
Abstract
U.S. military personnel deployed to high-risk areas receive the live vaccinia virus (VACV) smallpox vaccine ACAM2000. VACV shedding from the vaccination site can result in autoinoculation and contact transmission. We previously found that the application of povidone iodine ointment (PIO) to the scarification site reduced viral shedding without altering the antibody response, as measured by plaque reduction neutralization or enzyme-linked immunosorbent assays. In this study, we used protein microarray assays to measure the amount of immunoglobulin G antibody bound to (1) ACAM2000 itself and (2) individual VACV antigens that are present within ACAM2000. We assessed antibody binding in sera from primary smallpox vaccinees who applied PIO to the scarification site beginning on day 7 (PIO group) and from those who did not apply PIO (control group). In both cohorts, the postvaccination antibody response—in terms of antibody binding, both to ACAM2000 and to 11 individual VACV antigens—was significantly greater than the prevaccination response (all p < 0.0001). The postvaccination antibody binding levels of vaccinees in the PIO group did not differ from those of control vaccinees. These findings further support the topical application of PIO, starting on day 7, to reduce the viral shedding associated with smallpox vaccination.
Introduction
T
Previously, we reported (25) that VACV can continue to shed from the vaccination site for as long as 42 days after administration of the vaccine, continuing even after the scab separates in some individuals. Povidone iodine ointment (PIO), applied to the vaccination site starting on postvaccination day 7, reduced the levels and duration of viral shedding among primary vaccinees—compared with control vaccinees who did not apply PIO—without affecting the immune response. In that study, we assessed immunogenicity using methods such as a plaque reduction neutralization test (PRNT) and qualitative immunoglobulin G (IgG) and IgM enzyme-linked immunosorbent assays (ELISAs). In the present study, we subjected prevaccination and postvaccination sera from the same subjects to a protein microarray to compare primary vaccinees in the PIO and control cohorts for the amount of IgG antibody bound to ACAM2000 antigens. We assessed this in terms of (1) serum antibody binding to ACAM2000 itself (i.e., to unidentified antigens present within the vaccine), and (2) serum antibody binding to 11 individual VACV proteins known to be present within the ACAM2000 proteome, each of which we previously found (26) to be highly prevalent among smallpox-vaccinated individuals.
Materials and Methods
Subjects and vaccination
This study was conducted according to a protocol (FY08-19) approved by the Human Use Committee at the U.S. Army Medical Research Institute of Infectious Diseases. Informed consent was obtained from interested, eligible soldiers as described previously (25). Enrolled subjects were assigned to either the PIO group or the control group. Subjects in the PIO group were instructed to apply PIO to the scarification site with every dressing change (approximately every 1–3 days) beginning on postvaccination day 7; subjects in the control group were given the same instructions for dressing changes, but were not told to apply PIO. All participants in this study were primary vaccinees.
The lyophilized vaccine used in this study (lot VV04-003A; Sanofi Pasteur Biologics) was reconstituted according to the package insert (30) and was administered percutaneously, as described previously (25,26). Blood samples were collected from subjects before vaccination and on postvaccination day 28 (±2 days). Serum was separated and frozen until ready for use.
ACAM2000 microarray
To prepare the microarray, the lyophilized ACAM2000 vaccine, a replication-competent vaccine made from a single, plaque-purified VACV clone (New York City Board of Health strain, Dryvax clone 2), was reconstituted in 150 μL of 25% glycerol in sterile water. The vaccine vial contained ∼60 doses of 5–25 × 105 plaque-forming units in 0.0025 mL per dose. The following controls were included in the microarray: spotting buffer (1× phosphate-buffered saline [PBS], pH 7.4 with 25% glycerol) used for dilutions, dilutions of Vero cell lysates (50–400 μg/mL), and a 1:50 dilution of AlexaFluor 647-conjugated goat anti-human IgG secondary antibody. The reconstituted vaccine and controls diluted into the same buffer were printed onto nitrocellulose-coated SuperNova 16-pad slides (Grace Bio-Labs, Inc.) using a noncontact inkjet microarray printer (Arrayjet). Optimization steps were performed both for the amount of ACAM2000 antigen to be printed on the planar microarray surface and the dilution of serum sample to use for microarray assays. Specific serum antibody binding to printed array spots of the ACAM2000 vaccine was readily detected from neat to a 1:16 dilution. Because serum dilutions between 1:50 and 1:150 produced the best signals—statistically significant events above background and below saturation—we chose to use a dilution of 1:100 in this study. The vaccine was printed undiluted and in four 1:2 serial dilutions within the optimal range. Each pad on the microarray used a 12 × 4 layout, containing a total of 48 ligands, with the vaccine and controls printed in quadruplicate. Printed microarray slides were desiccated overnight under vacuum and stored at −20°C until use.
All incubations and microarray manipulations were performed at 22°C. Microarrays were blocked for 1.5 h with gentle shaking in 1× SuperG blocking buffer (Grace Bio-Labs, Inc.). The microarrays were rinsed with wash buffer (1× PBS, pH 7.4, 0.2% Tween-20, 1% bovine serum albumin [BSA]), and a ProPlate 16-well module (Grace Bio-Labs, Inc.) was applied to the microarray to perform several independent microarray experiments on the same slide. Microarrays contained within individual wells were probed (1 h) with 2 μL of serum diluted 1:100 in probe buffer (1× PBS, pH 7.4, 0.1% Tween-20, 1% BSA) and washed four times (5 min, wash buffer); antibody binding was detected by incubation (1 h) with a 1:2,000 dilution of goat anti-human IgG (H + L) AlexaFluor 647. The microarrays were washed as above and rinsed for 1 min with deionized water to remove residual salts.
Microarray slides were dried by centrifugation (300 relative centrifugal force, 2 min) and then scanned by confocal laser (GenePix 4000B; Molecular Devices), using a 635-nm wavelength. Raw pixel counts were generated by imaging the microarrays using a power setting of 33% and a photomultiplier tube gain of 450, which did not produce saturated signals (>65,500 relative fluorescence units [RFU]). Acquired images were analyzed with GenePix Pro 7 software, which corrected the median relative fluorescence for each microarray spot through local background subtraction.
VACV proteome microarray
Proteins encoded by VACV [Copenhagen; Genbank M35027.1; also see Ref. (9)] open-reading frames were produced as described previously (16,26,31). Briefly, sequence-verified entry clones were recombined into the pDEST20 (Invitrogen) glutathione S-transferase (GST) expression vector. All GST-tagged recombinant vaccinia proteins were expressed in Sf9 insect cells and purified to ≥80% purity using glutathione-based affinity purification. A total of 245 GST-tagged recombinant VACV proteins (∼90% of the VACV proteome) passed quality control criteria; these proteins, as well as several control proteins, were microarrayed onto thin-film nitrocellulose PATH slides (GenTel Biosciences). The protein microarrays were stored at −20°C until use.
All incubations and microarray manipulations were automated by using an HS 400 Pro Hybridization Station (Tecan Group Ltd), set at 22°C, using previously described (16,26,31) methods. Serum samples collected before vaccination and from 28 (±2) days after vaccination were normalized separately, and outliers among data replicates were removed from further analysis, as described previously (26).
As described previously (26), 21 VACV proteins elicited antibody responses that were more robust in postvaccination samples compared with prevaccination samples (p ≤ 0.0002). The 21 viral antigens that we identified included VACV surface proteins (intracellular mature virion [IMV], and extracellular enveloped virion [EEV]), as well as secreted and intracellular proteins. To examine whether the PIO and control cohorts differed in antibody response, we selected, for statistical analysis, 11 of the 21 proteins that were observed with ≥65% prevalence among postvaccination antibody responses. All 11 of the VACV proteins (A10L, A11R, A13L, A27L, A33R, C3L, C21/B27R, D8L, D13L, F13L, and H3L) are present within the ACAM2000 proteome.
Statistical analyses
An analysis of covariance model was used to compare the PIO and control groups in their postvaccination IgG antibody response to the ACAM2000 vaccine or to the individual VACV proteins, controlling for baseline (prevaccination) IgG antibody response. A paired t-test was used to compare the prevaccination and postvaccination IgG antibody response to the ACAM2000 vaccine and to the 11 VACV proteins within each cohort. We corrected for multiple testing using the Bonferroni method. After log2 transformation, IgG antibody binding-level data met assumptions of normality and homogeneity of variance. For correlation analyses between assays, Pearson's correlation coefficient (R) and coefficient of determination (R 2) were calculated from linear regression analysis. All statistical analyses were conducted using the SAS software system, version 9.4 (SAS Institute, Inc.).
Results
The 60 subjects who completed the study (30 individuals in each cohort) were predominantly male (68.3%), white (78.3%), and not Hispanic or Latino (96.7%) with an age range of 19–39 years (mean = 25 years). As described previously (25), the two cohorts did not differ significantly in baseline age, gender, race, or ethnicity or in the frequency of adverse events. All subjects had a classic “take” reaction on the day 7 visit.
Antibody responses to ACAM2000 vaccine
Figure 1A shows the prevaccination and postvaccination log2-transformed IgG antibody binding levels, measured in RFU, for each cohort. We found no significant difference between cohorts in postvaccination IgG antibody binding level: the geometric mean was 26,785 (95% confidence interval [CI]: 23,376–30,691) RFU and 24,233 (95% CI: 21,798–26,941) RFU in the control and PIO groups, respectively (p = 0.2947). In each cohort, the postvaccination level of IgG antibody bound was significantly greater than the prevaccination level (Fig. 1A and Table 1): the geometric mean increased from 13,153 (95% CI: 10,926–15,834) RFU prevaccination to 26,785 (95% CI: 23,376–30,691) RFU postvaccination in the control group (p < 0.0001) and from 9,565 (95% CI: 8,362–10,942) to 24,233 (95% CI: 21,798–26,941) RFU in the PIO group (p < 0.0001).
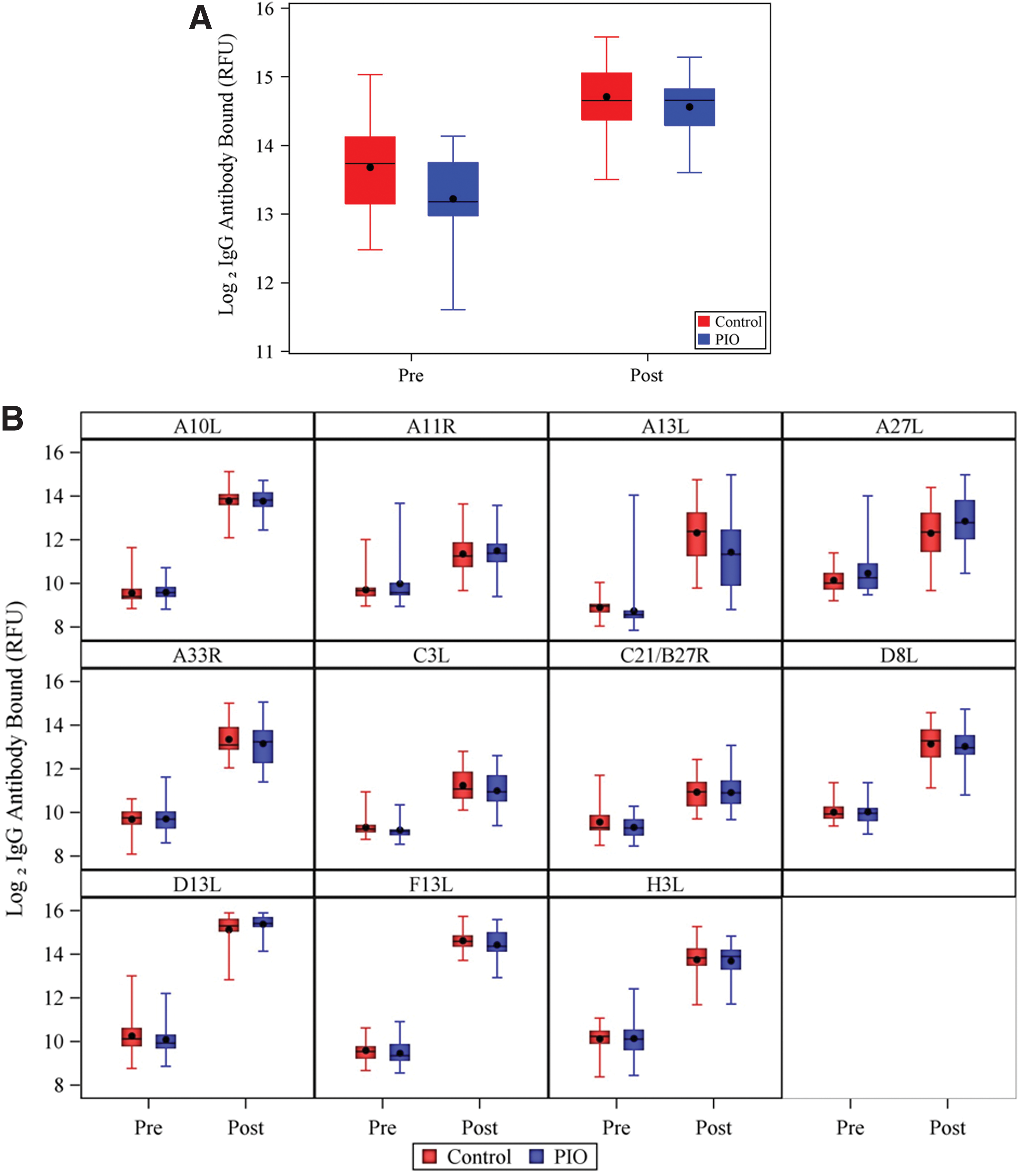
Distribution of prevaccination and postvaccination (day 28) log2 IgG antibody bound (in relative fluorescence units) to
Geometric mean values are in relative fluorescent units.
Percentage of subjects with at least a twofold increase between pre- and postvaccination in IgG antibody binding levels.
CI, confidence interval; EEV, extracellular enveloped virion; IEV, intracellular enveloped virion; IgG, immunoglobulin G; IMV, intracellular mature virion; IV, immature virion; PIO, povidone iodine ointment; VACV, vaccinia virus.
Antibody responses to VACV proteins
We found no significant differences between control and PIO cohorts in antibody binding to any of the 11 individual VACV proteins in postvaccination samples (Fig. 1B and Table 1) (p = 0.42–1.00). In each cohort, postvaccination IgG recognition of each of the 11 VACV antigens was significantly higher than that of prevaccination samples (all p < 0.0001).
Discussion
Previously, we found that PIO applied to the scarification site reduced viral shedding levels, the percentage of individuals who shed virus from the site, and the duration of viral shedding from the vaccination site among primary smallpox vaccinees (25). This reduction in shedding was achieved without affecting IgM, IgG, or PRNT seroconversion rates or other measures of immunogenicity (25). Hammarlund et al. (11) reported similar findings with PIO used after administration of Dryvax, the previously licensed VACV smallpox vaccine, with subjects who were mostly revaccinees.
The results of the present protein microarray study lend further support to our earlier conclusion that PIO application does not negatively affect antibody responses to smallpox vaccination. This was the case whether we examined levels of antibody bound to unidentified antigens within ACAM2000 (i.e., when the vaccine itself was printed on the microarray) or to 11 recombinant VACV antigens that are found within ACAM2000. Five of the VACV proteins, D8L, A13L, A33R, H3L, and A27L, were previously shown to elicit protective humoral immune responses, some of which were associated with virus neutralization (1,5,7,23,37).
The protein microarray provides a distinct approach for measuring antibody responses, and we would not expect the results of this assay to mirror exactly those of the PRNT. However, we did observe a moderate positive correlation between the previously reported (25) PRNT50 antibody titers from these subjects and IgG antibody binding to two IMV surface proteins, A13L and D8L (R = 0.4388, R 2 = 0.1925, p = 0.0005, and R = 0.3809, R 2 = 0.1451, p = 0.0029, respectively). We observed no significant correlation for the other individual VACV proteins (p = 0.1271–0.9693) or for the ACAM2000 vaccine (R 2 = 0.05, R = 0.22, p = 0.0836).
The protein microarray assay differs from other methods for evaluating immunogenicity. For example, the visual readout of the traditional PRNT is somewhat subjective (12), whereas the microarray results are based on the digital fluorescent units, which are a direct measurement of the relative level of IgG that bound to the VACV antigens within the spotted ACAM2000 vaccine. The vaccinia-specific IgG and IgM ELISAs performed in our earlier study (25) used VACV strain Western Reserve, a mouse-neurovirulent, highly passaged strain whose genome differs from that of ACAM2000 in a number of ways (24). In contrast, the microarray assay employed in this study used ACAM2000 itself. Furthermore, the microarray assay is higher throughput than the ELISA: it requires only microliter volumes of samples, but has the ability to assess antibody responses to multiple antigens on a single surface. By objectively measuring the amount of IgG from human serum that bound to antigens present within the smallpox vaccine, the microarray assay used in this study strengthens our previous conclusion: PIO can be applied to the scarification site without compromising the immune response to the smallpox vaccine.
The smallpox vaccine presented a moderate level of autofluorescence that impacted the overall sensitivity of the microarray assay. This may explain why we observed a greater increase in the level of antibody binding from baseline responses to individual VACV antigens than to the ACAM2000 vaccine (Table 1 and Fig. 1). Purified preparations of VACV may be an alternative test antigen.
In conclusion, applying PIO to the vaccination site and covering the site until 1–2 weeks after scab separation may reduce the rates of inadvertent autoinoculation and contact transmission of VACV without compromising the efficacy of vaccination with ACAM2000.
Footnotes
Acknowledgments
The research described herein was sponsored by the Military Vaccine Agency, USAMRIID plan number 1491541. This research was supported in part by an appointment of CP to the Postgraduate Research Participation Program at the U.S. Army Medical Research Institute of Infectious Diseases administered by Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and USAMRMC.
Author Disclosure Statement
No competing financial interests exist.
Opinions, interpretations, conclusions, and recommendations are those of the authors and are not necessarily endorsed by the U.S. Army. Research on human subjects was conducted in compliance with DoD, federal, and state statutes and regulations relating to the protection of human subjects and adheres to principles identified in the Belmont Report (1979). All data and human subjects research were gathered and conducted for this publication under an IRB-approved protocol, number FY08-19.