
Abstract
In this study, canine adenoviruses (CAdVs) from two acute fatal cases of infectious canine hepatitis (ICH) were analyzed using molecular detection and sequencing of the pVIII, E3, and fiber protein genes. Pathological findings in affected dogs were typical for CAdV-1 associated disease, characterized by severe centrilobular to panlobular necrohemorrhagic hepatitis and the development of disseminated intravascular coagulation in the terminal stages of disease. Comparison of partial genome sequences revealed that although these newly detected viruses mainly had CAdV-1 genome characteristics, their pVIII gene was more similar to that of CAdV-2. This likely suggests that a recombination has occurred between CAdV-1 and CAdV-2, which possibly explains the cause of vaccine failure or increased virulence of the virus in the observed ICH cases.
Introduction
C
Large genome capacity and unique replication strategy of adenoviruses have enabled them to tolerate wide ranges of recombination and gene manipulation. Experimental studies demonstrated that recombination in adenoviruses can produce strains with either intermediate or unique immunogenic and tropic properties (3,20). Previous findings with human adenoviruses have shown that intraspecies recombination is important for the emergence of new virus strains with improved fitness through changes in their antigenic properties or tropism (14,19,22,27,28).
The region of particular importance for mastadenoviruses is the early region 3 (E3), located between pVIII and fiber genes, suggested to play a role in virus pathogenicity by modulating the host's antiviral immune response and cell tropism (8,16).
In this study, two fatal cases of ICH associated with vaccine failure have been investigated by routine pathological examination and further molecular analysis. These were done to characterize the pathogen involved and to investigate the cause of vaccine failure observed.
Materials and Methods
Two cases of fatal acute hepatitis were submitted to the University of Adelaide's Veterinary Diagnostic Laboratory within a 1-month period during late summer 2013. Both the dogs were from the same geographical region of Adelaide Hills. Both cases occurred in young previously vaccinated dogs, a 6-month-old male intact Rottweiler (case 1), and a 11-week-old female intact poodle cross terrier (case 2). Both dogs were subjected to a complete postmortem and representative tissues were processed for histopathological examination using standard techniques. Fresh liver tissue samples were collected from each case for molecular analysis.
Total DNA and RNA was extracted from ∼50 mg liver tissue using a DNA extraction kit (the AxyPrep Multisource Genomic DNA Miniprep Kit, Axygen Biosciences) and the QIAamp Viral RNA Minikit (QIAGEN AG, Hombrechtikon, Switzerland), respectively.
RNA and DNA samples were subjected to in-house polymerase chain reaction (PCR) assays for canine distemper virus and canine parvovirus virus. For detection of the canine adenovirus genome, a pair of oligonucleotide primers CAdV-HA1 and CAdV-HA2 (Table 1) was used (13). This primer set produced a 508 bp product for CAdV-1 and 1,030 bp product for CAdV-2.
The primers CAdV1-HAF and R used for differentiation of CAdV-1 from CAdV-2 (13).
Internal primers used for sequencing of CAdV1-HAF and CAdV1-4R product.
To identify and select suitable primers for additional genomic sequencing, all available CAdV-1 full genome sequences were aligned using the ClustalW program (12). The primers with the closest match to the CAdV-1 reference strain (GenBank accession number AC_000003) were selected for synthesis. All primers were synthesized by GeneWorks (Australia).
To obtain a 2,506 nucleotide block from CAdV-1 samples, four sets of primer pairs that amplify the overlapping fragments encompassing the entire pVIII, E3 12.K, E3 ORF A, unidentified exon (U-exon), and partial fiber gene sequence were used (Table 1 and Fig. 1).

The binding sites for the selected primers that cover the sequences for pVIII, E3 12.K, E3 ORF A, U-exon, and partial sequence of fiber protein. The gene arrangement and the nucleotide positions retrieved from the CAdV1 reference strain (GenBank accession number AC_000003). CAdV, canine adenovirus. Color images available online at
Amplifications were carried out in a volume of 50 μL reactions, comprising of 5 μL 10 × High Fidelity PCR Buffer, 4 μL 50 mM MgSO4, 1 μL 10 mM dNTP Mix, 1 μL 10 μM of each primer, ∼500 ng DNA template, and 0.25 μL Platinum Taq DNA Polymerase (Invitrogen). The PCR cycling conditions were 2 min at 94°C, followed by 35 cycles of denaturation at 94°C for 15 s, annealing at 52°C for 30 s, and extension at 68°C for 1 min, followed by a final extension phase of 68°C for 5 min.
The amplified products were electrophoresed on a 1.5% agarose gel and DNA bands were purified using the QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany). Purified PCR products were submitted to the Australian Genome Research Facility Ltd (AGRF) for Sanger sequencing.
To confirm the correct sequence, all fragments were sequenced both in forward and reverse directions. To reconstitute the full-length sequence, overlapping sequences were edited and assembled using BioEdit package v.7.0.4.1 (11). Nucleotide sequences from CAdV-1 and CAdV-2 strains retrieved from GenBank were used as reference strains for nucleotide alignments using ClustalW program (12), and phylogenetic tree generation was reconstructed using the neighbor-joining method using the MEGA program (Version 6.06) (23). The reliability of the branching orders at each branch node was tested by 100 bootstrap replicates. To determine the probable recombination sites and relationship of two ICH viruses to CAdV-1 and CAdV-2 strains, a Boot-scanning program (SimPlot version 3.5.1) was used (18). Bootscan analysis was performed on the nucleotide sequence of selected CAdV-1 and CAdV-2 reference strains in comparison to the newly detected ICH viruses. To confirm the existence of the recombinant block, the resulting fragment for CAdV1-1F and CAdV1-1R primers was subcloned into the vector pGEMT-T Easy by T4 DNA ligated enzyme (Promega, Madison, WI). The recombinant vector was sequenced in both forward and reverse directions using vector-specific primers (M13).
Results
Case 1 presented to the referring veterinarian, collapsed and weak, with a short history of vomiting. This dog has received a complete puppy vaccination schedule in accordance with the manufacturer's recommendations (Virbac). Shortly after arrival, the dog developed cardiopulmonary arrest and died. Serum biochemistry performed on blood collected before death revealed markedly elevated serum ALT and a mild elevation of serum ALKP. At postmortem, the dog was diffusely pale, with hemabdomen, hemothorax, and widespread serosal, pleural, and endocardial petechial and ecchymotic hemorrhages. The liver was moderately enlarged with rounded margins, the parenchyma was friable and mottled dark red and orange with an enhanced lobular pattern, and petechial hemorrhages were present over the capsular surface. The thymus was expanded by edema and hemorrhage. Microscopically the liver showed acute centrilobular to panlobular necrosis and hemorrhage, and there was widespread hemorrhage and lymphocytolysis in the thymus and spleen. The pleural cavity contained 500–800 mL of unclotted blood. The lungs were incompletely collapsed and dark pink. Histological examination of the upper and lower respiratory tract revealed vascular congestion. Viral inclusions were not observed in the tissues examined.
Case 2 presented to the referring veterinarian with a history of vomiting, weakness, and mucohemorrhagic diarrhea. This puppy had received a single trivalent vaccine at 6 weeks of age (Boehringer Protech C3). Shortly after admission, the dog became obtund and nonresponsive, and developed cardiopulmonary arrest. Attempts at resuscitation were unsuccessful. Hematology and serum biochemistry performed before death identified marked elevation in serum ALT, mild azotemia, mild hyperbilirubinemia, as well as mild anemia and thrombocytopenia. At postmortem examination, the cadaver was pale and the liver was moderately enlarged with rounded lobar borders, an enhanced red–brown lobular pattern (Fig. 2), and edema of the gallbladder wall. The adrenal glands and thymus were edematous with multifocal petechial and ecchymotic hemorrhages, and there was diffuse mucosal congestion and intraluminal hemorrhage throughout the gastrointestinal tract. Microscopically, there was marked centrilobular to midzonal hepatocellular degeneration, necrosis, and hemorrhage (Fig. 3) and microscopic examination of the gallbladder wall confirmed mural edema and hemorrhage. Other findings included lobular hemorrhage and lymphocytolysis in the thymus, follicular lymphocytolysis and hemorrhage in the mesenteric lymph nodes and spleen, and adrenocortical medullary hemorrhage. Gross and histological examination of the upper and lower respiratory tract revealed no significant findings. Viral inclusions were not observed in the tissues examined.
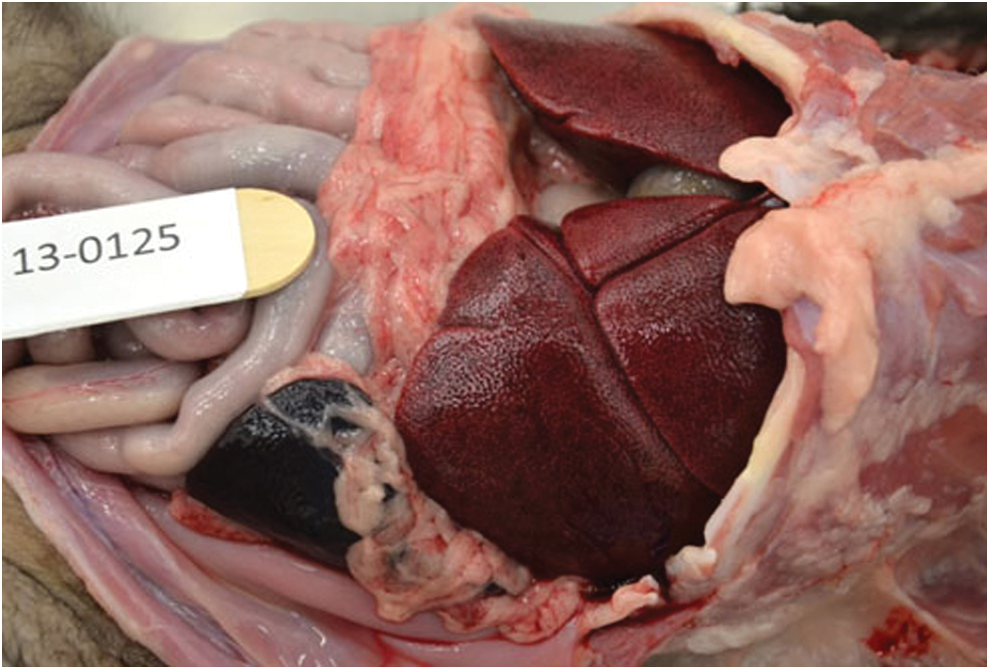
There is moderate to marked hepatomegaly with rounding of lobar borders. An enhanced red–brown lobular pattern was evident over the capsular and cut surfaces of the liver. 173 × 117 mm. Color images available online at
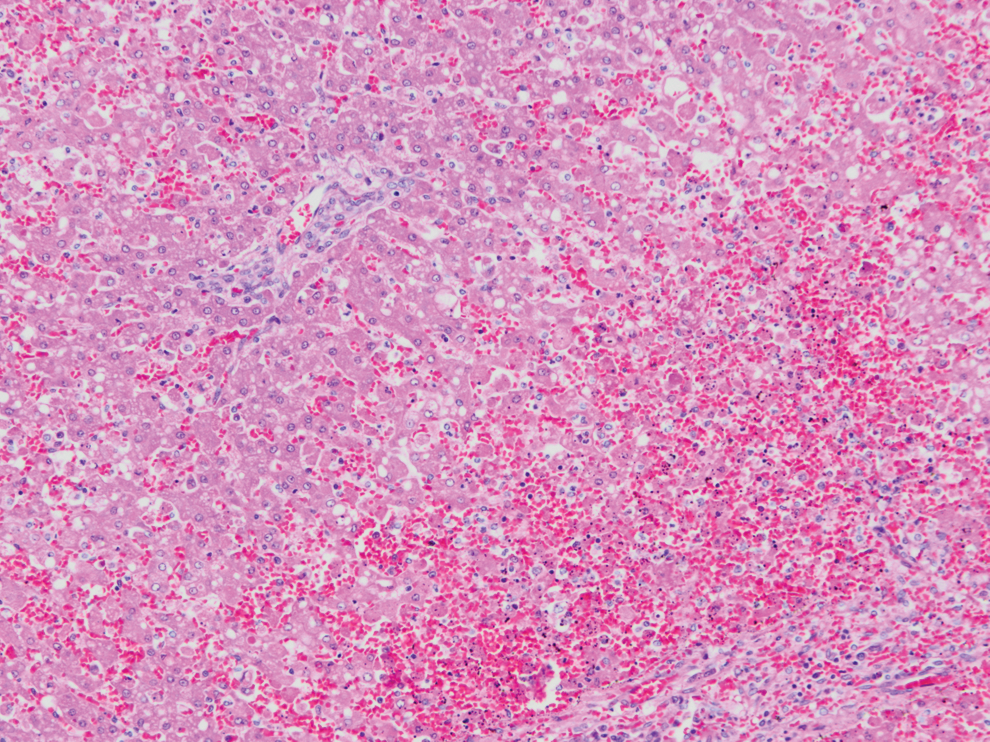
Liver from case 2, widespread severe centrilobular to midzonal hepatocellular degeneration, necrosis, and hemorrhage. HE (10 × objective). Similar histopathological lesions were seen in case 1. Color images available online at
In both cases, the PCR assays using the canine parvovirus and canine distemper virus were negative, whereas, CAdV1-HAV and CAdV1-HAR primers resulted in a single 508-bp fragment that was proven to originate from CAdV-1 genomes. An additional four genomic regions of pVIII (699 bp), E3 12.5K (858 bp), E3 ORF A and U exon (849 bp), and Fiber (1,009 bp, respectively) of the viral genome were amplified, purified, and submitted for sequencing. Partial genome sequences of the detected viruses were assembled based on the CAdV-1 reference strain genome (AC_000003). This provided a total of 2,470 bp sequences for each of the detected viruses. The nucleotide sequence data reported in this article have been submitted to GenBank with the accession numbers KT853096-KT853097.
Phylogenetic analysis of CAdV-1 and 2 sequences showed that the newly detected CAdV-1 showed 99.7% identity and shared some genome properties with both CAdV-1 and CAdV-2 and formed an independent sub cluster (Fig. 4). According to the nucleotide alignments of the two ICH viruses, a possible recombination event was indicated in the pVIII gene. For both viruses, a block of 477 bp nucleotide of pVIII gene showed a strong relationship with CAdV-2, whereas the rest of the genome was more similar to CAdV-1.
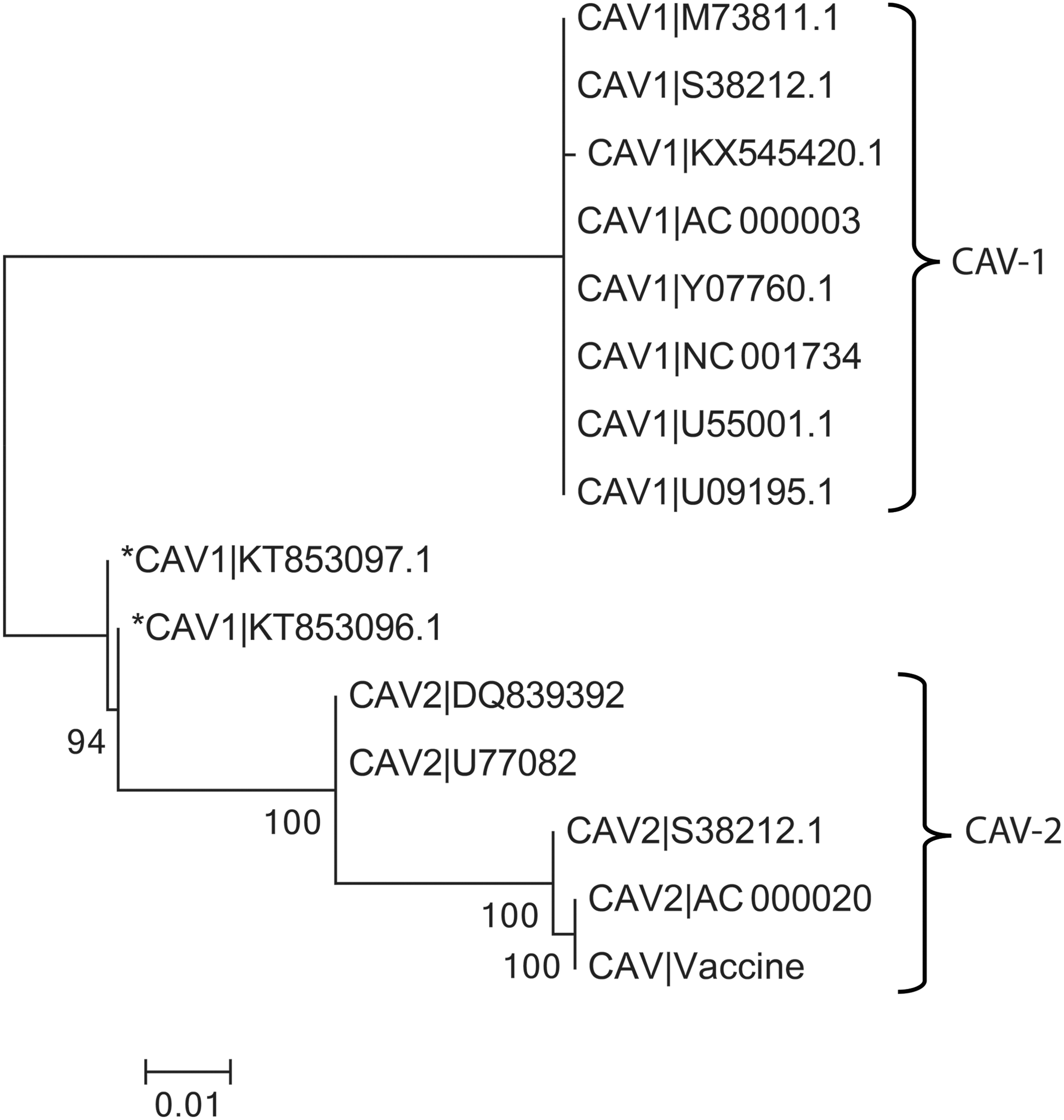
Neighbor-joining phylogenetic tree constructed by distance matrix analysis (100 replicates) of pVIII, E3 12.K, E3 ORF A, U-exon, and fiber protein fragments (2,470 bp length) comparing the novel canine adenovirus (*CAdV-1|KT853096.1 and *CAdV-1|KT853097.1) to other CAdV-1. Sequences were aligned using ClustalW program (12), and phylogenetic analysis was in Mega 6.06 version, under bootstrap resampling method with 100 replicates [branch length scale is shown at the left bottom and bootstrap value is shown at each branch].The detected viruses are indicated by asterisk (*).
Sequence identity matrix analysis (BioEdit package v.7.0.4.1.) on the 2,470 bp sequences showed that the newly detected viruses possessed 97.4% identity to CAdV-1 and 68.8% to CAdV-2 reference strains (data not shown). At the same time, the sequence identity matrix for the pVIII gene of the CAdV-1 and CAdV-2 viruses revealed 98.8% to 99.9% identity to CAdV-2 reference strain and the sequence identity matrix for the whole 2,470 bp sequence for the new viruses was 86% to the CAdV-1 reference strains. This finding revealed that the entire sequence of the new viruses showed similarities to the CAdV-1 genome, except a part of pVIII that suggested possible recombination with CAdV-2.
In the Bootscan analysis of query sequences, a sudden alteration in similarity plots indicates a recombination crossover in pVIII gene in CAdV-2. It is clearly demonstrable in sequence alignments that a block of 477 bp of the CAdV-1 and CAdV-2 viruses showed 98% homology to pVIII gene of CAdV-2 viruses (Fig. 5).
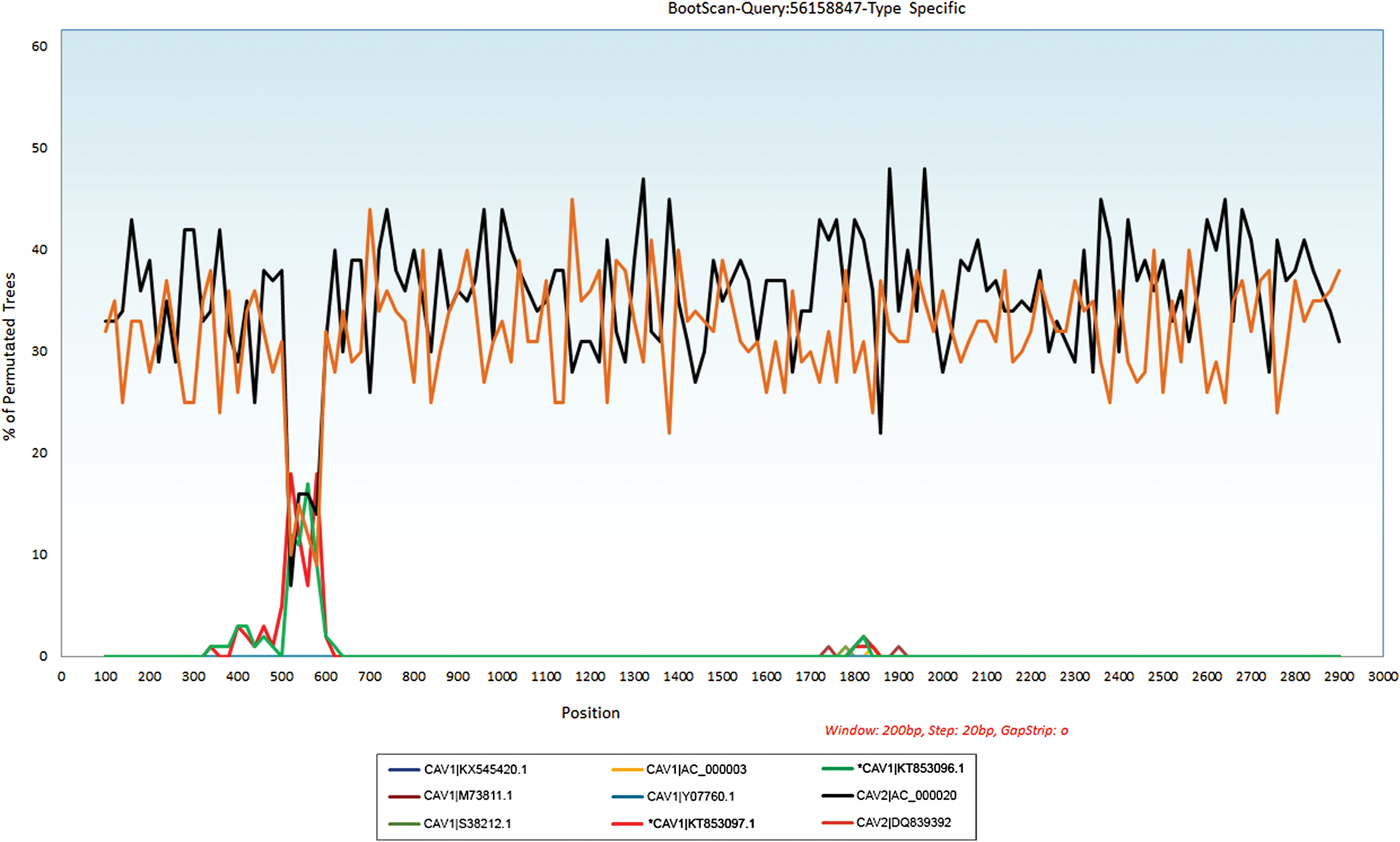
Bootscan for recombination analysis using SimPlot (3.5.1). Bootscan plots demonstrate a possible recombination between new ICH viruses and the CAdV2 strains. The upstream and downstream parts of the recombination site demonstrate the sequence similarity between the query sequences and the CAdV-1 strains. The Y axis gives the percentage of permutated trees using a sliding window of 200 bp wide centered on the position plotted, with a step size between plots of 20 bp. CAdV-1|KT853096.1 and CAV1|KT853097.1, CAdV1 and CAdV2-type strains were used as the outgroup to determine the breakpoint. ICH, infectious canine hepatitis. Color images available online at
Discussion
Intramolecular recombination plays an important role in the evolution of viruses. Possible recombination and its role in the alteration of viral properties have been demonstrated in some human and animal adenoviruses (4,27). Genetic recombination between human adenovirus has been reported in mixed infection in cell culture studies. Incidences of coinfection in vaccinated patients have been reported to contribute to emergence of new recombinant variants, resulting in breakthrough infections (25). In this study we demonstrated a unique recombination in newly detected ICH viruses from acute lethal forms of the infection. These viruses were detected from liver samples of two fatal cases of ICH associated with CAdV vaccine failure. The presence of clinical disease in these dogs with documented vaccination history could be due to improper vaccination, or as a result of recombination of CAdV-1|KT853096.1 and CAdV-1|KT853097.1 viruses, or emergence of a new virus (1,6). Vaccine failure due to improper vaccination cannot be detected and pre-exposure serum samples are necessary to find the antibody titers against CAdVs (24), and for these two cases the pre-exposure serum were not available to test.
In this report, we described the possible connection between the novel recombination in pVIII gene and two fatal ICH cases. Instead of significant genome stability in adenoviruses, the intertypic recombination and its roles in virulence of adenoviruses were mentioned by different researchers (24). In genome analysis, the partial pVIII gene showed 98% homology to the CAdV2 strains. The possible role of recombinant pVIII gene in the pathogenicity of CAdV-1 has not been described and it needs to be evaluated at both molecular and host animal levels. It has been described that the mutations of the pVIII can affect the virion's sensitivity to heat and thus the infectivity of the virus (17). It has been shown in different viruses that the alteration of the surface antigens plays a big role in vaccine failure (9).
Pathological findings in these two cases were typical of and consistent with CAdV-1 infection, characterized by severe necrohemorrhagic hepatitis and subsequent disseminated intravascular coagulation as the cause of death, although viral inclusions were not identified within affected tissues. Gallbladder wall edema, a classic and common lesion associated with ICH, was only observed in case 2 (1). As it has been mentioned in the Materials and Methods section, the generic materials from both cases were subjected to diagnostic PCR assays for canine parvovirus and canine distemper viruses. The PCRs for these viruses showed negative results. Both the molecular and histopathology results support detection of CAdV-1 virus in both cases.
There are number of vaccines in the market that protect against CAdV-1, and those are generally derived from CAdV-2, as they produce effective immunity against CAdV-1. Existing use of CAdV-2 strains as the source of vaccines and exposure of animals to CAdV-1 viruses provides a plausible condition for virus coinfection and possible viral recombination in vivo (3,4). Adenoviruses are generally host species specific, specifically within human and usually infect representatives of a single, or sometimes several closely related animal species. Only adenovirus interspecies transmission has been mentioned in a number of reports (2,15,29). In contrast, CAdVs have been detected in a wide range of other carnivores like wolves, raccoons, sea lions, and bears (26). In a recent study of bat adenovirus type 2 (BtAdV-2), a close phylogenetic relationship to CAdVs was observed (15).
As has been mentioned in case histories, both dogs were from the same geographical region with no evidence of possible contact. This new virus may have a broader geographical range, however, given the limited number of the submitted, samples from suspected CAdV-1 disease for molecular diagnosis, additional work is required to determine this formally to demonstrate the distribution and the significance of the new CAdV-1 in acute infections in dogs.
Detection of this putative CAdV-1 recombination event in combination with the pathological findings in these two cases suggest increased virulence of fatal disease, and suggests that the modified virus may be a highly virulent strain or any other causes due to vaccine failure. Complete genome sequencing of this novel recombinant virus should be pursued in an attempt to determine the recombination events that may have led to increased virulence of the virus, and apparent vaccine failure in these two cases of fatal ICH.
Footnotes
Acknowledgments
This work was supported by funds provided by the University of Adelaide. The granting agencies had no role in study design, data collection and analysis, or article preparation. The authors would like to acknowledge Dr. Wayne Boardman and Dr. Melanie Buote from The University of Adelaide, Dr. Mark Kelman from The Disease Watchdog, and veterinarians from the Woodville Veterinary Clinic for their assistance with case histories. The authors would also like to thank Ms Faustina Niap from the University of Adelaide for providing technical assistance.
Author Disclosure Statement
No competing financial interests exist.