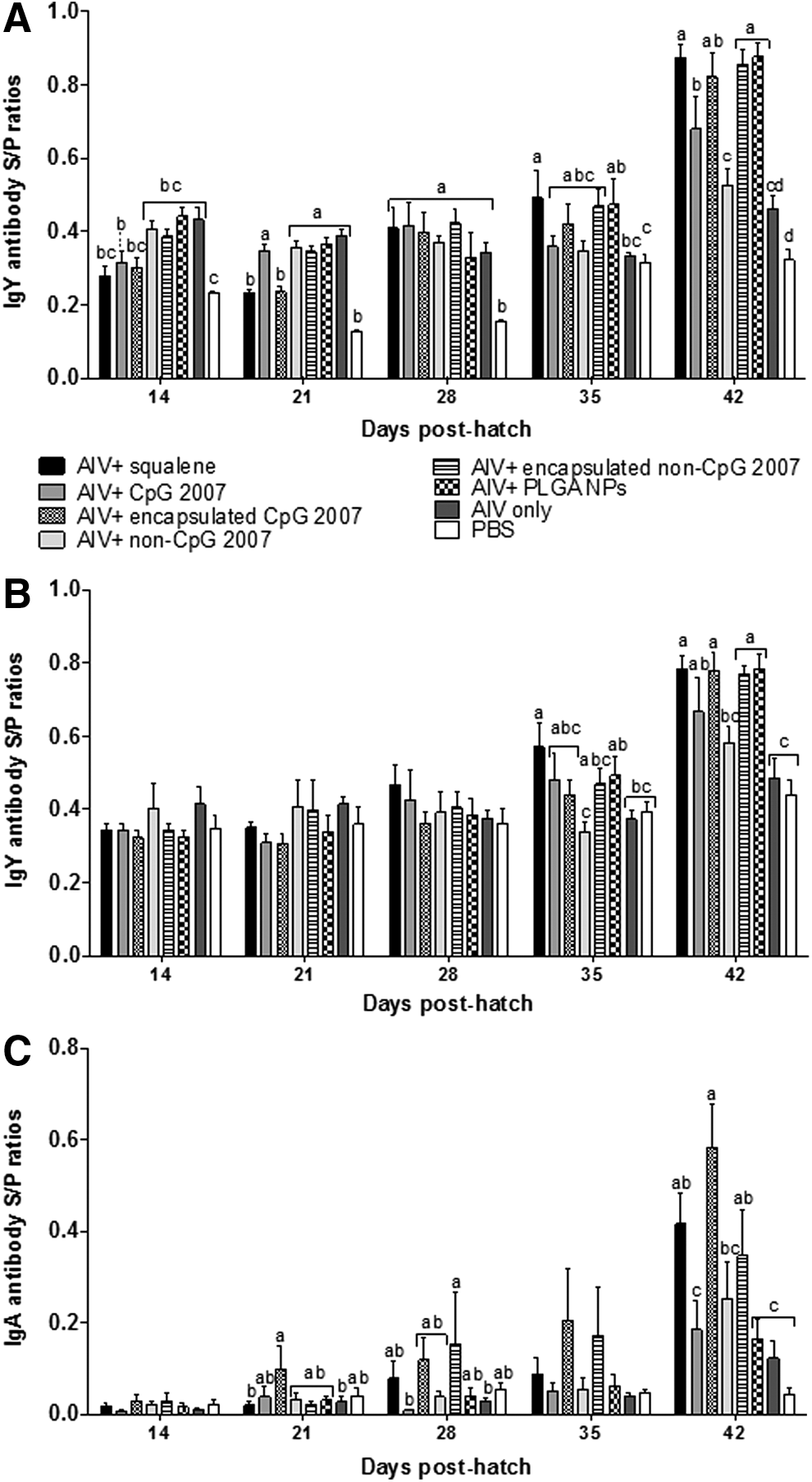
Serum IgY S/P ratios
Virus-specific IgY and IgA S/P ratios in lacrimal secretions generated by aerosol administration
The vaccine formulation adjuvanted with encapsulated CpG 2007 generated higher IgY S/P ratios in lacrimal secretions compared to the formulation with nonencapsulated CpG 2007 at 1 week post-tertiary vaccination (day 42) although these results were not statistically significant (Fig. 5B); however, when compared to the inactivated AIV only formulation, significantly higher IgY S/P ratios were observed. The correlation between serum IgY antibody levels (Fig. 5A) and IgY antibodies in lacrimal secretions (Fig. 5B) was evaluated using Pearson's correlation at 1 week post-tertiary vaccination (day 42) for nonencapsulated and encapsulated vaccine formulations delivered by the aerosol route [R2 = 0.447 (p < 0.02)]. Mean IgA S/P ratios (Fig. 5C) were significantly higher in lacrimal secretions of chickens receiving the AIV vaccine adjuvanted with encapsulated CpG 2007 compared to the vaccine formulation containing nonencapsulated CpG 2007 at 1 week post-tertiary vaccination (day 42).
Discussion
We previously reported the vaccine efficacy of nonencapsulated CpG 2007 at a dose of 2 μg, when administered intramuscularly with 15 μg of formalin-inactivated H9N2 virus (22). In this study, we examined the effect of PLGA encapsulation of the CpG ODN adjuvant component on the kinetics of the immune response to formalin-inactivated AIV vaccine formulations using intramuscular and aerosol routes of administration.
The encapsulated CpG 2007 vaccine formulation, when administered by aerosol route, generated significantly higher antibody responses based on HI titers compared to the formulation adjuvanted with nonencapsulated CpG 2007 at 1 week post-tertiary vaccination (day 42 posthatch). Furthermore, the encapsulated CpG 2007 vaccine formulation induced significantly higher IgA S/P ratios in lacrimal secretions at 1 week post-tertiary vaccination compared to the nonencapsulated CpG 2007 formulation. Although mean serum IgY S/P ratios generated by the encapsulated CpG 2007 vaccine formulation were higher than IgY S/P ratios generated with the nonencapsulated CpG 2007 formulation, these findings were not significant. Serum IgY S/P ratios were moderately correlated (R2 = 0.447) with IgY S/P ratios in lacrimal secretions for vaccines delivered by the aerosol route at 1 week post-tertiary vaccination. Therefore, the majority of IgY antibody levels in lacrimal secretions was likely due to local production as opposed to serum transudation. A previous report by de Geus et al. (9) suggests that the lack of detectable antibodies against inactivated influenza virus in chickens when administered through aerosol may be explained by insufficient virus concentrations or a requirement for booster vaccination. Our findings suggest that PLGA encapsulated, adjuvanted, inactivated AIV vaccine formulations were more efficacious than nonencapsulated, adjuvanted formulations when administered by the aerosol route. Our data also suggest that at least two boosters are required for the induction of an immune response with inactivated AIV vaccine formulations when given by aerosol administration. It is also noteworthy that, in mice, particulate vaccine adjuvants have been shown to promote NALP3 inflammasome activation and the production of proinflammatory cytokines (21). Although, the existence of the NALP3 inflammasome in chickens remains undetermined, we must consider the potential implication of the combination of encapsulated CpG 2007 and PLGA nanoparticles in terms of overall enhancement of antibody-mediated immune responses. The size of the encapsulated ODNs used in these experiments was estimated to be 674.5 nm based on dynamic light scattering. We speculate that enhanced vaccine efficacy observed with PLGA-encapsulated formulations delivered by the aerosol route may be attributed to increased residential time at respiratory mucosal surfaces providing the opportunity for maximum interaction with APCs. Considering the size of the nanoparticles, it is likely that deposition occurred at the level of the lower respiratory tract, allowing access to APCs located in the subepithelium, connective tissue, and parabronchial linings (7,8).
In contrast to aerosol administration, inactivated vaccines containing nonencapsulated CpG 2007 induced significantly higher immune responses compared to encapsulated formulations based on serum HI titers from 1 to 3 weeks postsecondary vaccination, and at 2 weeks postsecondary vaccination for lacrimal IgY S/P ratios when delivered by the intramuscular route. Local environmental conditions at both sites of administration such as pH, presence of enzymes, and temperature (4) might account for some observed differences. Furthermore, although in vivo nanoparticle decomposition is required for release of encapsulated contents, predicting in vivo behavioral characteristics of nanoparticles is still in its infancy. Additionally, it must be noted that administration of CpG ODN through different routes (systemic versus local) has been demonstrated to result in immunosuppressive or immunostimulatory effects at some dosage levels in other species (31).
Unexpected findings included vaccine immunogenicity in the context of encapsulated non-CpG 2007 and nonencapsulated non-CpG 2007 for both routes of administration. With respect to the aerosol route of delivery, the encapsulated non-CpG 2007 vaccine formulation generated significantly higher mean IgY S/P ratios in both serum and lacrimal secretions compared to the respective nonencapsulated adjuvanted formulation at 1 week post-tertiary vaccination. Furthermore, although the vaccine containing nonencapsulated non-CpG 2007 elicited an early IgM response compared to nonencapsulated CpG 2007 during intramuscular administration, the exact mechanism of action of non-CpG ODNs remains to be investigated in chickens. Interestingly, St. Paul et al. have observed that in vivo administration of non-CpG 2007 at a dose of 50 μg was able to induce similar levels of cytokine gene expression in chicken spleens to CpG 2007. These cytokine transcripts were IFN-α, IFN-γ, IL-1β, IL-8, MyD88, and major histocompatibility complex II (26). Recent studies have demonstrated the ability of non-CpG ODN to influence early T cell and late B cell activation in mice (13). Moreover, non-CpG ODNs have also been shown to costimulate mouse and human CD4+T cells through a MyD88-independent mechanism (17). Finally, it has been reported that non-CpG ODNs can act synergistically when combined with specific antigen resulting in the activation of the NF-κB pathway and B cell proliferation in mice (30).
In this study, PLGA nanoparticles were used to encapsulate CpG 2007 by a water-in-oil-in-water (double emulsion) technique for aerosol and intramuscular delivery with inactivated H9N2 AIV. We showed that nonencapsulated CpG 2007 in inactivated AIV vaccines induced higher immune responses when delivered intramuscularly in chickens. Furthermore, we showed that encapsulated CpG 2007 in inactivated AIV vaccines generated higher responses than nonencapsulated AIV vaccines with aerosol administration. An implication of these results is that encapsulated vaccine formulations may be more applicable to mucosal administration than to systemic administration in chickens. Future studies may be aimed at investigating protective effects of these vaccine formulations against homologous and heterologous virus challenge.
Footnotes
Acknowledgments
This research was funded by the Canadian Poultry Research Council, Agriculture and Agri-Food Canada Poultry Cluster, and the Ontario Ministry of Agriculture, Food, and Rural Affairs (OMAFRA). The authors wish to thank the staff members at the Ontario Veterinary College Isolation Facility for assistance with the housing and daily care of the chickens.
Author Disclosure Statement
No competing financial interests exist.