
Abstract
Purinergic P2X receptors are plasma membrane bound, ATP-gated ion channels that are expressed on wide range of cells and respond to varying ATP concentrations in extracellular environment. Upon activation they increase membrane permeability for Ca2+ ions and trigger a cascade of signaling complexes. During the course of hepatitis C virus (HCV) infection, ATP is released from the infected hepatocyte, which binds with Purinergic receptors (P2X) on peripheral blood mononuclear cells (PBMCs) and initiate downstream signaling pathways by disturbing the ionic balance of the cell. The present study investigates quantitative expression of P2X7 and P2X4 along with selected host genes PEPCK, transforming growth factor β (TGF-β), MAPK, Rho, and Akt in PBMCs of chronic HCV infection patients. PBMCs were isolated from collected blood samples of study subjects. Transcript analysis of P2X7, P2X4, and targeted downstream genes was done using quantitative real-time polymerase chain reaction. Relative expression analysis was performed by unpaired Student's t test on GraphPad Prism version 5. We found a notable increase of threefolds and 1.8-folds in the expression of P2X7 and P2X4 receptors in treatment naïve category while the expression of PEPCK, TGF-β, MAPK, AKT, and Rho A increased by 2.8, 1.9, 2.2, 2.2, and 1.8-folds, respectively. In sustained virological response patients, P2X7 significantly increased up to 3.5-folds while the expression of P2X4 receptor was increased up to twofold. In third category, treatment nonresponder, the expression of P2X7, P2X4 receptors, and targeted markers remained un-altered. This study deals with two major aspects of P2X4 and P2X7 receptors in PBMCs of chronic HCV individuals. One is their role in providing antiviral immunity to host against HCV; second aspect is the role of P2X receptors in inducing HCV pathogenesis via AKT, TGF-β, Rho A, PEPCK, and MAPK expression.
Introduction
H
HCV is a positive sense single-stranded virus that belongs to family Flaviviridae (30). To date, 6 major genotypes (GTs) and more than 100 subtypes of HCV have been identified. Among these GT-3a is the most commonly occurring in Pakistan (19). Patients infected HCV GT-2 or -3, undergo standard treatment of pegylated interferon plus ribavirin (PEG-IFN/RBV) and patients infected with HCV GT-1 include one of the two recently approved first generation protease inhibitors, telaprevir and boceprevir, to PEG-IFN/RBV for 48 weeks (15). Sofosbuvir, NS5B polymerase inhibitor is a once-daily HCV nucleotide-analog approved for the treatment of chronic infections with HCV GTs 1–4. In a phase 3 trial in patients infected with HCV GT-1, treatment-naive patients, including a subgroup with compensated cirrhosis, 89% achieved sustained virological response (SVR) after 12 weeks of treatment with sofosbuvir and peginterferon or ribavirin 24 weeks (26).
HCV mainly attacks the hepatocytes but some studies have reported its invasion in cells other than hepatocytes. These include peripheral blood mononuclear cells (PBMCs), bone marrow, and central nervous system. Viral proteins interact with numerous cellular mechanisms in a complex fashion that leads to a variety of pathological events (1).
Purinergic receptors comprise of two families, P2X and P2Y. P2X family includes seven receptors, P2X1, P2X2, P2X3, P2X4, P2X5, P2X6, and P2X7. All of these have different affinity for ATP (5). P2X receptors are ATP-gated ion channels that undergoes conformational changes upon binding of ATP to their extracellular portion resulting in efflux of potassium ions (K+) and influx of calcium (Ca2+) and sodium Na+ ions. P2X4 and P2X7 are capable of undergoing further conformational changes that greatly enhances permeability of membrane (31). Membrane depolarization consequently activates voltage-gated calcium channels and thus accumulating calcium ions (Ca2+) in the cytoplasm. P2X receptors activate paracrine and autocrine signaling and majority of them are expressed on liver cells and PBMCs (18). P2X have been characterized in various immune cells including lymphocytes, platelets, and macrophages, where they have been associated with apoptosis, migration, platelet aggregation, and cytokine release (4,43).
Excess of the extracellular ATP released from the stressed cells trigger immune cells with the help of Purinergic signaling (3). Purinergic receptors are not only involved in chemotaxis but also in the modulation of immune responses (34). In addition, continuous elevated levels of ATP cause desensitization of the receptor that initiates perturbed downstream signaling and ultimately cause heightened inflammation, insulin resistance, vascular injury, and abnormal liver cell regeneration (7). Similarly, several stages of the life of a T lymphocyte may be affected by purinergic signaling, including differentiation, activation, and proliferation. Thymocytes are susceptible to apoptosis in the presence of extracellular ATP via the activation of purinergic receptors in vitro and in vivo (31). P2X7 receptors have a known role in abrupt activation of caspase-1 and the release of inflammatory cytokine interleukin 1β (IL-1β) from activated macrophage and microglia (13). P2X7 receptor also impairs the suppressive activity of regulatory T cells (Treg) by converting them into Th17 cells that secrete IL-17 (33). P2X7 receptor shows a marked homology (40%) with P2X4 receptor at amino acid level (35). Previously it was thought that P2X4 is only proviral having a major role in HCV induced pathogenesis (27) but several studies have identified its synergistic role with P2X7 along with pannexin-1 in facilitating inflammasome formation, programmed cell death, and generation of reactive oxygen species (ROS) (17,22,35). Combined expression of P2X4 and P2X7 was observed in immune cells where they contribute to inflammation (3,45). In a study it was demonstrated that P2X4 receptor also cooperates with P2X7 receptors to enhance inflammation in dendritic cells derived from mouse bone marrow (35).
In our previous studies we have demonstrated the pro-antiviral responses of purinergic receptors (1,27). And it is well established that P2X receptors have a role in ATP-induced apoptosis, fibrosis, cell volume regulation, and hepatocellular carcinoma (HCC). This can further be correlated with activation of P2X receptors and downstream activation of genes involved in HCV-induced pathogenesis.
Our previous studies have characterized the role of P2X receptors in HCV pathogenesis and in inducing antiviral immune response (1,27); however, their downstream signaling events were not focused. Hence, this study was designed to analyze probable association of liver pathophysiology and P2X receptors signaling in HCV infection. This was observed while taking the transcript expression of P2X receptors and targeted host genes reportedly involved in pathophysiology of liver. Targeted host genes include AKT, transforming growth factor β (TGF-β), Rho A, PEPCK, and MAPK that may be triggered by P2X receptors in PBMCs of HCV-infected individuals. Three groups were formulated for the experiment. These were (i) treatment responders, (ii) treatment nonresponders, (iii) treatment naïve category patients. The study may help to comprehend relation of P2X receptors with immune system and disease pathogenesis in a much better way and will provide possible solutions for combating HCV-induced complications.
Materials and Methods
Study design
A total of 30 HCV-infected patients were recruited. These patients were grouped into treatment naïve, nonresponders, and responder's category with SVR on the basis of their response to standard HCV therapy, that is, PEGylated interferon and Ribavirin. Same numbers of healthy patients were enrolled for control samples. Written and informed consent was taken from all the HCV patients and healthy individuals. The study was approved by the Ethical Committees of Atta-Ur-Rahman School of Applied Biosciences (ASAB), NUST Pakistan and Armed Forces Institute of Pathology (AFIP), Rawalpindi (Pakistan).
Collection of blood samples
Peripheral blood samples of chronic HCV subjects were collected from AFIP Rawalpindi (Pakistan) for the duration January 2015 to June 2015. Samples of healthy individuals were provided by diagnostic laboratories of Atta-Ur-Rahman School of Applied Biological Sciences (H-12 Campus, Islamabad, Pakistan). All HCV patients were positive for GT-3a, which is most prevalent in Pakistan. The age range of these patients was between 25 and 55 years. Moreover, the samples were inspected for co infection with other viruses like hepatitis B virus (HBV) and HIV. Such patients who were infected with other viruses or had any comorbidities were excluded from the study. Samples were confirmed for the presence of HCV RNA using Applied Biosystems® 7300 quantitative real-time polymerase chain reaction (PCR).
PBMCs isolation
PBMCs were isolated from peripheral blood of diseased patients and healthy controls. For this purpose Histopaque®-1077 (Catalogue No. 10771; Sigma-Aldrich) was used, which acts as density gradient media and helps in separating various blood cells. The opaque layer of peripheral mononuclear cells at the interphase was collected and washed twice with phosphate buffer saline before further utilization in experiment.
Extraction of RNA from PBMCs and complementary DNA synthesis
Cellular RNA was isolated using TRIzol reagent (Catalog No.15596-026; Invitrogen). Homogenizing of sample, phase separation, RNA precipitation, washing and suspension constitute major steps of RNA extraction. Extracted RNA was treated with DNase I (Catalog No. EN0521; Fermentas) to prevent DNA contamination. The RNA pellet was resuspended in 20 μL of DEPC-treated water. The quantity and quality of extracted RNA was determined using Eppendrof's Biophotometer Plus. Furthermore, RNA was qualitatively analyzed on 1% agarose gel stained with ethidium bromide. complementary DNA (cDNA) of 1 μg was synthesized from previously extracted RNA using oligo (dT) (Fermentas) and RevertAid H minus reverse transcriptase (Fermentas) along with Ribolock RNase Inhibitor (Fermentas).
Qualitative and quantitative expression of targeted genes
Gene-specific primers were designed using Primer 3 software (Table 1). Reaction profile was optimized on conventional PCR (Model No. 2720 Applied Biosystems). Optimization of the primers was done using cDNA of concentration ranging from 1 μg to 5 μg as a template from healthy individuals. GAPDH was used as a positive control. PCR amplicons were observed by gel electrophoresis using 2% agarose gel. Targeted genes and receptors were quantified on Applied Biosystems 7300 Real Time PCR system using SYBER Green qPCR Master Mix (2 × ). Passive dye ROX was used during the PCR run.
F, forward; R, reverse; TGF-β, transforming growth factor β.
The expression of targeted host genes TGF-β, MAPK, Rho, Akt, PEPCK, P2X4, P2X7 and GAPDH was compared in healthy controls and different categories of HCV patients. Results obtained were interpreted by calculation of cycle threshold (Ct) value and analysis of amplification plots. Primer dimers and secondary amplification products were ruled out by analysis of dissociation curve. All reactions were performed in triplicate and GAPDH housekeeping gene was used for normalization to calculate gene/GAPDH fold change.
Statistical analysis
All statistical tests were applied using Graph pad prism software version 5.0. The unpaired student's t test was performed. *p < 0.05, **p < 0.005, ***p < 0.001 was considered significant.
Results
Identification of the targeted genes and P2X receptor expression
Gene-specific primers designed using Primer 3 software were used to determine expression of targeted genes and P2X receptor on PBMCs. High quality cDNA was used for the optimization of selected genes on conventional and real-time PCR. Amplification results of the PCR detected clear expression of P2X4, P2X7, TGF-β, AKT, MAPK, PEPCK, and GAPDH genes on PBMCs when observed using gel electrophoresis.
Transcript analysis of selected markers in treatment naïve patients
Quantitative expression analysis of selected markers in treatment naïve category and healthy individuals showed elevated expression of P2X7, P2X4, PEPCK and TGF-β. P2X7 and P2X4 receptor expressions were raised by threefolds and 1.8-folds, respectively. Expression of PEPCK was increased by 2.8-folds and Rho A was increased by 1.8-folds. Least statistically significant results were observed in case of TGF-β, MAPK, and Akt (Fig. 1).
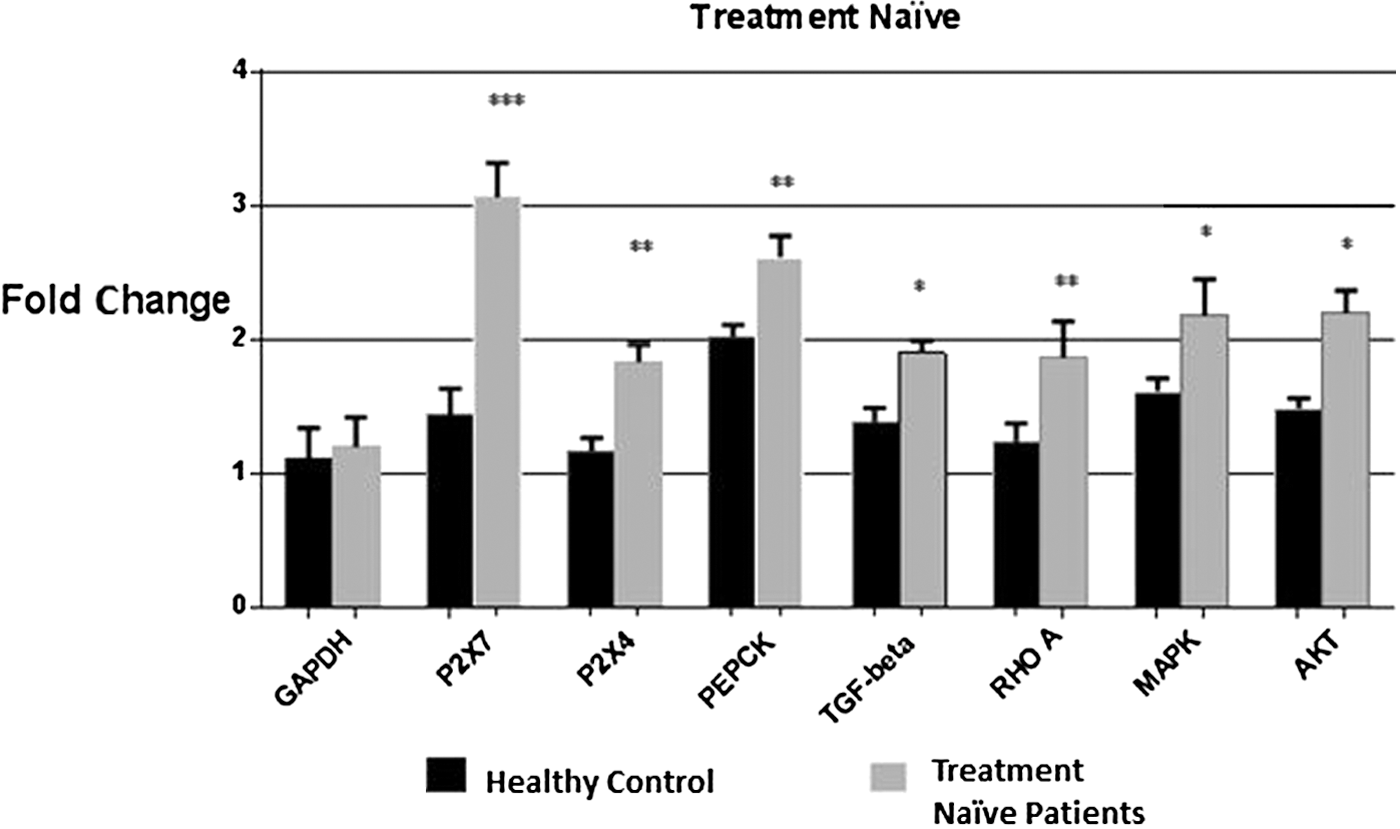
Comparative expression analysis of the targeted genes and P2X receptors in treatment naïve category and healthy controls. Graph showing significant increase in mRNA levels of all targeted genes with maximum expression observed in case of P2X7 receptors (threefolds). Student's t-test was used and all the values were expressed as ±SEM. P-values <0.05 are considered significant and *p < 0.05, **p < 0.01, ***p < 0.001 indicate low significance, medium significance, and high significance of data, respectively. mRNA, messenger RNA; SEM, standard error of the mean.
Transcript analysis of selected markers in treatment nonresponders
Patients included in this group received standard therapy for 24 weeks against HCV, but they failed to show any response to this treatment. Expression analysis of selected genes in the immune cells was done to find out the causes of their negative response to antiviral treatments. Transcript expression of selected genes in this category did not show any significant difference in expression as compared to healthy controls (Fig. 2).
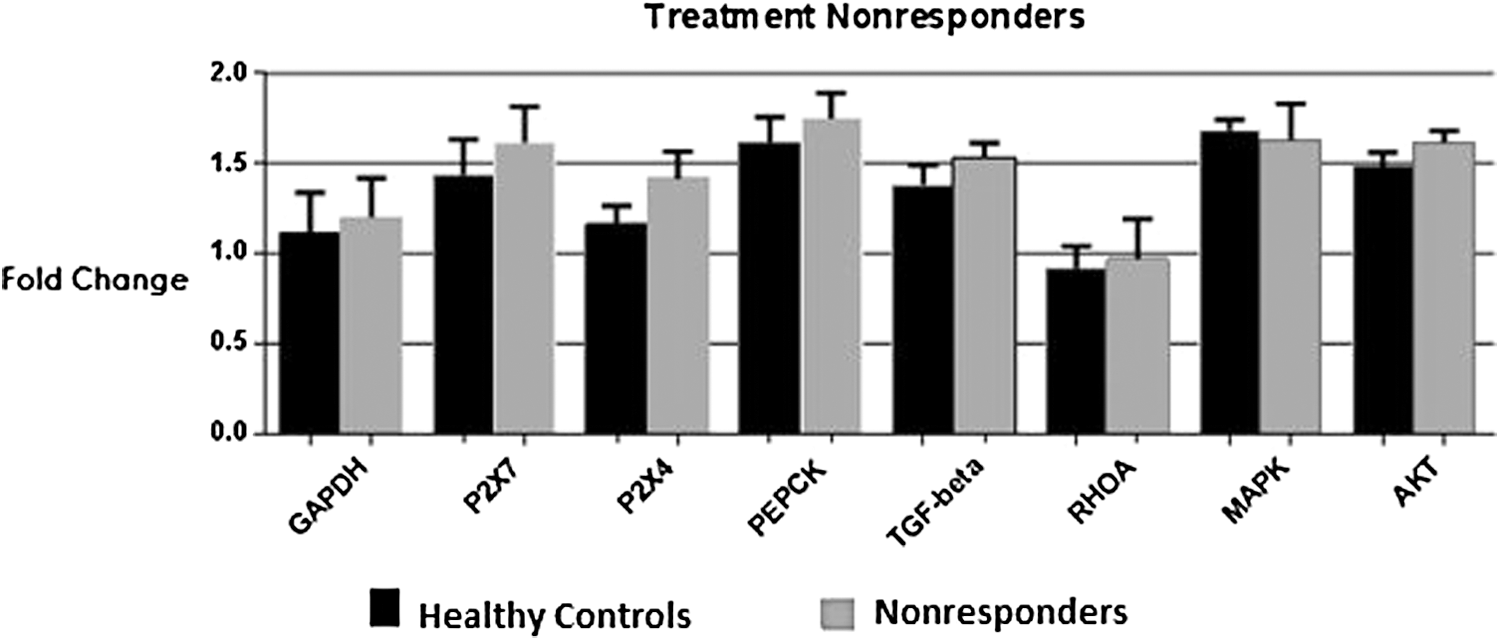
Targeted genes and P2X4, P2X7 receptor expression in PBMCs of treatment nonresponders in comparison with healthy controls. The expression levels of purinergic receptors P2X4 and P2X7 along with host genes PEPCK, TGF-β, RhoA, MAPK, and Akt remained unaltered. Student's t-test was applied and all the values were expressed as ±SEM, p < 0.05. Graph shows no significant difference in expression. HCV, hepatitis C virus; PBMCs, peripheral blood mononuclear cells; TGF-β, transforming growth factor β.
Transcript analysis of selected markers in patients with SVR
Patients included in this category are those who received antiviral therapy against HCV and showed complete viral clearance 24 weeks after completion of therapy. The expression analysis of the targeted genes showed that P2X7 was significantly increased by 3.5-folds; however, expression of other genes was altered insignificantly (Fig. 3).
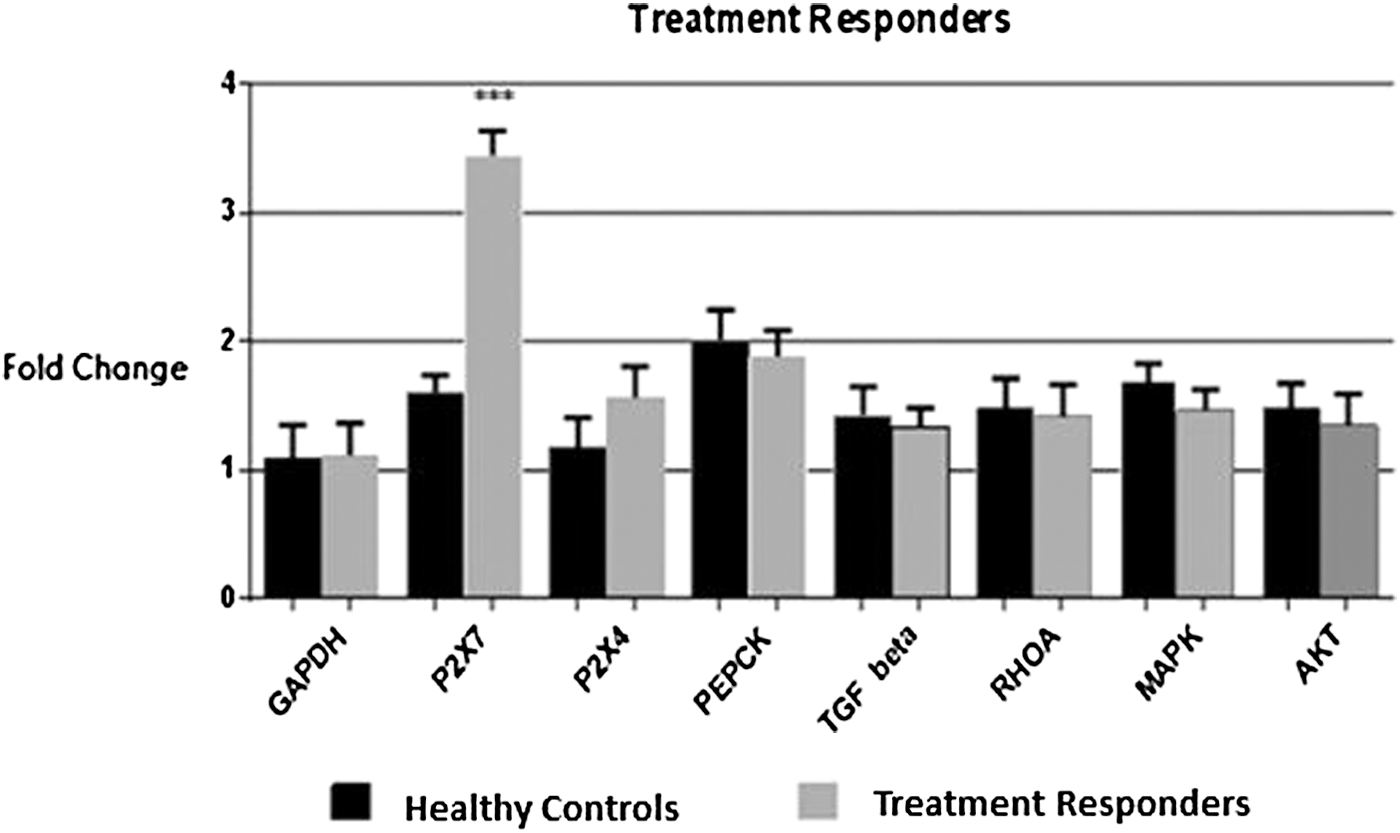
Expression analysis of the targeted genes and P2X4, P2X7 receptors in treatment responders HCV patients in comparison with healthy controls. Statistically significant expression was only detected in case of P2X7 receptors with 3.5-fold increase in expression. Student's t-test was used and all the values were expressed as ±SEM, p < 0.05. ***p < 0.001 indicates highly significant data.
Expression analysis of P2X4 and P2X7 in different categories of HCV patients
P2X7 receptor expression was found to be elevated in treatment naïve patients as well as in SVR (Fig. 4). While the expression of P2X4 was found to be significantly increased in only treatment naïve category of chronic HCV patients (Fig. 5).
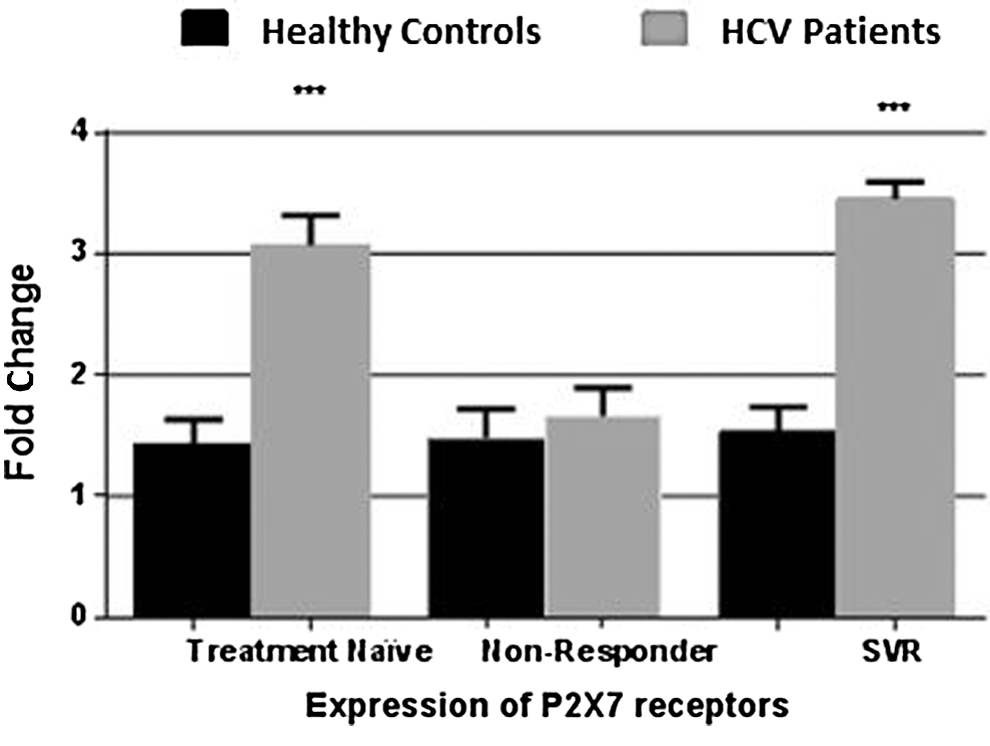
Graph depicts elevated expression of P2X7 receptor in PBMCs of treatment naïve patients and HCV patients with sustained virological response. No significant difference was observed in treatment nonresponders category. All values are expressed as ±SEM with p values <0.05 considered as significant. ***p < 0.001 shows high level of significance among the two categories.
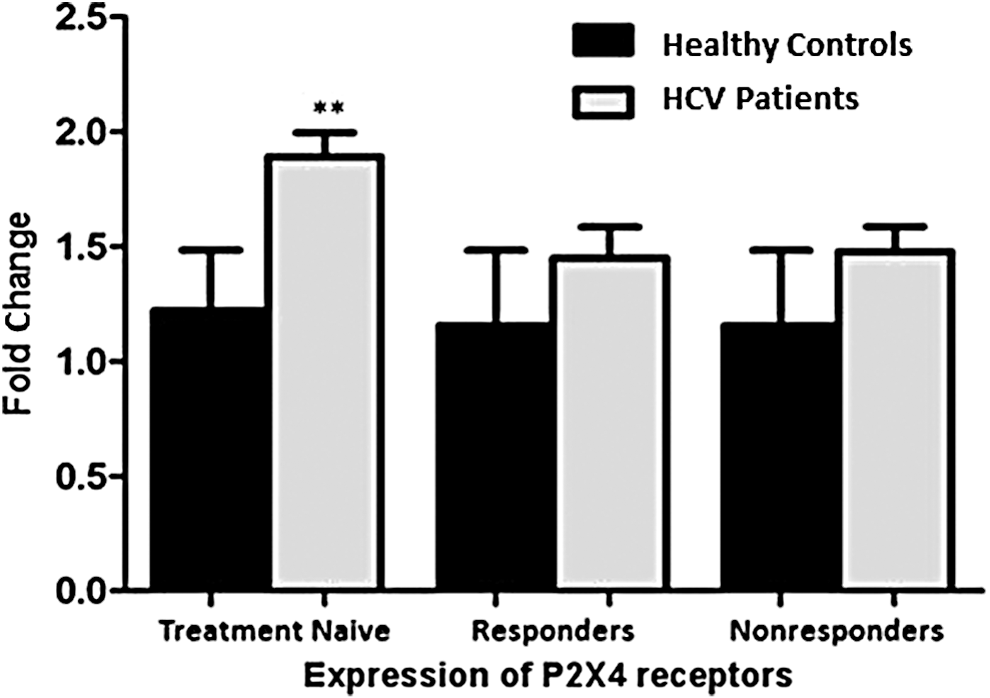
Graph depicts significant increase in expression of P2X4 receptor in PBMCs of treatment naïve category only. All values are expressed as ±SEM with p values <0.05 considered as significant. **p < 0.01.
Discussion
Purinergic signaling constitutes a series of events from the release of extracellular nucleotides to activation of P2X and P2Y receptors followed by regulation of various enzymes and signaling pathways (2). Over the past many scientists have contributed to comprehending possible role of purinergic signaling in disease conditions (6,33), however, the downstream signaling events of P2X receptors in HCV pathogenesis and immunity has been focused for the first time in our study.
During the course of viral infection, ATP is excessively released into the extracellular environment (10). Upon binding of ATP to P2X7R inflammasome gets activated with the release of plenty of cytokines IL-1β, IL-8, IL-6, IL-18, and tumor necrosis factor α (TNF-α) (29). Besides stimulation of P2X7R, excessive exposure to ATP may result in activation of pannexin-1, membrane permeablization, and eventually death of encountered cells, which then release more ATP and further activate macrophages. Both P2X and P2X4 play key role in inflammatory responses by activating the immune cells (18).
In our previous studies, we have revealed the involvement of P2X receptors in pathophysiology of HCV (1,27). This study has investigated the role of P2X7 and P2X4 receptors in downstream signaling during chronic HCV infection. The expression of P2X7, P2X4, PEPCK, TGF-β, AKT, Rho A, and MAPK on PBMCs was determined at transcriptional level in treatment responders, nonresponders, and treatment naïve category patients chronically infected with HCV.
Our findings report interesting results in all categories of chronic HCV patients subjected to study. In treatment naïve patients, there was a significant increase in the expression of P2X7, P2X4, and all targeted markers. P2X7 receptor showed threefolds increase while the expression of P2X4, PEPCK, TGF-β, MAPK, Akt, and RhoA increased up to 1.8, 2.8, 1.9, 2.2, 2.2, and 1.8-folds, respectively. Co-expression of P2X4 and P2X7 was found in many immune and nonimmune cells (9). P2X4 receptor is proviral, it enhances the copy number of virus, and therefore in this category its expression is also high (27). Our previous study regarding overexpression of HCV structural proteins (E1/E2) in huh-7 cell line demonstrated enhanced expression level of P2X4 receptors (27). The enhanced expression of P2X4 initiates all downstream signaling and further activates selected genes in treatment naïve patients. The factors that regulate P2X7 expression have not been explained fully but it has been claimed that different cytokines, including IL-2, IL-6, and TNF-α, and other proinflammatory stimuli upregulate its synthesis (14). In accordance to our study, Epstein–Barr virus (EBV) induced transformation of B cells resulted in elevated expression of P2X7 receptors (25). Another study confirmed that P2X7 and P2X4 are required for fusion of HIV envelope with CD4-expressing cells (33). Moreover, P2X7 are activated via release of high concentration of ATP by Panx-1 hemmichannels and support viral replication in case of HIV (34). Séror et al. reported that HIV-infected cells release ATP through pannexin-1 hemichannels, which is required for initial infection (37). Similarly inhibition of pannexin-1 channels by small interfering RNA (siRNA) suppressed HIV-mediated infection and cell death (40).
The messenger RNA (mRNA) levels of PEPCK enzyme in treatment naïve category was found to be significantly increased. Viral protein NS5A causes mitochondrial damage, which results in the production of ROS. HCV-induced ROS production causes the JNK activation. Its activation results in decrease of phosphorylation and accumulation of FoxO1 in the nucleus. FoxO1 enhances the expression of PEPCK and G6Pase, which eventually leads to increased glucose production by gluconeogenesis. High levels of glucose in the PBMCs then may lead to the enhanced HCV replication (8). In concordance with our study it has been reported earlier that P2X4 expression is increased in hepatocytes that are rich in glycogen suggesting its involvement in glucose metabolism (12).
Purinergic signaling also confers to antigen recognition by T cells after formation of immunological synapse (33). Recently, it was revealed that pannexin-1 and P2X4 receptors translocate to immune synapse and contribute to Ca2+ transport and T-cell activation (44). Upon T-cell stimulation CD4 T cells release ATP, which stimulates MAPK and secretion of IL-2 (36,46). P2X7 receptor also mediates MAPK pathway that leads to proliferation of cells (20). Elevated expression of MAPK in treatment naïve patients may depict stimulation and active proliferation of T cells. Consistent to our findings it was reported earlier that in astrocytes, activation of the P2X7 receptor triggers MAPKs ERK1, ERK2, and p38, resulting in increased level of monocyte chemoattractant protein-1 expression (32). This shows its involvement in inflammation and immunity.
Furthermore, the expression of TGF-β was also increased up to twofolds in PBMCs of treatment naïve HCV patients. TGF-β phosphorylates and activates its receptors ALK-1 and ALK-5. ALK-1 releases various cytokines that activate c-Abl, which further causes the phosphorylation of Smad1 and Smad4 (15). Kountouras et al. also reported the induction of stress and proliferation by TGF-β (24). Upregulated expression of TGF-β is associated with fibrotic disease. High serum levels of TGF-β were reported in chronic HCV infection (28). Although TGF-β1 inhibits the upregulation of P2X7 receptors in inflammation, the rise in P2X7 expression might be because of some intrinsic activity or due to the administration of interferon therapy (1).
Supporting our study, P2X7 activation promoted the growth of human cancer cells via downstream signaling of MAPK, AKT, and NF-κB-mediated pathways (26). Moreover, P2X7R have also found to be involved in RhoA activation in both macrophages and HEK-P2X7 cells (41). Previously, it was demonstrated that P2X7 accounts for apoptosis of immune cells, including macrophages, monocytes, and lymphocytes. In addition, this receptor is also involved in lymphoid cell proliferation, macrophage fusion, and release of proinflammatory cytokines. In HIV-1 infection purinergic signaling stimulates the activation of NLRP3 inflammasome and pyroptosis that are responsible for long-term inflammation (38). These results reveal downstream effects of nucleotide receptors in apoptosis, cell cycle, proliferation, and glucose metabolism.
In treatment responders the expression of P2X7 was up to 3.5-folds. These are the patients who received antiviral therapy for 6 months and showed viral reduction after 24 weeks. Previous studies have reported that IFN-δ and TNF-α induce the mRNA expression of P2X7 receptor (16). So, this noticeable increase in the P2X7 expression in treatment responders could be the outcome of HCV therapy, which makes use of interferon as an important antiviral agent (39). It is noteworthy that all the targeted markers except P2X7 and P2X4 receptors in this category showed decreased expression. It is because after achieving the treatment, the viral titer decreased and there are no viral structural proteins left to activate the mitochondrial damage and release excessive ATP molecules that may bind with these receptors and cause their opening. In this category P2X7 is not intrinsically high but is elevated due to interferon therapy. This might be a possible explanation of why targeted markers did not show increased expression even when P2X7 expression is enhanced.
Another interesting finding of our study was the unaltered expression P2X4 and P2X7 receptors along with host genes in treatment nonresponders. This accounts for the lack of immune system to combat virus due to its insufficient activation. Slight increase of eATP concentration in the extracellular environment may also result in receptor internalization or receptor downregulation, thus weakening the immune response elicited by the cells (21).
In conclusion, ion channel-mediated purinergic receptors P2X in viral infections not only participate in the innate and adaptive responses but also modulate the immune responses. Our study unleashes the possible outcome of P2X receptor activation on glucose metabolism, Akt, and MAPK pathway in PBMCs. These downstream signaling pathways may account for inflammation and pathogenesis associated with HCV infection. Moreover, as P2X7R is known to be upregulated during inflammation, antagonists of this receptor may serve as novel anti-inflammatory agents. Further research is required to explore therapeutic potentials of purinergic antagonist in chronic HCV patients.
Footnotes
Author Disclosure Statement
No competing financial interests exist.